Книга: Слепой часовщик. Как эволюция доказывает отсутствие замысла во Вселенной
Назад: Глава 9 Пререкания о прерывистом
Дальше: Глава 11 Обреченные противники
Глава 10
Единственное истинное древо жизни

Главный предмет этой книги — эволюция как решение проблемы сложного “замысла”, как правдивое объяснение тех явлений, которые, по мнению Пейли, долж ны доказывать существование божественного часовщика. Вот почему в своем повествовании я постоянно возвращаюсь то к глазу, то к эхолокации. Но есть еще целый ряд вещей, объясняемых теорией эволюции. Есть, например, явления, связанные с биологическим разнообразием: распределение различных типов животных и растений по планете и различных признаков между ними. И хотя в первую очередь меня интересуют глаза и прочие детали сложных механизмов, не следует пренебрегать и другими сторонами эволюционной теории, которые помогают нам понять природу. Так что эта глава будет посвящена систематике, или таксономии.
Таксономия — это наука о классификации. Некоторые люди незаслуженно считают ее скучной, отдающей музейной пылью и формалином, как будто бы это не таксономия, а таксидермия. На самом же деле ее можно назвать какой угодно, но только не скучной. По причинам, не вполне мне ясным, это одна из тех областей биологической науки, в которых случаются самые бурные перепалки. Ею интересуются философы и историки. Она играет значительную роль в любой околоэволюционной полемике. И именно из рядов систематиков вышли некоторые из тех современных биологов, которые откровенно называют себя антидарвинистами.
Хотя в основном систематики занимаются изучением животных и растений, классифицировать можно все что угодно: минералы, военные корабли, книги в библиотеке, звезды, языки. Упорядоченная классификация обычно считается вопросом удобства, практической необходимости, и отчасти это действительно так. Большая библиотека будет практически бесполезна, если не расположить в ней книги таким неслучайным образом, чтобы при желании всегда можно было отыскать нужные. Науку — или, если угодно, искусство — библиотечного дела можно рассматривать как пример прикладной систематики. Руководствуясь сходными соображениями, биологи тоже находят для себя удобным рассортировывать живые организмы по “полочкам” с общепринятыми наименованиями. Но сказать, что систематика животных и растений нужна только для этого, значит упустить почти все самое важное. Для биолога-эволюциониста классификация живых существ представляет собой нечто особенное, нечто отличное от всех прочих разновидностей систематики. Из наших представлений об эволюции следует то, что для всех существующих организмов можно составить только одно правильное генеалогическое древо и на нем основывать свою классификацию. Помимо своей уникальности такая система будет обладать еще одним замечательным свойством, которое я назову “идеальной вложенностью”. Что это за свойство и почему оно столь важно — главная тема настоящей главы.
Возьмем в качестве примера небиологической таксономии библиотеку. Уникального, единственно верного способа систематизировать книги в библиотеке или в книжном магазине не существует. Какой-нибудь библиотекарь, возможно, разложит все собрание книг по следующим основным категориям: наука, история, литература, другие искусства, книги на иностранных языках и т. д. Каждый из этих главных библиотечных отделов тоже, в свою очередь, будет разделен. Например, крыло библиотеки, отведенное под научную литературу, займут такие отсеки, как биология, геология, физика, химия… А в биологическом отделении научного крыла книги будут, вероятно, рассортированы по отдельным стеллажам, посвященным физиологии, анатомии, биохимии, энтомологии и т. п. Наконец, на каждом из этих стеллажей книги можно расставить в алфавитном порядке. Другие главные отделы библиотеки — исторический, литературный, иноязычный и прочие — будут приведены в систему сходным образом. Такая библиотека будет иметь иерархическую структуру, которая позволит читателю прицельно отыскивать интересующую его книгу. Иерархическая классификация в данном случае удобна: благодаря ей пользователь библиотеки сможет быстро ориентироваться в книжном собрании. Слова в словарях расположены по алфавиту с той же целью — для удобства ориентирования.
Но не существует никакой единственной иерархии, в соответствии с которой мы были бы обязаны систематизировать библиотечные книги. Другой библиотекарь, возможно, предпочел бы другую, хотя и тоже иерархическую классификацию. К примеру, не стал бы объединять вместе все книги на иностранных языках, а разместил бы их по тематическим отделам: биологические книги на немецком — в биологическую секцию, исторические книги на немецком — в историческую, и так далее. Третий библиотекарь подошел бы к вопросу радикально, расставив все книги независимо от того, о чем они, в хронологическом порядке по году издания, а для тематического поиска использовал бы карточный каталог (или его компьютерный эквивалент).
Эти три способа организации библиотеки довольно сильно отличаются друг на друга, но любой из них, по всей вероятности, работал бы нормально и пришелся бы по нраву многим пользователям, даже если бы, к слову, и не удовлетворил того раздражительного пожилого завсегдатая одного из лондонских клубов, которого мне довелось как-то слышать по радио. Он на чем свет стоит ругал правление своего клуба за то, что оно наняло библиотекаря: клубная библиотека обходилась без какой-либо организации сотни лет, так зачем же понадобилось что-то менять теперь? Журналист осторожно поинтересовался, как же, по его мнению, следует систематизировать книги. “По размеру: большие слева, маленькие справа!” — ни на секунду не задумываясь, проревел недовольный. Книжные магазины, рассчитанные на широкую публику, сортируют свои книги в соответствии с массовым потребительским спросом. Вместо науки, истории, литературы, географии и т. п. их основные отделы посвящены садоводству, кулинарии, телепередачам, оккультизму, а однажды мне попался стеллаж с огромной, бросающейся в глаза надписью “РЕЛИГИЯ И НЛО”.
Итак, правильного ответа на вопрос, как классифицировать книги, не существует. У библиотекарей могут быть серьезные разногласия на этот счет, но критерии, по которым будет оцениваться та или иная система классификации, не будут связаны с ее “истинностью” и “правильностью”. Вместо этого дискуссия будет строиться вокруг таких критериев, как “удобство для посетителей”, “скорость нахождения книг” и т. п. В этом смысле систематику книг в библиотеке можно назвать произвольной. Это не значит, что разработка удобной системы классификации в данном случае неважное дело, вовсе нет. Но это значит, что даже если бы мы жили в идеальном мире и владели бы всей полнотой информации, то какой-то одной системы, которую можно было бы признать за единственно верную, тут не существовало бы. А вот систематика живых организмов, как мы увидим, отличается той строгостью, которой лишена систематика книг. По крайней мере если смотреть с эволюционных позиций.
Разумеется, можно придумать множество систем для классификации живых существ, но я собираюсь показать, что все они, кроме одной, будут так же произвольны, как и любая библиотечная система расстановки книг. Если классификация животных создается исключительно ради удобства, то музейный хранитель может распределять экспонаты по размеру и способу их хранения: большие в виде чучел, маленькие засушенные на булавках, законсервированные в склянках, микроскопические на стеклах и так далее. В зоопарках тоже принято группировать животных по принципу удобства. В Лондонском зоопарке носороги живут в “Слоновьем доме” по той лишь причине, что для них нужны такие же массивные решетки, как и для слонов. Тот, кто занимается прикладными аспектами биологии, может делить животных на вредных (которые в свою очередь будут подразделяться на возбудителей заболеваний человека, сельскохозяйственных вредителей, кусачих и жалящих), полезных и нейтральных. Диетолог мог бы создать классификацию животных, основываясь на пищевой ценности их мяса для человека, причем и в этом случае разделение на категории и подкатегории оказалось бы сложным и замысловатым. Моя бабушка вышила однажды тряпичную книжку для детей, где животные распределялись по группам в зависимости от формы лап. Антропологи сообщают об изощреннейших системах классификации животных, используемых различными племенами по всему миру.
Но из всех систем классификации, какие можно выдумать, только одна является уникальной в том смысле, что, располагая полной информацией, к этой системе абсолютно правомерно применять такие понятия, как “правильно” и “неправильно”, “истинно” и “ложно”. Эта единственная в своем роде классификация строится на эволюционном родстве организмов. Во избежание путаницы я буду называть эту систему так, как биологи называют самую строгую из ее разновидностей: кладистическая таксономия.
В кладистической таксономии главным критерием распределения организмов по группам служит степень их родства или, другими словами, давность их происхождения от общего предка. Например, птицы отличаются от нептиц тем, что все они имеют такого общего предка, который не приходится предком никому, кроме птиц. Все млекопитающие тоже происходят от общего предка, более ничьим предком не являющегося. У птиц с млекопитающими был более далекий общий предок, который также дал начало и многим другим животным — например, змеям, ящерицам и гаттериям. Всех животных, произошедших от этого общего предка, называют амниотами. Таким образом, птицы и млекопитающие — амниоты. “Пресмыкающиеся”, с точки зрения кладистов, — ненастоящая таксономическая группа, потому что она определяется методом исключения: все амниоты, кроме птиц и млекопитающих. Иначе говоря, ближайший общий предок всех “рептилий” (змей, черепах и т. д.) является предком и некоторых других животных, а именно птиц и млекопитающих.
Внутри класса млекопитающих относительно недавно жили общие предки мышей и крыс, леопардов и львов, шимпанзе и людей. Близкородственными называют тех животных, чей общий предок жил недавно. Чем меньше степень родства, тем раньше жил общий предок. Совсем дальние родственники, такие как люди и слизни, имеют очень далекого общего предка. Организмы не могут быть совсем не родственными друг другу, поскольку практически наверняка жизнь в том виде, в каком мы ее знаем, зарождалась на земле всего однажды.
Подлинная кладистическая таксономия является строго иерархической — это означает, что она может быть представлена в виде ветвящегося дерева, ветви которого никогда не сходятся вновь. По моему мнению (не разделяемому некоторыми из тех таксономических школ, которые мы вскоре обсудим), ее строгая иерархичность связана не с тем, что иерархические классификации удобны, как они удобны в библиотеках, и не с тем, что все в мире имеет свою естественную иерархию, а только лишь с тем, что эволюционное происхождение осуществляется по иерархическому принципу. Стоит двум новым ветвям древа жизни разойтись на некоторое минимальное расстояние (превышающее хотя бы рамки одного вида), и они уже никогда и ни при каких обстоятельствах не сойдутся снова. (У этого правила могут быть редчайшие исключения — например, упоминавшееся в главе 7 возникновение эукариотической клетки.) Птицы и млекопитающие происходят от общего предка, но теперь это различные ветви эволюции, которые уже никогда не объединятся: гибрид между птицей и млекопитающим невозможен. Группа организмов, произошедшая от общего предка, не приходящегося предком никому из представителей других групп, называется клада, что по-гречески означает “ветвь”.
Эту идею строгой иерархии можно представить и по-другому — в понятиях “идеальной вложенности”. Мы можем взять большой лист бумаги, выписать на него названия любых групп животных и обвести кружком названия близкородственных групп. Например, крыса и мышь будут объединены в рамках одного маленького кружочка, обозначающего, что это близкие родственники, чей общий предок жил недавно. Другой такой маленький кружок объединит вместе морскую свинку и капибару. Эти два кружка — крыса/мышь и морская свинка / капибара — попадут, в свою очередь, в один более крупный общий кружок, имеющий свое собственное название “грызуны”, где им составят компанию бобры, дикобразы, белки и многие другие животные. Маленькие внутренние кружки как бы “вложены” в более крупные внешние. Где-то на другом участке того же листа бумаги в одном небольшом кружке будут объединены тигр и лев. Этот кружок вместе со многими другими будет включен в состав круга, называемого “кошки”. Кошки, собаки, хорьки, медведи и т. п. будут образовывать сложную систему вложенных друг в друга окружностей, но все они попадут в один большой круг под названием “хищные”. Два больших круга, грызунов и хищников, составят часть системы более крупных окружностей внутри очень большого круга, который называется “млекопитающие”.
Получившаяся схема из кругов обладает важной особенностью: она является идеально вложенной. Никогда, ни в каком из случаев нарисованные нами окружности не пересекаются друг с другом. Это правило, не знающее исключений. Если два круга перекрываются, значит, один полностью расположен внутри другого, и никак иначе. Область, ограниченная внутренней окружностью, всегда целиком входит в состав внешнего круга. Частичного перекрывания быть не может. Это свойство идеальной таксономической вложенности не характерно ни для книг, ни для языков, ни для типов почвы, ни для философских течений. Библиотекарь, который в один круг поместит книги по биологии, а в другой — книги по теологии, обнаружит, что два эти круга частично перекрываются. В зону перекрытия попадут книги с заголовками типа “Биология и христианское вероучение”.
На первый взгляд, свойством идеальной вложенности должна обладать и классификация языков. Как мы уже видели в главе 8, эволюция языков во многом напоминает эволюцию животных. Языки, которые недавно произошли от общего предка, такие как, например, шведский, норвежский и датский, имеют намного больше сходства друг с другом, нежели с языками, ответвившимися раньше — например, с исландским. Однако языкам свойственно не только расходиться, но и соединяться. Современный английский язык возник как гибрид между германскими и романскими языками, разошедшимися много раньше, и следовательно, будет непросто аккуратно разместить его на диаграмме иерархического ветвления с идеальной вложенностью. Окружности, внутрь которых попадет английский, будут пересекаться, частично накладываться друг на друга. А в биологической классификации такие перекрывания невозможны, поскольку на уровнях выше видового биологическая эволюция всегда является дивергентной.
Но вернемся к примеру с библиотекой. Никакому библиотекарю не удастся полностью избежать проблемы промежуточных форм или “пересечений”. И тут бессмысленно располагать биологический и теологический отделы в соседних помещениях, а “промежуточные” книги — в соединяющем эти помещения коридорчике. Куда тогда прикажете девать книги, посвященные вопросам на стыке биологии и химии, физики и теологии, истории и теологии, истории и биологии? Думаю, я не ошибусь, если скажу, что проблема промежуточных форм является неизбежной, неотъемлемой частью любой таксономической системы за исключением тех, которые выводятся из эволюционной биологии. Что касается меня, то эта проблема причиняет мне почти физические страдания, когда в моей профессиональной жизни в весьма скромных масштабах возникает необходимость привести что-либо в порядок — расставить свои личные книги и систематизировать те копии научных статей, что мне присылают (руководствуясь самыми добрыми намерениями) мои коллеги, разобрать документы, старые письма и т. п. Какую совокупность категорий ни придумай, всегда найдутся странные предметы, которые не будут подходить никуда, и вынужден с огорчением признаться: из-за моей досадной нерешительности такие бумаги остаются на столе, где иногда валяются годами — до тех пор пока их нельзя со спокойной душой выбросить. В качестве не слишком удачного выхода из положения можно создать для них категорию “разное” — категорию, которая, только создай ее, начнет разрастаться с угрожающей быстротой. Я иногда задаюсь вопросом: не страдают ли библиотекари и хранители коллекций в музеях (за исключением биологических музеев) язвенной болезнью чаще прочих?
При систематике живых организмов таких проблем не возникает. Не бывает животных из категории “разное”. Пока мы остаемся на уровне более высоком, чем вид, и пока мы изучаем только современных животных (или животных, взятых в любой другой конкретный момент времени, — см. ниже), неудобных промежуточных форм быть не может. Если животное выглядит как неудобная промежуточная форма — например, как нечто среднее между млекопитающим и птицей, — то эволюционист может быть уверен в том, что на самом деле это наверняка либо одно, либо другое. Наблюдаемая неопределенность непременно окажется иллюзорной. А несчастному библиотекарю неоткуда взять такую уверенность. Книга вполне может быть и биологической, и исторической одновременно. Биологи с кладистическим образом мыслей никогда не позволят себе рассуждений библиотекарского типа о том, кем “удобнее” считать китов: млекопитающими, рыбами или чем-то средним. Удобство здесь ни при чем; единственное, что имеет значение, — это факты. Факты же приводят всех современных биологов к одному и тому же заключению. Киты — это млекопитающие, а не рыбы, и они ни в малейшей степени не являются чем-либо промежуточным. К рыбам они ничуть не ближе, чем люди, утконосы или любые другие млекопитающие.
Действительно, важно понимать, что все млекопитающие — люди, киты, утконосы и прочие — в точности одинаково родственны рыбам, поскольку все млекопитающие связаны с рыбами родством через одного и того же общего предка. То, что млекопитающие якобы образуют “лестницу”, где “низшие” ближе к рыбам, чем “высшие”, — миф, имеющий отношение к снобизму, а не к эволюции. Это старинное, еще доэволюционистское понятие, называемое иногда “великой цепью бытия”, должно было быть уничтожено эволюционной теорией, но вместо этого таинственным образом проникло в представления многих людей об эволюции.
Тут я не могу совладать с искушением и не обратить внимание на то, какая ирония заключается в том вызове, который креационисты столь охотно бросают эволюционистам: “Предъявите ваши промежуточные формы! Если эволюция — правда, тогда должны существовать животные, представляющие собой нечто среднее между кошкой и собакой или между лягушкой и слоном. Но разве кто-нибудь встречал когда-нибудь лягушкослона?” Я не раз получал по почте креационистские брошюры, высмеивающие эволюцию при помощи карикатур с изображением гротескных химер, составленных из частей разных животных — например, задняя часть тела лошади была приставлена к передней части тела собаки. По всей видимости, авторы этих брошюр думают, будто эволюционисты предполагают существование таких промежуточных форм. Это не просто неправильное понимание истины — это ее антитеза. Один из самых убедительных выводов из эволюционной теории заключается в том, что подобные промежуточные формы не долж ны существовать. В этом была суть моего сравнения животных с библиотечными книгами.
Итак, систематика живых существ, возникших путем эволюции, обладает уникальным свойством: в лучшем из миров, где мы владели бы всей полнотой информации, она не вызывала бы никаких разногласий. Именно это я имел в виду, когда говорил, что к утверждениям кладистической таксономии применимы такие понятия, как “истинность” или “ложность”, неприменимые в библиотечной систематике. Здесь необходимо сделать две оговорки. Во-первых, в действительности мы полной информацией не располагаем. Биологи могут расходиться во взглядах на родословную живых организмов, и нехватка данных — например, из-за недостаточного количества ископаемых — может приводить к ожесточенным спорам. К этой проблеме мы еще вернемся. Вторая же проблема, напротив, может возникнуть при наличии избытка ископаемых. Четкая и аккуратная дискретность классификации рассеется как дым, если мы попытаемся вместить в нее всех животных, которые когда-либо существовали, а не только современных. Связано это с тем, что, какими бы дальними родственниками ни были два современных животных, скажем птица и млекопитающее, давным-давно у них все же был общий предок. И найти ему место в нашей современной таксономии может оказаться непростой задачей.
Как только мы начинаем учитывать вымерших животных, утверж дение об отсутствии промежуточных форм сразу же перестает быть верным. Напротив, теперь мы оказываемся один на один с потенциально непрерывным рядом промежуточных форм. Разграничение между современными птицами и современными нептицами — например, млекопитающими — является четким только лишь потому, что все промежуточные формы, ведущие назад к их общему предку, мертвы. Для большей наглядности я попрошу вас снова представить себе, что природа оказалась вдруг “благосклонна” к нам и обеспечила нас полной палеонтологической летописью, сохранив ископаемые останки всех когда-либо живших животных. Впервые воспользовавшись этой выдумкой в предыдущей главе, я упомянул, что в одном отношении такая природа на самом деле оказалась бы отнюдь не любезной. Тогда я подразумевал тяжкий труд по изучению и описанию всех ископаемых, который пришлось бы проделать, но теперь нам предстала еще одна грань этой парадоксальной нелюбезности. Полная геологическая летопись сделала бы крайне затруднительным распределение животных по обособленным систематическим группам, которым можно было бы дать названия. Обладай мы таким “подарком” природы, нам пришлось бы отказаться от названий для конкретных групп и прибегнуть к каким-то математическим или графическим обозначениям плавно меняющихся параметров. Человеческий разум намного охотнее имеет дело с отдельными наименованиями, так что в некотором смысле даже хорошо, что палеонтологическая летопись столь бедна.
Если мы будем рассматривать всех когда-либо живших, а не только современных животных, то такие понятия, как “человек” или “птица”, станут столь же расплывчатыми и неточными, как понятия “высокий” или “толстый”. Палеонтологи могут до бесконечности спорить о том, считать им то или иное ископаемое птицей или нет. Именно этот вопрос, в самом деле, часто поднимается вокруг знаменитого ископаемого Archaeopteryx. Выходит, что различие между птицей и нептицей четче различия между большим и маленьким только потому, что в первом из этих случаев все неудобные промежуточные формы вымерли. Если бы какая-нибудь отличающаяся странной избирательностью эпидемия вдруг выкосила всех до единого людей среднего роста, тогда понятия “высокий рост” и “низкий рост” стали бы такими же определенными, как “птица” или “млекопитающее”.
Тот удобный факт, что большинство промежуточных форм вымерли, спасает от затруднительной неопределенности не только классификацию живых организмов, но также человеческую этику и законодательство. Наша юридическая система и наши представления о нравственности жестко ограничиваются видовыми рамками. Директор зоопарка имеет право “списать” лишнего шимпанзе, в то время как попытка избавиться от ненужного сторожа или билетера может быть воспринята как возмутительный произвол. Шимпанзе — собственность зоопарка. А людей в наши дни не считают чьей-либо собственностью, хотя редко кто удосуживается разумно обосновать такую дискриминацию — лично я сомневаюсь, что тут вообще может быть какое-либо логическое обоснование. “Видизм” нашего исповедующего христианские ценности общества просто ошеломляет: аборт одной-единственной человеческой зиготы (большинству из которых суждено погибнуть в любом случае) может вызвать больше душевных терзаний и благородного негодования, чем вивисекция любого количества смышленых взрослых шимпанзе! Я знаю достойных, либерально мыслящих ученых, которые на самом деле не собираются резать живых шимпанзе, но тем не менее с пеной у рта отстаивают свое право делать это, если им вздумается, без каких-либо препятствий со стороны закона. Эти же люди часто первыми приходят в ярость при малейшем посягательстве на права человека. Причина, по которой наша совесть мирно уживается с такими двойными стандартами, заключается только в том, что промежуточных форм между человеком и шимпанзе не осталось.
Последний общий предок людей и шимпанзе жил, судя по всему, около 5 млн лет назад, то есть определенно позже, чем общий предок шимпанзе и орангутанов и, вероятно, на 30 млн лет позже общего предка шимпанзе и всех прочих обезьян. Более 99 % генов у нас с шимпанзе идентичны. Если бы на каком-нибудь забытом острове вдруг обнаружилась сохранившаяся промежуточная форма, восходящая к общему предку шимпанзе и человека, то можно не сомневаться: наши законы и нравственные нормы подверглись бы тяжелому испытанию — особенно если бы на каком-либо участке имеющегося спектра форм сохранилась возможность для скрещивания между разновидностями. Пришлось бы либо даровать права человека всем представителям спектра (избирательное право для шимпанзе!), либо разработать сложную систему апартеида с дискриминирующими законами, где суды разбирали бы вопрос, считать того или иного индивидуума “юридически шимпанзе” или “юридически человеком”, а родители беспокоились, как бы их дочь не вышла замуж за одного из “этих”. Наш мир, как я подозреваю, достаточно хорошо изучен, чтобы человечество могло не бояться такого заслуженного наказания. Но каждому, кто считает, будто человеческие “права” — это что-то очевидное и само собой разумеющееся, стоит задуматься о той случайной удаче, какой оказалось для нас отсутствие смазывающих четкую картину промежуточных форм. Впрочем, возможно, что, если бы шимпанзе были открыты сегодня, они показались бы нам промежуточной формой и создали бы затруднения.
Те, кто читал предыдущую главу, могут заметить, что вся эта аргументация насчет того, что категории не будут размытыми только в том случае, если классификация ограничится современными животными, основана на допущении, что эволюция не прерывиста, а идет с постоянной скоростью. Чем более плавным, постепенным представляется нам ход эволюции, тем более безнадежной нам должна казаться сама возможность применять такие понятия, как “птица” и “нептица”, “человек” и “нечеловек” по отношению ко всем когда-либо жившим животным. А крайний сальтационист, напротив, мог бы считать, что на свете действительно существовал самый первый человек, чей мутантный мозг был вдвое больше как мозга его отца, так и мозга его брата-шимпанзе.
Как мы видели, сторонники теории прерывистого равновесия в большинстве своем не являются истинными сальтационистами. Тем не менее проблема неопределенности названий поневоле должна быть для них менее затруднительной, чем для тех, кто придерживается не столь радикальных взглядов на эволюцию. Если бы буквально каждое животное сохранялось в ископаемом виде, тогда проблема с наименованиями категорий встала бы даже перед пунктуалистами, ведь на самом-то деле они градуалисты, стоит только как следует разобраться. Однако, поскольку они исходят из того предположения, что вероятность найти ископаемые свидетельства кратких переходных периодов чрезвычайно мала, а ископаемые, оставшиеся от долгих периодов стазиса, напротив, обнаруживаются относительно легко, “проблема названий” все же не должна стоять перед пунктуалистами так остро, как перед теми, кто не считает эволюцию прерывистой.
Именно по этой причине пунктуалисты, особенно Найлз Элдридж, придают большое значение тому, чтобы считать понятие “вид” реальной “сущностью”. Для того, кто не является пунктуалистом, это понятие поддается определению лишь постольку, поскольку неудобных промежуточных форм более не существует. Если крайний антипунктуалист посмотрит со своей точки зрения на всю эволюционную историю в целом, то он вообще не увидит там никаких обособленных “видов”, один только смазанный континуум. По его мнению, у вида не может быть четко определяемого начала и только иногда бывает четко определяемый конец (вымирание); зачастую же вид не пресекается, а потихоньку превращается в новый. Зато пунктуалисту все представляется так, будто каждый вид возникает в определенный момент времени (строго говоря, имеется переходный период, длящийся несколько десятков тысяч лет, но по геологическим меркам это недолго). Более того, он считает, что у видов должно быть и четкое (ну или по крайней мере скоротечное) завершение, а не постепенный переход в другой вид. Поскольку с пунктуалистской точки зрения почти вся жизнь вида проходит в неизменном состоянии стазиса и поскольку у видов имеются обособленные начало и конец, то для пунктуалиста правомерно говорить об определенной и поддающейся измерению “продолжительности жизни” вида. А с непунктуалистской точки зрения понятие “продолжительность жизни” к видам, в отличие от индивидуальных организмов, применяться не может. Для крайнего пунктуалиста слово “вид” означает нечто реально существующее и вполне достойное иметь свое собственное название. А для крайнего антипунктуалиста “вид” — это произвольно выбранный участок непрерывно текущей реки, и нет никаких особых оснований считать какую-либо точку его началом или концом.
В написанной с пунктуалистских позиций книге по эволюционной истории некоей группы животных — например, по истории лошадей за последние 30 млн лет — действующими лицами вполне могут быть не индивидуальные организмы, а виды, поскольку с точки зрения автора-пунктуалиста вид является реально существующим “объектом”, который обладает выраженной индивидуальностью. Виды будут внезапно выходить на сцену и внезапно исчезать, уступая место видам-преемникам. Это будет история династии видов, поочередно сменяющих друг друга. Но если ту же самую историю будет писать антипунктуалист, то он будет употреблять видовые названия исключительно ради некоего смутного удобства. Рассматривая шкалу времен по всей ее длине, он перестает различать виды как обособленные единицы. Настоящими героями драмы в книге этого второго автора будут отдельные животные внутри меняющихся популяций. Особи будут уступать место новым особям, а не виды — новым видам. Так что вряд ли стоит удивляться, что пунктуалисты склонны верить в существование своеобразного естественного отбора на видовом уровне, аналогичного традиционному дарвиновскому отбору, действующему на уровне индивидуальных организмов. А антипунктуалисты, напротив, скорее всего, будут рассматривать действие естественного отбора на уровне особи, но не выше. Идея “межвидового отбора” меньше придется им по вкусу, поскольку они не воспринимают виды как нечто обособленно существующее в геологических временны´х масштабах.
Подошел удачный момент, чтобы коснуться гипотезы межвидового отбора — темы, фактически оставшейся нам в наследство от предыдущей главы. Я не стану уделять ей много времени — мои сомнения насчет способности межвидового отбора играть значимую роль в эволюции подробно разъяснены в “Расширенном фенотипе”. Подавляющее большинство когда-либо существовавших видов вымерли — это бесспорно. Справедливо и то, что интенсивность возникновения новых видов по меньшей мере компенсирует вымирание, благодаря чему мы имеем некий “видофонд”, состав которого постоянно меняется. Неслучайное попадание видов в этот “фонд” и неслучайное выбывание их оттуда теоретически могли бы в самом деле дать начало своеобразному естественному отбору высшего порядка. Также вполне возможно, что какие-то характерные особенности видов оказывают влияние на вероятность их вымирания и на их способность давать начало новым видам. Те виды, что уже существуют, должны быть склонны к обладанию такими качествами, которые позволили им, прежде всего, вообще возникнуть — “видообразоваться”, — равно как и такими, которые не дают им вымереть. При желании это можно назвать разновидностью естественного отбора, хотя я подозреваю, что отбор этот скорее одноступенчатый, нежели накапливающий. Единственное, в чем я здесь сомневаюсь, — это то, что отбор такого рода может быть хоть сколько-нибудь важен для понимания эволюционных процессов.
Может быть, у меня просто предвзятое мнение насчет того, что считать важным. Как я уже сказал в начале этой главы, мое главное требование к любой эволюционной теории состоит в том, чтобы она объясняла существование таких сложно устроенных механизмов, как сердце, рука, глаз или эхолот. Никакой, даже самый пылкий приверженец гипотезы межвидового отбора ни на что такое не претендует. Некоторые полагают, что межвидовым отбором можно объяснить некоторые долгосрочные тенденции, наблюдаемые в геологической летописи, — например, нередко встречающуюся тенденцию к увеличению средних размеров тела с течением эпох. Мы уже упоминали, что, скажем, современные лошади крупнее, чем их предки, жившие 30 млн лет назад. Видовые селекционисты отказываются считать это следствием того, что преимущества для отдельных особей всегда оставались одними и теми же. Они не рассматривают данную тенденцию как свидетельство неизменно большей успешности крупных лошадей по сравнению с мелкими представителями тех же самых видов. По их мнению, дело было так. Было много различных видов, некий видофонд. У представителей одних видов средний размер тела был крупным, а у представителей других видов — мелким (возможно, в связи с тем, что у каких-то видов преуспевали крупные, а у каких-то — мелкие особи). Но виды, представленные крупными организмами, были менее склонны к вымиранию (или более склонны к отпочковыванию новых, себе подобных видов). В соответствии с такой точкой зрения, какие бы процессы ни происходили внутри видов, ископаемая летопись будет показывать тенденцию к увеличению размера тела, возникшую благодаря сменяющей друг друга последовательности видов, представленных все более и более крупными животными. Возможно даже, что у большинства видов естественный отбор благоприятствовал более мелким особям, но даже в этом случае палеонтологические данные будут показывать всю ту же тенденцию к увеличению. Иначе говоря, межвидовой отбор мог благоприятствовать меньшинству тех видов, у которых преимущество имели крупные особи. Именно эта мысль (правда, скорее в форме интеллектуальной забавы) была высказана крупнейшим теоретиком неодарвинизма Дж. К. Уильямсом еще задолго до того, как современные видовые селекционисты появились на научной арене.
Возможно, здесь, да и вообще во всех предполагаемых примерах межвидового отбора мы имеем дело не столько с эволюционным направлением, сколько с направлением с укцессии, напоминающей то, как безжизненную территорию последовательно заселяют все более и более крупные растения: сначала совсем небольшие травянистые формы, затем более заметные, затем кустарники и, наконец, зрелое, “климаксное” сообщество — лес. Так или иначе, видовые селекционисты вполне могут быть правы, считая, что подобные тенденции — как их ни называй, эволюционными или сукцессионными, — часто встречаются палеонтологам при изучении следующих друг за другом геологических пластов. Но, как я уже заметил, никто не собирается утверждать, будто межвидовой отбор представляет хоть какую-то важность для нашего понимания эволюции сложных приспособлений. И вот почему.
Сложные приспособления являются по большей части свойствами особей, а не видов. У видов не бывает ни глаз, ни сердец, но они могут быть у особей, принадлежащих к этим видам. Если вид вымирает по причине слабого зрения, то это, скорее всего, подразумевает гибель каждой особи из-за того, что она плохо видела. Острота зрения — признак индивидуальных организмов, а не видов. А какие свойства можно было бы считать именно видовыми? Долж но быть, те, которые влияют на выживаемость и размножение вида таким образом, что это не может быть сведено к совокупности их влияний на выживаемость и размножение особей. В нашем гипотетическом примере с лошадьми я высказал предположение, что те немногие виды, у которых естественный отбор благоприятствовал более крупным особям, вымирали реже тех многих видов, у которых индивидуальное преимущество имели более мелкие особи. Но все это не слишком убедительно. Трудно представить себе, с какой стати выживаемость вида должна быть не связана с суммарной выживаемостью его представителей.
Удачнее будет другой гипотетический пример признака, относящегося к видовому уровню. Предположим, все представители какого-то вида добывают себе пропитание из одного и того же источника. Например, коалы живут на эвкалиптовых деревьях и не питаются ничем, кроме эвкалиптовых листьев. Такой вид можно назвать однородным. А какой-то другой вид представлен непохожими друг на друга особями, по-разному добывающими себе пропитание. Каждый отдельно взятый индивид вполне может быть так же узкоспециализирован, как и отдельно взятая коала, но у вида в целом имеется целый спектр пищевых привычек. Одни его представители питаются только листьями эвкалипта, другие только пшеницей, третьи только ямсом, четвертые только кожурой лайма и так далее. Назовем такой вид неоднородным. Думаю, нетрудно вообразить себе обстоятельства, при которых однородные виды могут вымереть с большей вероятностью, чем неоднородные. Коалы всецело зависят от наличия эвкалиптов, и какая-нибудь эпидемия среди эвкалиптовых деревьев, аналогичная голландской болезни вяза, вполне может уничтожить и коал. А вот у разнородного вида при эпидемии, которая поразит любое из употребляемых в пищу растений, какие-то представители все равно выживут, и вид продолжит свое существование. Нетрудно также предположить, что неоднородные виды смогут легче по сравнению с однородными давать начало новым, дочерним видам. Возможно, тут и найдутся какие-то примеры подлинного межвидового отбора. В отличие от близорукости и длинноногости “однородность” и “неоднородность” — это свойства, действительно относящиеся к видам. Беда в том, что примеры таких свойств крайне немногочисленны.
Американский эволюционист Эгберт Ли выдвинул интересную теорию, которую можно рассматривать как гипотетический пример настоящего межвидового отбора, хотя она и была предложена еще до того, как словосочетание “межвидовой отбор” вошло в моду. Ли занимался вечной проблемой — эволюцией “альтруистического” поведения индивидуальных организмов. Он справедливо признавал, что в случае если интересы особи вступят в конфликт с интересами вида, то первые — краткосрочные, индивидуальные интересы — непременно возобладают. По-видимому, ничто не способно остановить неумолимую поступь эгоистичных генов. Однако Ли сделал следующее любопытное предположение. Должны существовать такие виды, у которых случайно вышло так, что то, что хорошо для особи, в немалой степени совпадает с тем, что хорошо для вида. При этом должны существовать и другие виды, у которых интересы особей и интересы вида случайно оказались особенно несовпадающими друг с другом. Вполне возможно, что при прочих равных условиях виды из второй категории будут склонны к вымиранию с большей вероятностью. Тогда некая форма межвидового отбора будет благоприятствовать — нет, не личному самопожертвованию, но тем видам, от представителей которых не требуется приносить в жертву свое собственное благополучие. И мы можем наблюдать формирование явно неэгоистичного поведения особей, поскольку межвидовой отбор благоприятствовал таким видам, где выгода индивидуальных организмов лучше всего достигается за счет их кажущегося альтруистическим поведения.
Самый впечатляющий пример свойства, о котором можно говорить именно на уровне вида, связан, вероятно, со способом размножения: половое или бесполое. По причинам, в которые у меня нет возможности здесь углубляться, существование полового размножения создает серьезную теоретическую головоломку для дарвинистов. Много лет назад Р. Э. Фишер, обычно враждебный к любым идеям о возможности отбора на уровне более высоком, чем индивидуальный организм, все же был готов сделать исключение для раздельнополости как таковой. Он утверждал, что по причинам, в которые я опять-таки не буду вдаваться (они не так очевидны, как можно было бы подумать), виды, размножающиеся половым путем, способны эволюционировать быстрее видов, не имеющих полового размножения. Эволюция — это то, что происходит с видом, а не с особью: нельзя рассматривать организм как нечто эволюционирующее. Таким образом, согласно гипотезе Фишера, отбор на уровне видов отчасти является причиной того, почему половое размножение столь распространено среди современных нам животных. Но, даже если и так, мы здесь имеем дело с примером не накапливающего, а одноступенчатого отбора.
В соответствии с этими рассуждениями виды, не имеющие полового размножения, возникнув, склонны к вымиранию, поскольку они эволюционируют недостаточно быстро для того, чтобы соответствовать меняющейся окружающей среде. А виды, которые размножаются половым путем, имеют обыкновение выживать, так как скорость их эволюции может быть достаточно высокой. Вот почему мы видим вокруг себя главным образом организмы с половым размножением. Однако та “эволюция”, скорость которой различна у тех и у других, — это, разумеется, все та же обычная дарвиновская эволюция, движимая накапливающим отбором на уровне индивидуумов. Описываемый здесь межвидовой отбор является одноступенчатым — это простейший выбор между двумя признаками: отсутствием и наличием полового процесса, медленной эволюцией и быстрой эволюцией. Все обеспечивающие половое размножение механизмы: половые органы, половое поведение, аппарат деления половых клеток — все это должно было быть собрано и отлажено действующим на низком уровне традиционным накапливающим отбором по Дарвину, а никак не межвидовым отбором. Как бы то ни было, современные ученые единодушно отвергают старую теорию о том, что существование полового размножения поддерживается некой разновидностью отбора на уровне группы или вида.
Подведу итоги этой дискуссии о межвидовом отборе. С его помощью можно объяснять, почему в некий определенный момент времени землю населяет тот, а не иной набор видов. Отсюда следует, что межвидовым отбором в принципе можно объяснять и изменение видового состава по мере того, как одни геологические эпохи уступают место другим — иначе говоря, изменения, происходящие с палеонтологической летописью. Но межвидовой отбор не является существенной силой при эволюции сложных механизмов живого. Самое большее, на что он способен, — это выбирать между несколькими вариантами сложных механизмов, уже созданных настоящим дарвиновским отбором. Как я указывал прежде, межвидовой отбор может существовать, но делает он, по всей видимости, немного! Теперь давайте вернемся к систематике и ее методам.
Я сказал, что кладистическая таксономия обладает преимуществом перед классификациями библиотечного типа в том смысле, что в природе имеется одна-единственная, истинно иерархическая модель вложенности, которая ждет, когда ее откроют. Все, что от нас требуется, — это разрабатывать методы для ее выявления. К сожалению, тут мы сталкиваемся с практическими трудностями. Главной головной болью систематиков является эволюционная конвергенция. Это настолько замечательный феномен, что я уже посвятил ему половину главы 4. Там мы могли видеть, как снова и снова в разных частях света биологи обнаруживают животных, похожих благодаря сходству в образе жизни на неродственных им животных с других континентов. Муравьи-легионеры Нового Света напоминают муравьев-кочевников Старого Света. В ходе эволюции возникло поразительное сходство между имеющими весьма отдаленное родство электрическими рыбами Африки и Южной Америки, а также между настоящими волками и тасманийским сумчатым волком Thylacinus. Приводя все эти примеры, я просто утверждал без каких-либо пояснений, что речь идет о конвергентном сходстве, выработанном неродственными видами животных независимо друг от друга. Но откуда мы знаем, что эти виды не родственны? Если для определения степени родства организмов ученые пользуются их внешним сходством, то почему же такие на удивление общие признаки, которые, казалось, должны были объединить каждую из вышеперечисленных пар, не смогли обмануть систематиков? Или поставим этот вопрос в еще более неудобной форме: когда систематики говорят нам, что два вида — скажем, зайцы и кролики — действительно являются близкими родственниками, откуда нам знать, что это не обманчивая конвергенция, которая зашла очень далеко?
Вопрос в самом деле неприятный, поскольку история таксономии изобилует случаями, когда ученые-систематики заявляли, что их предшественники заблуждались, именно по этой причине. Как мы помним из главы 4, один аргентинский систематик утверждал, будто литоптерны являются предками настоящих лошадей, в то время как теперь сходство литоптерн с лошадьми объясняют конвергенцией. Африканский дикобраз долгое время считался близким родственником американских дикобразов, но теперь полагают, что две эти группы организмов обзавелись своей колючей шубой независимо друг от друга. По-видимому, колючки служат и тем и другим для одних и тех же целей на обоих континентах. Кто, однако, может гарантировать, что следующие поколения систематиков снова не переменят свою точку зрения на противоположную? Как мы можем полагаться на классификацию, когда конвергентная эволюция — такой мастер убедительных подделок? Лично я смотрю на эти проблемы с оптимизмом главным образом потому, что на сцену вышли новые и эффективные методы молекулярной биологии.
Вернемся на минутку к содержанию предыдущих глав. Все животные, растения и бактерии, как бы ни были они на первый взгляд непохожи друг на друга, оказываются, если добраться до молекулярных основ, на удивление единообразными. Ярче всего это можно увидеть на примере генетического кода как такового. Словарь генов состоит из 64 слов, по три буквы в каждом. Каждое из этих слов имеет точный перевод на язык белков, обозначая либо какую-то определенную аминокислоту, либо знак препинания. Этот язык является, по всей видимости, произвольным в том же смысле, в каком произвольны и человеческие языки (в звучании слова “дом”, например, нет ничего такого, что само по себе могло бы навести слушателя на мысль о жилище). И потому нам представляется чрезвычайно красноречивым тот факт, что все живые существа, сколь бы ни отличались они друг от друга своим внешним видом, “разговаривают” практически абсолютно на одном и том же генетическом языке. Генетический код универсален. Я считаю это едва ли не окончательным доводом в пользу того, что все организмы происходят от одного-единственного общего предка. Один и тот же словарь произвольно установленных “значений слов” не мог возникнуть дважды — вероятность такого события невообразимо мала. В главе 6 мы говорили о том, что некогда могли существовать организмы, пользовавшиеся другим генетическим языком, но их, увы, больше нет с нами. Все уцелевшие живые существа произошли от одного общего предка и унаследовали от него практически идентичный, хотя и абсолютно произвольный, генетический словарь с почти что одним и тем же набором из 64 ДНК-слов.
Только подумайте, какое значение это может иметь для систематики! До наступления молекулярно-биологической эры зоологи могли быть уверены во взаимном родстве только тех животных, которые имели очень большое количество сходных анатомических признаков. А молекулярная биология внезапно обнаружила целый Клондайк общих признаков в дополнение к тому скудному списку, на какой нам позволяли рассчитывать анатомия и эмбриология. Шестьдесят четыре идентичные (слово “сходные” представляется тут слишком слабым) особенности общего генетического словаря — это только цветочки. Систематика преобразилась раз и навсегда. Слабые догадки относительно родственных взаимоотношений тех или иных организмов превратились в практически полную статистическую достоверность.
Почти что стопроцентная универсальность генетического словаря — подарок, с точки зрения систематиков, слишком щедрый. Она доказывает нам, что все живые существа приходятся друг другу родней, но ничего не говорит о том, какие из них состоят в более близком родстве друг с другом. Однако это мы можем узнать из других молекулярных данных, где имеют место различные степени сходства, а не полная идентичность. Напомню, что продуктом аппарата трансляции генов являются белки. Каждая белковая молекула — это предложение, последовательность аминокислотных слов из того самого словаря. Мы имеем возможность расшифровать эти предложения — как в переводе на белковый язык, так и в первоначальной форме последовательности ДНК. Хотя все формы жизни пользуются одним и тем же словарем, они составляют с его помощью отнюдь не одинаковые предложения. И это дает нам возможность рассчитать степень родства между организмами. Белковые “предложения” разных видов нередко имеют более-менее сходную структуру, различаясь при этом в деталях. У любой пары организмов можно найти предложения, достаточно похожие друг на друга для того, чтобы в них можно было узнать различные “искаженные” варианты одного и того же исходного предложения. Мы уже встречались с этим явлением на примере незначительных различий между последовательностями гистонов у коровы и гороха.
Теперь систематики имеют возможность сравнивать между собой молекулярные предложения так же, как они сравнивают черепа или кости конечностей. Предложения из белков или ДНК, обладающие значительным сходством, можно считать принадлежащими близкородственным видам, а предложения с бóльшим числом различий — относящимися к более дальним родственникам. Все эти предложения построены на основе единого для всех словаря, состоящего всего-навсего из 64 слов. Прелесть современной молекулярной биологии заключается в том, что она позволяет вычислить дальность родства между двумя организмами точно — то есть в виде конкретного числа, равного количеству различающихся слов в двух версиях одного и того же предложения. Если вспомнить генетическое гиперпространство из главы 3, то теперь мы можем измерить точное число шагов, отделяющих один организм от другого, — по крайней мере в том, что касается какой-то определенной молекулы белка.
Дополнительное преимущество изучения молекулярных последовательностей в систематике состоит в том, что, если верить “нейтралистам” — представителям влиятельного направления в генетике, с которым мы еще встретимся в следующей главе, — большинство эволюционных изменений, происходящих на молекулярном уровне, нейтральны. То есть они возникают не благодаря естественному отбору, а фактически случайно, и потому, если отбросить вероятность чрезвычайного невезения, в данном случае конвергенция не будет морочить ученым головы. Кроме того, как мы уже видели, молекулы, принадлежащие к одному типу, эволюционируют, судя по всему, примерно с одинаковой скоростью. А это значит, что количество различий между сопоставимыми молекулами двух животных — например, между человеческим цитохромом и цитохромом бородавочника — хороший показатель того, сколько времени прошло с тех пор, когда жил общий предок этих организмов. Мы располагаем довольно-таки точными молекулярными часами. Эти часы позволяют нам не только выяснить то, какие из организмов имеют наименее далекого общего предка, но также и узнать, когда приблизительно этот предок жил.
Здесь читатель может быть озадачен кажущимся противоречием. На протяжении всей этой книги подчеркивалось первостепенное значение естественного отбора. Как же мы можем теперь утверждать, что на молекулярном уровне эволюционные преобразования случайны? Предвосхищая главу 11, скажу, что, когда речь идет о приспособлениях — главном предмете настоящей книги, — никакого повода для споров в действительности нет. Даже самый ярый нейтралист не думает, будто такие сложные органы, как рука или глаз, возникли путем случайного дрейфа. Любой находящийся в здравом уме биолог согласится с тем, что они были сформированы естественным отбором. Но при этом нейтралисты полагают — и, думаю, они правы, — что адаптации, подобные этим, представляют собой только верхушку айсберга, а большинство эволюционных изменений, происходящих на молекулярном уровне, возможно, не имеют никаких функциональных последствий.
При условии что молекулярные часы реально существуют — а судя по всему, каждый тип молекул действительно претерпевает примерно одно и то же количество изменений на миллион лет, — мы можем воспользоваться ими, чтобы датировать точки ветвления эволюционного древа. А если большинство эволюционных изменений на молекулярном уровне в самом деле нейтральны, то систематикам несказанно повезло. Ведь это означает, что от проблемы конвергенции можно избавиться при помощи такого оружия, как статистика. В клетках у каждого животного записаны огромные массивы текста, который, если верить нейтралистской теории, никоим образом не связан с приспособлениями организма к особенностям его образа жизни и который практически не подвергается ни отбору, ни конвергентной эволюции, если не считать возможности случайных совпадений. Вероятность того, что два больших текстовых фрагмента, не играющих роли при отборе, окажутся похожими друг на друга в силу чистой случайности, можно вычислить, и поверьте, вероятность эта чрезвычайно мала. Замечательнее же всего то, что постоянная скорость молекулярной эволюции позволяет нам теперь датировать моменты расхождения родовых ветвей в эволюционной истории.
Трудно переоценить, насколько новые методы прочтения молекулярных последовательностей расширили арсенал ученых-систематиков. Разумеется, пока еще расшифрованы далеко не все молекулярные “предложения” из геномов всех на свете животных, но уже сейчас мы можем пойти в библиотеку и посмотреть точное — слово к слову, буква к букве — построение фраз в предложениях α-гемоглобина собаки, кенгуру, ехидны, курицы, гадюки, тритона, карпа и человека. Не у всех животных есть гемоглобин, но существуют и другие белки, к примеру гистоны, свой вариант которых имеется у любого животного и растения и многие из которых опять-таки уже можно найти в расшифрованном виде в библиотеке. И это не какие-нибудь расплывчатые параметры вроде длины ноги или ширины черепа, зависящие от возраста и состояния здоровья конкретного экземпляра или даже от глазомера естествоиспытателя. Здесь мы имеем дело с четко сформулированными альтернативными вариантами одного и того же предложения, написанными на одном и том же языке, которые можно сопоставить друг с другом с такой же точностью и подробностью, с какой дотошный специалист по древнегреческому мог бы сравнивать два пергамента с текстом одного и того же евангелия. Последовательности ДНК — это и есть подлинные евангелия всего живого, и мы научились разбираться в них.
Основное положение, из которого исходят систематики, состоит в том, что чем близкородственнее виды, тем более сходными вариантами одного и того же молекулярного текста они будут обладать. Это называется “принципом парсимонии”. Парсимония — то же самое, что и скаредная бережливость. Если у нас есть набор организмов, молекулярные “предложения” которых уже расшифрованы, например восемь животных, перечисленных в предыдущем абзаце, то наша задача выяснить, какая из возможных родословных, объединяющих эти организмы, наиболее экономична. Наиболее экономичным генеалогическим древом считается самое “скупое” на предположения, то есть допускающее минимальное количество эволюционных изменений и минимальный уровень конвергенции. Мы вправе считать конвергенцию минимальной исключительно из соображений вероятности. Крайне неправдоподобно, чтобы два неродственных животных могли случайно прийти к абсолютно одной и той же генетической последовательности — слово в слово, буква к букве, — особенно если предполагать, что значительная часть молекулярной эволюции нейтральна.
Однако при попытке рассмотреть все варианты родословной, какие только возможны, мы сталкиваемся с вычислительными трудностями. Когда нам надо классифицировать только три вида животных, возможных генеалогических деревьев всего-навсего три: A родственно B, а C в стороне; A родственно C, а B в стороне; и наконец, B родственно C, а A в стороне. Аналогичные расчеты можно произвести и для большего числа классифицируемых животных, причем количество возможных комбинаций будет расти экспоненциально. Если нам необходимо классифицировать четыре вида животных, то число альтернативных родословных будет еще приемлемым — всего 15. У компьютера не займет много времени вычислить, которая из 15 самая экономичная. Но если нам придется иметь дело с 20 животными, то количество возможных генеалогических деревьев будет уже совсем другим: я насчитал 8 200 794 532 637 891 559 375 (рис. 9). Чтобы найти наиболее экономичное из них, самому быстрому современному компьютеру понадобится 10 000 млн лет — приблизительный возраст нашей вселенной. А систематикам часто бывает необходимо составить родословную более чем 20 животных.
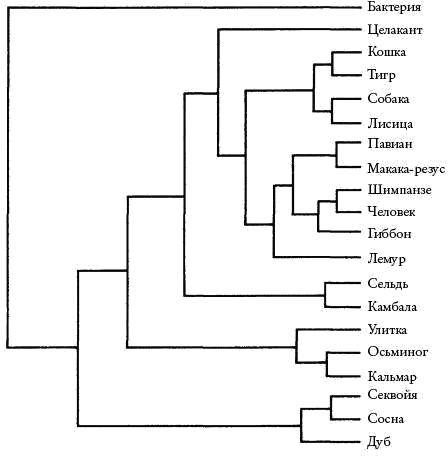
Рис. 9. Это генеалогическое древо является верным. Существует еще 8 200 794 532 637 891 559 374 способа систематизировать данные 20 организмов, и все они неправильные.
Хотя молекулярные систематики первыми стали придавать большое значение проблеме стремительно возрастающих огромных чисел, проблема эта в скрытом виде всегда присутствовала и в традиционной, немолекулярной таксономии. Просто обычные систематики избегали ее при помощи интуитивных догадок. Среди тех генеалогических деревьев, которые теоретически можно было бы рассмотреть, есть такие, которые есть смысл отбросить сразу же, — например, все те миллионы родословных, где люди будут ближе к дождевым червям, чем к шимпанзе. Систематики могут не тратить свое время на рассмотрение таких заведомо абсурдных генеалогических деревьев, а сосредоточиться вместо этого на тех относительно немногочисленных вариантах, которые не столь радикально расходятся с предварительными ожиданиями. Это, вероятно, правомерно, хотя всегда остается опасность того, что самая экономичная версия будет списана со счетов без рассмотрения. Компьютер тоже можно научить рационализировать расчеты, так что, к счастью, проблема больших чисел может оказаться не столь уж велика.
Молекулярная биология снабжает нас таким обилием информации, что мы можем систематизировать организмы снова и снова, каждый раз исходя из сравнения различных белков. Выводы, полученные в результате изучения одних молекул, могут быть использованы для проверки результатов, полученных на других молекулах. И если мы боимся, что в эволюционную историю, рассказанную какой-то одной белковой молекулой, закралась затемняющая истину конвергенция, нам будет нетрудно выяснить, так ли это, взглянув на другую молекулу. Конвергентная эволюция — это, в сущности, особая разновидность совпадения. А совпадение — оно на то и совпадение, чтобы случаться однажды с большей вероятностью, нежели дважды, а дважды — с большей вероятностью, чем трижды. Чем больше молекул белка мы сравниваем, тем ничтожнее возможность случайных совпадений.
Так, например, группа биологов из Новой Зеландии в одном своем исследовании взялась систематизировать 11 видов животных — причем не единожды, а пять раз независимо, с использованием пяти различных белков. Этими животными были овца, макак-резус, лошадь, кенгуру, крыса, кролик, собака, свинья, человек, корова и шимпанзе. Идея была в том, чтобы вначале составить родословную этих 11 видов по одному белку, а затем посмотреть, будет ли она такой же, если использовать другой белок. Теоретически — например, если эволюции не существует, — каждый из этих пяти белков мог бы выявить свою собственную, не похожую на другие картину “родственных взаимоотношений”.
Все последовательности данных пяти белков каждого из 11 видов животных уже имелись в наличии в библиотеках. Для 11 видов существует 654 729 075 возможных вариантов генеалогического древа, и потому пришлось прибегнуть к стандартным приемам рационализации. Компьютер распечатал наиболее экономичные схемы родства, определенные путем независимого сопоставления каждого из пяти белков — пять самых близких догадок относительно того, каким должно быть подлинное генеалогическое древо этих 11 организмов. Идеальным результатом для нас была бы полная идентичность всех пяти получившихся родословных. Вероятность того, что это случайность, была бы в самом деле чрезвычайно мала и равнялась бы числу с 31 нулем после запятой. Однако отсутствие такого полного совпадения вряд ли могло бы сильно нас удивить: определенная доля конвергенции и случайностей неизбежна. Но если бы получившиеся схемы не оказались в существенной степени похожими друг на друга, вот тогда стоило бы забеспокоиться. На самом же деле эти пять генеалогических деревьев получились не полностью идентичными друг другу, но тем не менее почти что одинаковыми. Все пять были единодушны в том, что человека, шимпанзе и резуса следует расположить вместе, но вот насчет того, какое животное считать их самым близким родственником, возникли некоторые разногласия. Гемоглобин B утверждал, что собаку, фибринопептид B — что крысу, фибринопептид A предлагал кластер, состоящий из крысы и кролика, а гемоглобин А — кластер, включающий в себя крысу, кролика и собаку.
У человека и у собаки определенно был некий общий предок; определенно был и общий предок у человека и крысы. Каждый из этих двух предков действительно существовал в какой-то момент эволюционной истории. Какой-то из них непременно должен был жить раньше другого, так что либо гемоглобин B, либо фибринопептид B дает неверную оценку родственных взаимоотношений этих организмов. Как я уже сказал, такие незначительные разногласия не должны нас беспокоить. Определенная доля конвергенции и совпадения вполне ожидаема. Если на самом деле мы более близкие родственники с собакой, это значит, что у нас с крысой произошла конвергенция по фибринопептиду B. А если мы ближе к крысе, то у нас с собакой произошла конвергенция по гемоглобину B. Мы можем выяснить, какое из этих предположений более вероятно, проанализировав еще какие-нибудь молекулы. Но я не буду продолжать эту тему: мысль понятна и так.
Я уже упоминал, что систематика — одна из самых сварливых биологических дисциплин. “Определения и оскорбления” — так метко охарактеризовал ее Стивен Гульд. Систематики демонстрируют ту пылкую приверженность к своим научным школам, какой можно ожидать от политологов или экономистов, но никак не от академических исследователей. Члены какого-то одного таксономического направления явно считают себя братством, осажденным со всех сторон врагами, наподобие первых христиан. Я впервые осознал это, когда один знакомый систематик, бледный от ужаса, сообщил мне “известие”, что такой-то (имя значения не имеет) “переметнулся к кладистам”.
Нижеследующий краткий обзор таксономических школ, вероятно, раздосадует кое-кого из их представителей, но уж точно не сильнее, чем они сами обычно раздражают друг друга, так что никакого дополнительного вреда я не причиню. Если говорить о самых основах их философии, то все систематики делятся на два больших лагеря. С одной стороны располагаются те, кто без всякого стеснения откровенно заявляет, что их цель заключается в выявлении эволюционного родства. Для них (как и для меня) хорошая система живых организмов — это и есть генеалогическое древо. Занятие систематикой состоит в том, чтобы, используя все доступные методы, сделать как можно более правдивые предположения относительно взаимного родства тех или иных животных. Этой группе систематиков трудно подобрать название, потому что самое очевидное — “эволюционные систематики” — уже узурпировано одним более узким направлением. Иногда их называют приверженцами “филетики”. Таким образом, пока что я писал эту главу, стоя на филетических позициях.
Однако многие систематики подходят к делу иначе, имея на то довольно веские основания. Хотя они, вероятно, и согласны с тем, что конечной целью классификации является выявление эволюционного родства, они настаивают на том, что таксономическая практика должна быть отделена от теории — по всей видимости, от эволюционной теории, — объясняющей, откуда взялись общие признаки у тех или иных организмов. Они изучают сходство как таковое. Они не делают исходных предположений о том, является ли оно следствием эволюционной истории и связано ли близкое сходство с близким родством, и предпочитают строить свою систему, основываясь исключительно на совокупности общих признаков.
Одно из преимуществ такого подхода состоит в том, что если у вас есть какие-то сомнения насчет существования эволюции, то вы можете использовать имеющуюся совокупность сходств и различий, чтобы проверить свои предположения. Если эволюция действительно существует, тогда сходства и различия должны образовывать некий предсказуемый паттерн, и прежде всего паттерн иерархической вложенности. Если же эволюция — выдумка, тогда бог его знает, какого паттерна следует ожидать, однако никаких явных причин предполагать иерархическую вложенность у нас не будет. Представители этого направления настаивают на том, что если вы занимаетесь систематикой исходя из существования эволюции, то вы не вправе использовать результаты своей работы для доказательства того, что эволюция существует, — ваша аргументация окажется порочной. Такие рассуждения имели бы ценность, если бы в существовании эволюции кто-нибудь всерьез сомневался. Для этого второго направления систематики — так же как и для первого — трудно подобрать подходящее название. Я буду называть его приверженцев “измерителями чистого сходства”.
Сторонники филетического подхода — то есть те систематики, которые откровенно занимаются выявлением эволюционного родства, — сами в свою очередь подразделяются на два течения. Это кладисты, которые следуют принципам, изложенным в знаменитой книге Вилли Хеннига “Филогенетическая ситематика”, и “традиционные” эволюционные систематики. Кладисты одержимы родовыми ветвями. По их мнению, цель таксономии состоит в том, чтобы выяснить, в каком порядке организмы ответв лялись друг от друга в ходе эволюционной истории, и их не интересует, насколько большие или насколько малые изменения претерпели эти ветви с того момента, как разошлись. А “традиционные” (не следует воспринимать этот эпитет как негативный) эволюционные систематики отличаются от кладистов главным образом тем, что помимо самого факта расхождения родовых ветвей также учитывают суммарное количество произошедших с этими ветвями изменений.
Кладисты, как только приступают к работе, сразу же начинают мыслить категориями ветвящихся деревьев. Теоретически в первую очередь они должны нарисовать все возможные варианты ветвящихся генеалогических деревьев для рассматриваемых ими животных (допускаются только разветвления надвое, и не более, ибо любому терпению есть предел!). Когда речь шла о молекулярной таксономии, мы уже видели, что если животных много, то сделать это не так-то просто, поскольку число возможных родословных астрономически велико. Но также мы видели и то, что, на наше счастье, существуют вполне работающие способы упростить задачу, и на практике такого рода систематика вполне возможна.
Если мы, просто для наглядности, попытаемся систематизировать всего-навсего трех животных: кальмара, сельдь и человека, — то возможны только следующие три дихотомически ветвящихся генеалогических древа:

Кальмар и сельдь являются друг для друга более близкими родственниками, чем для человека.
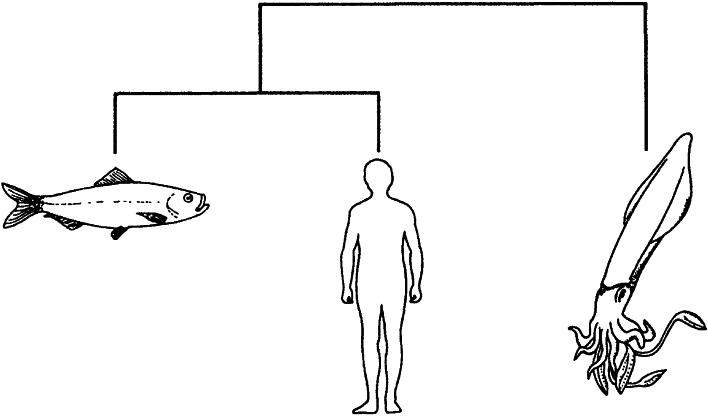
Человек и сельдь являются друг для друга более близкими родственниками, чем для кальмара.
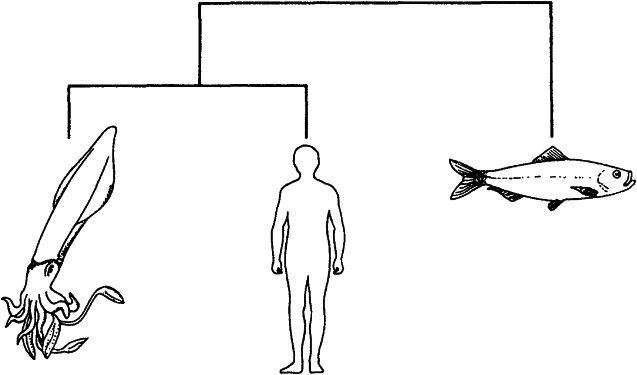
Кальмар и человек являются друг для друга более близкими родственниками, чем для сельди.
Кладист должен рассмотреть каждое из этих деревьев и выбрать наилучший вариант. Как его распознать? В принципе лучшим долж но являться то дерево, которое объединяет вместе животных, обладающих наибольшим количеством общих черт. Организм, имеющий наименьшее количество общих черт с остальными двумя, выносится “за группу”. Из трех представленных выше родословных предпочтение следует отдать второй, поскольку у человека с сельдью намного больше общих признаков, чем у любого из этих организмов с кальмаром. Кальмар вне группы, потому что у него мало общих черт как с человеком, так и с сельдью.
На самом деле все это несколько сложнее, чем простой подсчет общих признаков, поскольку некоторые признаки осознанно игнорируются. Особую важность кладисты придают тем признакам, что возникли недавно. К примеру, по древним отличительным особенностям, унаследованным всеми млекопитающими от их общего предка, невозможно составить классификацию млекопитающих. Методы, используемые для различения древних и новых признаков, интересны, но их описание увело бы нас слишком далеко от тематики настоящей книги. На данном этапе рассуждений нам достаточно лишь уяснить, что кладист — по крайней мере в принципе — рассматривает все возможные бифуркационные деревья, которые могли бы объединить классифицируемых им животных, и старается выбрать из этих деревьев единственное верное. А еще настоящий кладист нисколько не стесняется того, что считает эти ветвящиеся схемы, или “кладограммы”, отражением генеалогии, эволюционного родства.
Одержимость ветвящимися деревьями, доведенная до крайности, может приводить к неожиданным результатам. Теоретически вид может оказаться в мельчайших деталях идентичен своим дальним родственникам и при этом коренным образом отличаться от более близкой родни. В качестве примера давайте представим себе два очень похожих друг на друга вида рыб — условно назовем их Иаков и Исав, — которые жили 300 млн лет назад. Каждый из этих двух видов дал начало династии потомков, существующей и по сей день. Потомки Исава остановились в развитии. Они продолжали вести глубоководный образ жизни и не эволюционировали. Поэтому современный потомок Исава в сущности ничем не отличается от самого Исава, а следовательно, очень похож и на Иакова. Потомки же Иакова эволюционировали и множились. В конце концов они дали начало всем современным млекопитающим. Однако одна из ветвей этой обширной династии тоже осталась жить в морских глубинах и остановилась в своем развитии. Нынешние представители этой ветви являются рыбами, настолько похожими на современных потомков Исава, что их сложно отличить друг от друга.
Ну и как же прикажете классифицировать этих животных? Традиционный эволюционный систематик отдал бы должное огромному сходству между примитивными глубоководными потомками Иакова и Исава и объединил бы их в одну группу. Но строгий кладист не имеет права так делать. Глубоководные потомки Иакова, как бы ни были они похожи на глубоководных потомков Исава, приходятся тем не менее более близкими родственниками млекопитающим. Их с млекопитающими общий предок жил позже, пусть даже и самую чуточку позже, их общего предка с потомками Исава, и потому следует объединить их в одну группу с млекопитающими. Кому-то такой подход может показаться чудачеством, но лично я воспринимаю его без раздражения. По крайней мере, он абсолютно ясен и логичен. Свои преимущества в самом деле есть и у кладистики, и у традиционной эволюционной систематики, и для меня не слишком важно, как именно люди классифицируют животных, до тех пор пока они в состоянии связно объяснить мне принципы своей классификации.
Вернемся теперь к другому важнейшему направлению систематической науки — к “измерителям чистого сходства”. Их, в свою очередь, тоже можно разделить на две школы. Обе они сходятся в том, что нужно изгонять эволюцию из повседневных занятий систематикой. Однако по поводу того, в чем должны заключаться эти повседневные занятия, между ними согласия нет. Одно из этих поднаправлений иногда называют “фенетикой”, а иногда — “нумерической таксономией”. Я же буду называть их “измерителями средних расстояний”. А представители другой школы измерителей сходства сами себя называют “трансформированными кладистами”. Название это крайне неудачно, так как на самом деле они являются всем чем угодно, но только не кладистами. Когда Джулиан Хаксли придумал термин “клада”, он дал ему ясное и недвусмысленное определение в понятиях эволюционного ветвления и эволюционного родства. Клада — это совокупность всех организмов, являющихся потомками некоего общего предка. А поскольку главная идея “трансформированных кладистов” состоит в том, чтобы избегать каких бы то ни было упоминаний об эволюции и о предках, то называть себя кладистами не слишком разумно с их стороны. Причина того, что они все же так себя называют, коренится в истории: они начинали как настоящие кладисты и продолжают использовать некоторые методы кладистики, отказавшись от лежащих в ее основе философии и логики. Боюсь, что у меня нет выбора и придется, хоть и с крайней неохотой, называть их трансформированными кладистами.
Мало того что “измерители средних расстояний” отказываются использовать эволюционную теорию в своей систематике (хотя в существовании эволюции они и не сомневаются) — они призывают даже не исходить из того допущения, что паттерн имеющихся сходств между организмами непременно должен иметь форму простой ветвящейся иерархии. Они стараются применять такие методы, которые должны обнаружить иерархическую структуру, если она действительно существует, и не обнаружить ее, если ее нет. Они спрашивают природу, в самом ли деле она организована по иерархическому принципу. Задача не из простых, и возможно, справедливо будет признать, что в действительности методов для ее решения у нас нет. Тем не менее сама эта цель представляется мне разновидностью похвального стремления избегать предвзятости. Методы, которыми пользуется данная школа, зачастую насыщены изощренной математикой и в равной степени пригодны для классификации как живых организмов, так и неживых объектов, например минералов или археологических экспонатов.
Эти систематики начинают обычно с того, что измеряют у рассматриваемых ими животных все показатели, какие только возможно. Интерпретировать полученные таким образом данные — задачка для смекалистых, но вдаваться в подробности я не буду. В конечном итоге из совокупности всех измерений выводится коэффициент сходства (или обратная ему величина — коэффициент различия) между какими-либо двумя организмами. При желании вы можете представить себе животных в виде точек, образующих скопления в пространстве. Крысы, мыши, хомяки и т. д. будут собраны поблизости друг от друга. А вдалеке от них, в другой части этого пространства, можно будет увидеть еще одно скопление, состоящее из львов, тигров, леопардов, гепардов и им подобных. Расстояние между любыми двумя точками в таком пространстве является мерилом того, насколько два данных животных похожи друг на друга, если судить на основании большого числа признаков. Расстояние между львом и тигром небольшое, так же как и между крысой и мышью. Однако расстояние между крысой и тигром, мышью и львом велико. Сопоставление различных признаков осуществляется обычно с помощью компьютера. Это пространство, в котором находятся животные, отдаленно и поверхностно напоминает Страну биоморф, однако разница в том, что здесь “расстояние” между организмами отражает их собственное сходство, а не сходство их генов.
После того как среднее сходство (или расстояние) между каждым животным и каждым другим животным рассчитано, компьютер переходит к следующему пункту своей программы: тщательно перебрать весь имеющийся набор расстояний/сходств и попытаться втиснуть его в иерархическую схему разделения на группы. К сожалению, по поводу того, какой именно вычислительный метод использовать для выявления этих групп, имеется немало разногласий. Такого метода, который со всей очевидностью можно было бы назвать единственным верным, не существует, а разные методы не всегда дают один и тот же результат. Хуже того, не исключено, что некоторые из этих методов с чрезмерным “усердием” пытаются “увидеть” иерархическую организацию даже там, где ее нет. В последнее время школа измерителей расстояния, или “нумерических таксономистов”, несколько вышла из моды. Я считаю, что это временное явление, как оно нередко бывает с модой, и что подобную “нумерическую таксономию” не так-то просто списать со счетов. Полагаю, она еще возьмет реванш.
К другой школе измерителей чистого сходства относятся те, кто называет себя — как уже говорилось, по историческим причинам — трансформированными кладистами. Именно эта группа является основным источником тех самых “оскорблений”. Я не буду, как обычно делают, выискивать ее корни в рядах настоящих кладистов. Если говорить о философии, лежащей в основе их деятельности, то так называемые трансформированные кладисты имеют больше общего с другими измерителями чистого сходства — с теми, кто занимается “фенетикой” или “нумерической таксономией”, которую мы только что обсудили. Всех их объединяет неприязнь к тому, чтобы приплетать эволюцию к таксономической практике, хотя это и не обязательно свидетельствует о враждебности по отношению к идее эволюции как таковой.
Общими для трансформированных и для настоящих кладистов являются многие используемые ими практические методы. И те и другие изначально мыслят категориями ветвящихся деревьев. И те и другие подразделяют признаки на таксономически важные и на бесполезные для систематики. Однако логическое обоснование у такого подразделения в каждом случае свое. Подобно измерителям средних расстояний трансформированные кладисты отнюдь не стремятся к выяснению родословных. Свои деревья они выстраивают не на основании генеалогии, а руководствуясь исключительно сходствами и различиями. Они согласны с измерителями средних расстояний в том, что вопросом, является ли имеющееся распределение сходных признаков следствием эволюционной истории, задаваться не следует. Но если измерители расстояний готовы, хотя бы теоретически, узнать у природы, организована она по иерархическому принципу или нет, то трансформированные кладисты исходят из того, что организована. Это для них аксиома, символ веры: все на свете должно быть систематизировано в виде иерархического ветвления (или, что то же самое, в виде вложенных друг в друга ячеек). А поскольку их ветвящиеся деревья не имеют никакого отношения к эволюции, то не обязательно строить их только для живых организмов. Методы трансформированной кладистики подходят, если верить их сторонникам, для классификации не только животных и растений, но также камней, планет, библиотечных книг и горшков бронзового века. Иными словами, трансформированные кладисты не согласились бы с моей проводившейся выше мыслью, что эволюция — это единственное разумное основание для однозначной иерархической системы.
Как мы уже видели, измерители средних расстояний оценивают, насколько далеко располагается одно животное от другого: большая дистанция означает для них отсутствие сходства, а маленькая — его наличие. И только когда этот своеобразный суммарный показатель сходства уже вычислен, они пытаются представить свои результаты в виде ветвящейся “древовидной” диаграммы, где одни группы входят в состав других. Однако трансформированные кладисты, подобно настоящим кладистам, к которым они некогда принадлежали, сразу же начинают оперировать группами и бифуркациями. Их цель, как и цель настоящих кладистов, состоит — по крайней мере теоретически — в том, чтобы построить все возможные древовидные схемы и выбрать из них самую лучшую.
Но о чем они на самом деле говорят, когда рассматривают каждое возможное “дерево”, и что они имеют в виду под наилучшим? Какому гипотетическому состоянию мира соответствует каждая из диаграмм? Для истинного кладиста, последователя В. Хеннига, ответ на этот вопрос кристально ясен. Каждая из 15 альтернативных схем, которые могут объединить между собой четырех животных, отображает предполагаемую родословную. Из всех этих 15 мыслимых родословных одна и только одна должна быть верной. Предки данных четырех животных действительно существовали в реальности. При допущении, что генеалогическое древо способно ветвиться только дихотомически, мы имеем 15 возможных вариантов истории. Четырнадцать из них наверняка ошибочны. Только один является правильным, то есть соответствующим той истории, которая произошла на самом деле. Из всех 135 135 родословных, какие можно составить для восьми животных, 135 134 должны быть неправильными и только одна — соответствующей исторической правде. Определить, какая именно верна, бывает непросто, но настоящий кладист имеет основания быть уверенным хотя бы в том, что существует не более одного правильного варианта.
Но чему соответствуют 15 (или 135 135, или сколько бы ни было) возможных диаграмм, одна из которых верная, в неэволюционирующем мире трансформированного кладиста? Ничему особенному, как разъяснил мой коллега и бывший студент Марк Ридли в своей книге “Эволюция и классификация”. Трансформированный кладист запрещает себе пользоваться понятием происхождение. “Предок” для него — неприличное слово. Но при этом он настаивает, что его классификация должна обладать таким свойством, как иерархическое ветвление. Но если 15 (или 135 135) иерархически ветвящихся деревьев не представляют собой различные варианты родословной, то чем же, скажите на милость, они являются? Остается только позаимствовать у античной философии невнятные идеалистические представления о том, что мир просто так иерархически устроен и что все на свете имеет свою “противоположность”, свой мистический инь или ян. До чего-то более конкретного дело никогда не доходит. В неэволюционирующем мире трансформированного кладиста абсолютно невозможно делать четкие и ясные утверждения наподобие следующего: “Из 945 диаграмм, которые можно составить для шести животных, верной может быть только одна; все остальные непременно ошибочны”.
Почему слово “предок” является, с точки зрения кладистов, ругательным? Дело (надеюсь) не в том, что они думают, будто никаких предков никогда не существовало. Скорее, это следствие той установки, что предкам нет места в систематике. Пока речь идет о повседневной таксономической практике, такая позиция вполне оправданна. Кладисты никогда не помещают на свои диаграммы предков наряду с реальными животными, хотя традиционные эволюционные систематики иногда и поступают подобным образом. Кладисты всех мастей занимаются строго взаимосвязями между реально существующими, наблюдаемыми в природе животными, предпочитая говорить о родстве, а не о происхождении. И это чрезвычайно разумно. Неразумно только перегибать палку и превращать в табу само понятие “предок”, запрещая говорить о происхождении даже в тех случаях, когда речь идет о фундаментальном обосновании того, почему мы беремся представлять классификацию живых организмов в виде схемы с иерархическим ветвлением.
Самую необычную особенность трансформированной кладистики я приберег напоследок. Некоторым представителям этой школы показалось недостаточным вполне аргументированное убеждение, что эволюционным спекуляциям не место в практических занятиях систематикой, — убеждение, которое они разделяют с занимающимися фенетикой “измерителями расстояний”, — и они хватили через край, придя к заключению, что ошибочна сама эволюция. В такую эксцентричность практически невозможно поверить, но тем не менее некоторые из ведущих “трансформированных кладистов” выказывают открытую враждебность по отношению к самой идее эволюции, а главным образом — к дарвиновской эволюционной теории. Два сотрудника Американского музея естественной истории, Г. Нельсон и Н. Платник, дошли до того, что написали следующее: “Дарвинизм… если вкратце, — это теория, которая была проверена и оказалась ложной”. Я был бы счастлив узнать, в чем заключалась эта “проверка”, а еще любопытнее мне то, с помощью какой альтернативной теории Нельсон и Платник собираются объяснять явления, объясняемые дарвинизмом, в частности сложные адаптации.
Это не означает, что среди трансформированных кладистов есть убежденные креационисты. Как я сам понимаю, им просто доставляет удовольствие преувеличивать значимость систематики для биологии. Они решили — возможно, не без оснований, — что будут заниматься систематикой с бóльшим успехом, если забудут об эволюции, а главное — если не будут использовать понятие “предок” в своих рассуждениях о классификации. Точно так же, скажем, тот, кто изучает нервную систему, может прийти к выводу, что размышления об эволюции ничем ему не помогут. Нейробиолог не возражает против того, что нервные клетки возникли в результате эволюции, но ему не нужно использовать этот факт в своей исследовательской работе. Ему необходимо хорошо знать физику и химию, а вот дарвинизм, по его мнению, не имеет отношения к повседневному изучению нервных импульсов. Такая позиция имеет право на существование. Но нельзя, будучи в здравом уме, утверждать, что если некая теория не используется вами в ваших каждодневных узкоспециальных занятиях, то, следовательно, эта теория ложна. Сказать такое может только тот, кто считает свою научную отрасль исключительной, несопоставимой с остальными по важности.
Но даже в таком случае это нелогично. Физикам, чтобы заниматься физикой, дарвинизм определенно не нужен. Физик может считать биологию ничтожной дисциплиной по сравнению с физикой. Тогда, по его мнению, и научная ценность дарвинизма может быть невелика. Но это не дает ему никаких разумных оснований для того, чтобы считать дарвинизм ложным. Однако именно так, судя по всему, поступили некоторые из крупнейших представителей школы трансформированной кладистики. Обратите внимание: Нельсон и Платник использовали именно этот эпитет — “ложный”. Надо ли говорить, что их слова были подхвачены чувствительными микрофонами, упоминавшимися в предыдущей главе, и получили большую огласку, а их имена заняли почетное место на страницах религиозных и креационистских изданий. Недавно к нам в университет приезжал с лекцией один из ведущих трансформированных кладистов, и ему удалось собрать больше народу, чем любому другому приглашенному лектору, выступавшему в тот год. Нетрудно догадаться почему.
Нет никаких сомнений в том, что такие замечания, как “дарвинизм… это теория, которая была проверена и оказалась ложной”, исходя из уст авторитетных биологов, состоящих в штате почтенного государственного музея, будут лучшим подарком для креационистов и вообще для всех тех, кто активно заинтересован в том, чтобы распространять неправду. Только по этой причине я взялся надоедать своим читателям с трансформированной кладистикой. Как более сдержанно выразился Марк Ридли в своей рецензии на ту самую книгу, в которой Нельсон и Платник назвали дарвинизм ложным, “кто бы мог догадаться, что на самом деле они имели в виду всего-навсего то, что предковым формам затруднительно найти место в кладистической классификации?” Разумеется, определить точное систематическое положение эволюционных предков нелегко, и есть убедительные аргументы в пользу того, чтобы даже не пытаться этого делать. Но позволять себе высказывания, которые могут навести на мысль, будто никаких предковых форм вообще не было, значит злоупотреблять словами и изменять истине.
Пойду-ка я лучше возделывать свой сад.
Назад: Глава 9 Пререкания о прерывистом
Дальше: Глава 11 Обреченные противники

