Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Надзор за надзирателями: кто присматривает за регуляцией развития
Дальше: Заключение
Hox-гены обрели свободу — и змеи потеряли ноги
Напоследок рассмотрим исследование, проливающее свет на роль Hox-генов в эволюции позвоночных. Как известно, важнейшая функция Hox-генов состоит в том, что они подробно размечают эмбрион вдоль передне-задней оси. Дальнейшая судьба эмбриональных клеток, оказавшихся в той или иной части эмбриона, зависит от набора Hox-генов, экспрессирующихся в этой части. Для каждого Hox-гена характерна своя область экспрессии. Например, гены Hox12и Hox13, как правило, работают только в задней части эмбриона, которая в дальнейшем станет хвостом; гены Hox10у некоторых позвоночных работают от заднего конца эмбриона до той черты, которая станет границей между грудным отделом (где на позвонках есть ребра) и поясничным, где ребра не развиваются. « Hox-код», определяющий план строения организма, сложен и не совсем одинаков у разных групп позвоночных. Вряд ли можно сомневаться в том, что многие крупные эволюционные преобразования, затрагивающие план строения, были связаны с изменениями в структуре и экспрессии Hox-генов. Однако хорошо изученных примеров, иллюстрирующих эту связь, пока немного .
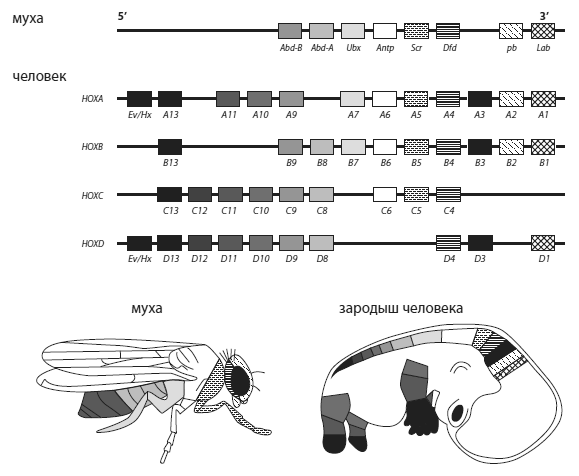 Hox -гены дрозофилы и человека.Прямоугольниками обозначены гены в том порядке, в каком они расположены в хромосомах. У мухи один наборHox -генов, у человека — четыре, частично дублирующие друг друга (они образовались из одного в результате двух полногеномных дупликаций). Кластеры A, B, C, D находятся на разных хромососмах (у мыши это хромосомы № 6, 11, 15 и 2, у человека — № у, 17, 2, 12). У змей, в отличие от мыши и человека, в кластере D отсутствует 12-й ген (Hoxd12 ). На изображениях мухи и зародыша человека области экспрессии соответствующих генов окрашены теми же цветами, что и сами гены. По последним данным, соответствие междуHox -генами членистоногих и позвоночных несколько менее однозначно, чем показано на этой схеме.
Hox -гены дрозофилы и человека.Прямоугольниками обозначены гены в том порядке, в каком они расположены в хромосомах. У мухи один наборHox -генов, у человека — четыре, частично дублирующие друг друга (они образовались из одного в результате двух полногеномных дупликаций). Кластеры A, B, C, D находятся на разных хромососмах (у мыши это хромосомы № 6, 11, 15 и 2, у человека — № у, 17, 2, 12). У змей, в отличие от мыши и человека, в кластере D отсутствует 12-й ген (Hoxd12 ). На изображениях мухи и зародыша человека области экспрессии соответствующих генов окрашены теми же цветами, что и сами гены. По последним данным, соответствие междуHox -генами членистоногих и позвоночных несколько менее однозначно, чем показано на этой схеме.
У многих животных, в том числе у позвоночных, Hox-гены в геноме располагаются кластерами, т. е. группами вплотную друг другу. Самое удивительное, что порядок расположения генов в Hox-кластерах часто (хотя и не всегда) совпадает с распределением областей экспрессии вдоль передне-задней оси: впереди находятся «головные» гены, за ними следуют гены, отвечающие за формирование средних участков тела, а замыкают кластер «задние» гены, управляющие развитием задних частей туловища. По-видимому, это связано со способом регуляции экспрессии Hox-генов: участок ДНК, где находится Hox-кластер, постепенно «раскрывается», становясь доступным для транскрипции по мере движения от переднего конца тела к заднему. Поэтому у переднего конца тела экспрессируются только передние Hox-гены, а чем ближе к хвосту, тем более задние гены включаются в работу. Удобный способ регуляции генов, отвечающих за разметку эмбриона вдоль передне-задней оси!
У предков позвоночных, как у современного ланцетника, в геноме был один Hox-кластер, включающий 14 генов. На ранних этапах эволюции позвоночных произошло две полногеномные дупликации. В результате позвоночные приобрели четыре Hox-кластера вместо одного. Это открыло перед позвоночными большие эволюционные возможности (см. главу 5). Отдельные Hox-гены в некоторых кластерах были утрачены, но в целом их набор и порядок расположения остался сходным во всех четырех кластерах. Паралогичные гены (т. е. копии одного и того же Hox-гена в разных Hox-кластерах) приобрели немного различающиеся функции, что дало возможность тонко регулировать эмбриональное развитие и облегчило развитие новых планов строения.
Биологи из Швейцарии, Новой Зеландии и США изучили работу Hox-генов у чешуйчатых рептилий (отряд Squamata) ( Di-Poi et al., 2010). Этот отряд, объединяющий ящериц и змей, интересен разнообразием планов строения и вариабельностью признаков, связанных с передне-задней дифференцировкой туловища (относительная длина отделов тела, число позвонков в них и т. п.) Поэтому логично было предположить, что Hox-кластеры чешуйчатых должны обладать специфическими особенностями и что Hox-гены ящериц и змей должны различаться.
Ранее было показано, что области экспрессии передних Hox-генов у змей расширились в заднем направлении по сравнению с другими позвоночными. Это хорошо согласуется с общим удлинением тела. Кроме того, было установлено, что правило колинеарности (т. е. одинаковый порядок расположения генов в кластере и областей их экспрессии в эмбрионе) у змей строго соблюдается.
Исследователи сосредоточились на задних Hox-генах (от 10-го до 13-го). Главными объектами исследования были хлыстохвостая ящерица Aspidoscelis uniparensи маисовый полоз Elaphe guttata. Кроме того, были отсеквенированы Hox-кластеры нескольких других ящериц, гаттерии и черепахи. Для сравнения использовались Hox-кластеры курицы, человека, мыши и лягушки.
Набор задних Hox-генов у всех исследованных видов оказался одинаковым, если не считать того, что у змей и лягушек «потерялся» ген Hoхd12(12-й Hox-ген из кластера D). Важные изменения были обнаружены в регуляторных участках Hox-кластеров. Оказалось, что все чешуйчатые рептилии утратили регуляторный участок между генами Hoхd13и Evх2 , а змеи вдобавок потеряли консервативный некодирующий элемент между Hoхd12и Hoхd13и некоторые регуляторные участки в других Hox-кластерах. Неожиданным результатом оказалось присутствие в Hox-кластерах чешуйчатых множества встроившихся мобильных генетических элементов. В результате общая длина задней части Hox-кластеров у чешуйчатых значительно выросла по сравнению с другими наземными позвоночными.
Все это, по-видимому, говорит о том, что у чешуйчатых ослабли эволюционные ограничения, препятствующие накоплению изменений в задней части Hox-кластеров. Очищающий отбор, отбраковывающий подобные изменения у других позвоночных, в эволюции ящериц и змей действовал менее эффективно. Этот вывод подтвердился и в ходе анализа кодирующих участков Hox-генов. В этих участках у ящериц, и особенно у змей, по сравнению с другими позвоночными накопилось много значимых замен. Одни из них, по-видимому, зафиксировались случайно, из-за ослабления очищающего отбора, тогда как другие закрепились под действием положительного отбора, т. е. были полезными.
Изучение характера экспрессии задних Hox-генов у эмбрионов ящерицы и полоза подтвердило предположение о том, что изменения плана строения в эволюции чешуйчатых были тесно связаны с изменениями в работе задних Hox-генов.
У ящерицы, как и у других наземных позвоночных, передний край области экспрессии генов Hoxa10и Hoxc10в точности соответствует границе между грудным и поясничным отделами. Одной из функций этих генов является подавление развития ребер. У змей нет поясничного отдела, а на бывших крестцовых позвонках (у змей они называются клоакальными) имеются особые раздвоенные ребра. По-видимому, эти особенности связаны с тем, что Hox-гены у предков змей утратили способность останавливать рост ребер.
Область экспрессии Hoxa10и Hoxc10у полоза заходит далеко в грудной отдел. Эти гены отвечают также за своевременное прекращение роста грудного отдела. По-видимому, эта их функция у змей тоже ослаблена, что могло быть одной из причин удлинения грудного отдела у змей по сравнению с их предками — ящерицами. Удлинение хвостового отдела у змей связано с тем, что из четырех генов, «тормозящих» рост хвоста у ящериц ( Hoxa13, Hoxc13, Hoxd13, Hoxd12) один ген у змей полностью утрачен ( Hoxd12), а два других ( Hoxa13, Hoxd13) не участвуют в передне-задней «разметке» эмбриона и используются только в формировании половых органов.
Многочисленные случаи независимой утраты и частичной редукции конечностей у чешуйчатых тоже могут быть связаны с тем, что в этом отряде задние Hox-гены получили нетипичную для других животных эволюционную «свободу». На них стал слабее действовать очищающий отбор, что позволило быстро накапливать мутации.
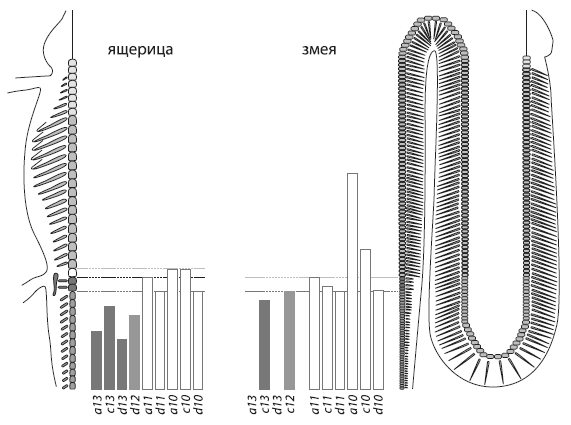 Области экспрессии заднихHox -генов у ящерицы и змеи. У ящерицы перед хвостовыми позвонками расположены два крестцовых(показаны темно-серым цветом) , затем следует один рудиментарный поясничный позвонок(белый) , а дальше идут грудные позвонки(серые) . У змеи нет поясничного отдела, а вместо крестцовых имеются четыре клоакальных позвонка с раздвоенными ребрами(темно-серые) .Вертикальными прямоугольниками показаны области экспрессии заднихHox -генов. ИзDi-Poi et al., 2010 .
Области экспрессии заднихHox -генов у ящерицы и змеи. У ящерицы перед хвостовыми позвонками расположены два крестцовых(показаны темно-серым цветом) , затем следует один рудиментарный поясничный позвонок(белый) , а дальше идут грудные позвонки(серые) . У змеи нет поясничного отдела, а вместо крестцовых имеются четыре клоакальных позвонка с раздвоенными ребрами(темно-серые) .Вертикальными прямоугольниками показаны области экспрессии заднихHox -генов. ИзDi-Poi et al., 2010 .
Известно, что задние Hox-гены играют ключевую роль не только в оформлении задних отделов туловища, но и в развитии конечностей. Поэтому некоторые мутации этих генов, ведущие, например, к удлинению тела или к редукции поясничного отдела, теоретически могут приводить и к таким побочным эффектам, как редукция конечностей. Удлинение тела в сочетании с редукцией конечностей встречается и в других группах позвоночных (например, у некоторых амфибий). Было ли это связано с такими же изменениями в работе Hox-генов, как у змей, или с другими, покажут дальнейшие исследования.
Эволюционная биология развития — быстро развивающаяся дисциплина, от которой следует ожидать важнейших научных прорывов. Расшифровка генно-регуляторных сетей, управляющих развитием, — одна из самых насущных задач биологии. Ее решение позволит понять не только соотношение между генотипом и фенотипом, но и важнейшие правила и закономерности эволюции сложных организмов. Когда эти правила, известные нам сегодня лишь в общих чертах, будут изучены досконально, вплоть до построения строгих математических моделей, перед человечеством откроются небывалые возможности. Проектирование «с чистого листа» биологических систем с нужными нам свойствами — лишь одна из них. Другая — совершенствование нашей собственной природы. Все это будет. Нужно лишь четко уяснить, для каких целей это нужно будущему человечеству, и надеяться, что культурное, социальное и морально-этическое развитие человечества к тому времени исключит возможность использования этих открытий во вред.

