Бактериальный суп
Когда одну бактерию помещают в богатый питательными веществами бульон и позволяют делиться сначала на две бактерии, потом на четыре, потом на восемь и так далее, происходит интересная вещь. Как мы видели ранее, всякий раз, когда ДНК воспроизводится — во время размножения — возникают случайные ошибки, называемые мутациями. Это те самые изменения в рецепте супа, которые происходят естественным образом при передачи его следующему поколению. Такая же картина наблюдается при делении бактерий. Таким образом, в нашем быстро заселяемом бактериями бульоне мы начинаем обнаруживать уникальные генетические линии, образующиеся в результате небольших изменений в их геномах. Если мы исследуем последовательности ДНК бактериальной популяции через несколько поколений, мы едва ли увидим различия между ними. Но если подождем несколько сотен поколений (для бактерий это всего лишь пара дней), то увидим огромное количество вариаций. Как и в случае эволюции белка в понимании Цукеркандля и Полинга, чем дольше растет популяция, тем больше мы видим изменений. Проще говоря, генетических различий больше между двумя бактериями, случайно отобранными из более старой популяции, чем между бактериями из более молодой популяции.
Эксперимент, который мы только что провели с нашим бактериальным супом, показал, что происходит в любой экспоненциально растущей популяции, где потомство удваивается в каждом поколении. Совершенно очевидно, что популяция быстро увеличивается в размерах — если бы мы действительно позволили бактериям беспрепятственно делиться в течение нескольких дней, они бы заполонили всю нашу планету. Однако гораздо важнее для нашей истории причина такого бурного роста популяции. А заключается она в том, что в популяции каждый индивидуум оставляет потомство. Никто не проигрывает в эволюционной лотерее — у каждой бактерии есть дети, у всех ее детей есть дети, и так далее. Это оказывает интересный эффект на генетическую структуру популяции.
Как мы теперь знаем, ответ на вопрос, сколько в среднем генетических различий между бактериями, составляющими растущую популяцию, зависит от того, как долго популяция росла. В действительности существует распределение различий между отдельными бактериями, отображаемое колоколообразной кривой Гаусса, которой нас мучили в школе на уроках математики. Среднее значение этого распределения (среднее количество различий между индивидуумами в образце) зависит от времени, в течение которого растет популяция. Если мы представим себе кривую как волну, движущуюся по мере накопления различий слева направо, то чем правее (другими словами, чем дальше от нуля), тем больше в популяции накопилось мутаций. И как при сравнении последовательностей гемоглобина лошади и гориллы, скорость, с которой волна движется слева направо, предсказуема, потому что скорость, с которой случаются мутации — постоянна, наши молекулярные часы отсчитывают время в А (а также в Ц, Г и Т). Благодаря этому мы можем вычислить время экспоненциального роста популяции, измеряя среднее значение распределения — середину волны. «Хорошо, — скажете вы, — это может быть интересно в качестве лабораторного упражнения университетского курса генетики, но совсем не относится к предмету нашего обсуждения… если, конечно, мы не увидим такую же модель и для других организмов».
Генри Харпендинг, антрополог из Пенсильванского университета, провел со своими коллегами анализ распределения генетических различий между последовательностями человеческой митохондриальной ДНК и обнаружил поразительную закономерность. Во-первых, распределение различий, называемое распределением несоответствий, совершенно ясно указывало на то, что человеческая популяция действительно росла так же быстро, как и бактерии. Плавная колоколообразная кривая красноречиво указывала на то, что человеческий вид разрастался с большой скоростью. В популяциях, имеющих постоянный (или уменьшающийся) размер, кривая распределения начинает деградировать, становясь со временем зубчатой из-за неравномерной потери генетических линий вследствие генетического дрейфа или, возможно, отбора. Таким образом, существует ясный генетический сигнал о быстром увеличении человеческой популяции. Но самым интересным стало рассчитанное Харпендингом время предполагаемого начала ее увеличения — примерно 50 000 лет назад, что соответствует нашей оценке времени, когда люди современного типа начали мигрировать из Африки, и почти точно — наступлению верхнего палеолита.
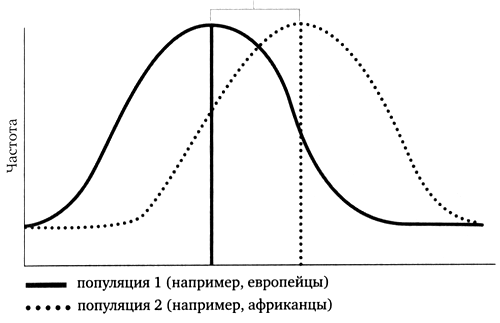
Рисунок 5. Распределение несоответствий митохондриальных ДНК (мтДНК) двух растущих популяций. Чем дольше росла популяция, тем выше среднее значение различий в последовательностях ее мтДНК
Харпендинг и его коллеги изучили данные по мтДНК двадцати пяти популяций из разных уголков Земли, и все они, за исключением двух, свидетельствовали об экспоненциальном росте этих популяций на протяжении последних 50 000 лет. Как показали другие данные, две популяции с зубчатым распределением недавно резко уменьшились в размере, и, таким образом, подобный анализ действительно способен дифференцировать эти два сценария. Кроме того, похоже, что популяции увеличивались фактически независимо друг от друга. Первыми примерно 60 000 лет назад начали африканцы, вслед за ними 50 000 лет назад — азиаты и, наконец, европейцы — 30 000 лет назад. Это был ошеломляющий результат. Данные по мтДНК полностью согласовывались с археологическими свидетельствами развития технологии верхнего палеолита: сначала в Африке, затем в Азии и, наконец, в Европе — совпали даже даты. Казалось, что «большой скачок» оставил свой генетический след в нашей ДНК, зафиксировав развитие доминирующих технологий по всему миру. В нем же содержался и намек на маршрут, однако деталей путешествия пришлось ждать до тех пор, пока дорогу не показали сыновья Адама.
Назад: Гимнастика для ума
Дальше: Глобальное похолодание

