Книга: Эволюция человека. Книга 2. Обезьяны нейроны и душа
Назад: А вот если бы мы все были клонами…
Дальше: Альтруизм, стремление к равенству и нелюбовь к чужакам
Межгрупповая вражда способствует внутригрупповому сотрудничеству
Если у какого-то вида животных кооперация уже развилась настолько, что вид перешел к общественному образу жизни, если сформировались компактные социальные группы с относительно постоянным составом, то появляется новый фактор, который может подстегнуть развитие внутригрупповой кооперации. Этот фактор — межгрупповая конкуренция. Теперь уже не только индивиды конкурируют между собой за ценные ресурсы, но и группы. Шансы одиночек (и маленьких, слабых групп) резко снижаются. "Корыстные интересы" индивида оказываются неразрывно связаны с интересами коллектива. При острой межгрупповой конкуренции индивид может успешно размножиться, только будучи членом успешной группы.
К чему это приводит, показывает модель, которую разработали американские этологи Керн Рив и Берт Холлдоблер (Reeve, Holldobler, 2007). Непосредственной целью исследователей было найти объяснение некоторым закономерностям, наблюдаемым в социальном устройстве общественных насекомых. Авторы упоминают четыре таких закономерности:
1. самая развитая кооперация (и наивысший уровень альтруизма, когда большинство особей отказывается от размножения ради заботы о чужом потомстве) характерна для коллективов, связанных близким родством;
2. у видов с большими колониями кооперация обычно развита лучше; в маленьких колониях чаще наблюдается эгоистическая "грызня" между соплеменниками за право оставить потомство;
3. у видов с мелкими и средними колониями общая производительность (отношение числа произведенных потомков к численности колонии) снижается с ростом колонии;
4. острая межгрупповая конкуренция, усиливающаяся при неравномерном пространственном распределении дефицитного ресурса, обычно коррелирует с высокоразвитой внутригрупповой кооперацией.
В модели Рива и Холлдоблера каждый индивид эгоистически расходует часть "общественного пирога" на то, чтобы увеличить свою долю этого пирога. Эта потраченная на внутригрупповые склоки часть называется "эгоистическим вкладом" или "эгоистическим усилием" данного индивида. Доля, доставшаяся в итоге каждому индивиду, зависит от соотношения его собственного "эгоистического усилия" и суммы "эгоистических усилий" остальных членов группы — подобно тому как в перетягивании каната итог определяется соотношением сил, приложенных к двум концам каната. Нечто подобное наблюдается у общественных насекомых, когда они осуществляют "взаимный надзор" — мешают друг другу откладывать яйца, стараясь при этом отложить свои (см. выше).
На тех же принципах строятся в модели и взаимоотношения между группами. Таким образом, получается вложенное, двухуровневое перетягивание каната. Чем больше энергии тратят индивидуумы на внутригрупповую борьбу, тем меньше ее остается для межгруппового перетягивания и тем меньше получается "общий пирог" группы.
Исследуя эту модель, авторы показали, что она неплохо воспроизводит (а значит и объясняет) все четыре перечисленные закономерности.
Мы не будем глубоко погружаться в математические дебри, но пару формул я все-таки приведу — хотя бы для демонстрации образа мысли авторов. Читатели, не любящие формул, могут их смело пропустить (тем более что все объясняется в тексте).
Если в группе есть редкие "мутанты", отличающиеся от сородичей величиной своего эгоистического усилия, то полученная мутантом персональная доля "общественного пирога" (ресурса, добытого группой) вычисляется по формуле:
S = х/(х + r (n — 1) х + (1 — r)(n-1)x*),
где х — эгоистическое усилие мутанта (х = ft, где t — изначальный "запас энергии" индивида, f — доля этого запаса, которую он потратит на эгоистические цели); r — это величина из формулы Гамильтона: степень внутригруппового родства, варьирующая от 0 (нет общих генов) до 1 (все гены общие); n — число особей в группе; х* = f*t — эгоистическое усилие немутантных членов группы. Здесь учитывается, что если в группе есть мутант и есть заданная внутригрупповая степень родства (г), то соответствующая фракция всей группы (r) будет иметь "мутантный ген" и проявлять "мутантное поведение", то есть демонстрировать эгоистическое усилие, равное х, а не х*.
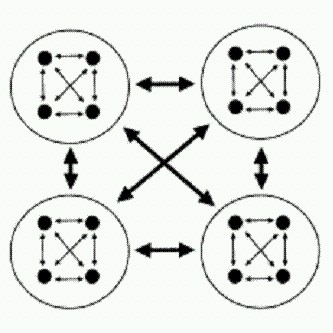
Вложенное перетягивание каната. Члены группы соревнуются друг с другом за свою долю "общественного пирога". Размер "пирога" в свою очередь зависит от успешности группы в соревновании с другими группами. Чем больше сил тратят особи на внутригрупповую борьбу, тем меньше их остается на общественно-полезную деятельность. По рисунку из Reeve, Holldobler, 2007.
Конкурентоспособность этой группы (G) в соревновании с другими группами вычисляется как сумма всех индивидуальных "неэгоистических" усилий, направленных на рост благосостояния колонии (неэгоистическое усилие равно (1 — f) t для мутантов и (1 — f*) t для не-мутантов).
Доля ресурса, отвоеванная группой в борьбе с другими группами (тот самый "общий пирог", который делят между собой члены группы), вычисляется по формуле:
S = G/(G + r' (N — 1) G + (1 — r') (N — 1) G*),
где r' — степень межгруппового родства, N — число групп, G* — конкурентоспособность тех групп, в которых нет мутантов.
Затем, играя с этими уравнениями по законам теории игр, авторы нашли эволюционно стабильное значение f*, то есть такую величину индивидуального эгоистического усилия, при которой никакой новый мутант не сможет распространиться в популяции при данных значениях четырех переменных (N, n, r и r').
Решение получилось такое:
f* = N (n — 1) (1 — r)/(Nn — 1 — г (n — 1) — (N — 1) r' (1 + r (n-1))).
Величина (1 — f*) отражает интегрированность, или "сверхорганизменность" колонии. Когда 1 — f* = 0, никакой кооперации нет, и группы как таковой тоже, это полный индивидуализм. Когда величина 1 — f* близка к единице, колония становится настоящим сверхорганизмом, аналогом многоклеточного животного.
Модель предсказывает, что внутригрупповая кооперация (эволюционно стабильное значение 1 — f*) должна расти с ростом внутригруппового родства (r). Это наблюдается в природе (см. выше закономерность № 1) и полностью соответствует теории родственного отбора. Данный результат говорит о том, что степень родства между членами группы — отнюдь не второстепенный фактор, как считают некоторые, а мощный регулятор развития кооперации. Максимальная кооперация, согласно модели, должна наблюдаться в колониях, где большинство индивидуумов — родные братья и сестры, то есть дети одной пары насекомых-основателей (царя и царицы). "Многоцарствие" должно снижать кооперацию и способствовать внутригрупповым конфликтам, однако этот эффект, согласно модели, может быть сглажен острой межгрупповой конкуренцией, то есть большим числом конкурирующих колоний (N) (закономерность № 4).
Кроме того, модель предсказывает нечто весьма удивительное и из теории родственного отбора не вытекающее. Она предсказывает, что "гены кооперации" могут быть поддержаны отбором даже при полном отсутствии родства между членами группы. Из последней формулы следует, что при отсутствии родства (r = r' = 0) внутригрупповая кооперация (1 — f*) будет больше нуля при наличии хоть какой-то межгрупповой конкуренции (N > 1).
Модель предсказывает, что при неизменном числе конкурирующих колоний (N) рост размера колонии (п) должен снижать кооперацию. Казалось бы, это противоречит закономерности № 2. По мнению авторов, тут все дело в том, что и размер колоний, и их количество на единицу площади зависят от неравномерности распределения дефицитных ресурсов и от богатства участков, где ресурс есть. Чем богаче участок, тем обычно больше на нем колоний общественных насекомых и тем крупнее каждая колония. Таким образом, один и тот же фактор (богатство участка) положительно коррелирует с обеими переменными (n и N). Когда богатство участка растет, одновременно увеличиваются и n (что ведет к снижению кооперации), и N (что ведет к ее росту). Но в модели степень кооперации чувствительнее к изменениям N, чем n. Поэтому в итоге кооперация все-таки растет.
Само по себе снижение кооперации при росте размера колонии (n) соответствует закономерности № 3 и говорит о том факте, верно отраженном моделью, что в больших коллективах вольготнее чувствуют себя обманщики, паразитирующие на чужом альтруизме (в модели это "мутанты" с повышенным значением f).
Авторы полагают, что их модель может быть приложима не только к насекомым, но и к другим социальным животным, включая человека. Особенно интригующим выглядит предсказание модели, согласно которому кооперация должна снижаться с ростом межгруппового родства (r'). Модель предсказывает, что кооперация должна быть развита сильнее у тех видов, у которых дочерние колонии закладываются далеко от материнских. Кроме того, межвидовая конкуренция должна лучше способствовать сплочению колонии, чем внутривидовая (так как в первом случае межгрупповое родство нулевое: r' = 0).
Предельный случай альтруизма и "сверхорганизменности" у насекомых — это полная утрата рабочими особями способности к размножению. Согласно модели, это должно происходить либо при чрезвычайно острой межгрупповой конкуренции, либо при очень высокой степени внутригруппового родства.
Главный вывод, который можно сделать из этой работы, состоит в том, что межгрупповая конкуренция — мощнейший стимул для развития внутригрупповой кооперации и альтруизма.
Аналогии с человеческим обществом вполне очевидны. Ничто так не сплачивает коллектив, как совместное противостояние другим коллективам. Множество внешних врагов (N) — обязательное условие существования тоталитарных империй и надежное средство "сплочения" населения в альтруистический муравейник.
Назад: А вот если бы мы все были клонами…
Дальше: Альтруизм, стремление к равенству и нелюбовь к чужакам

