Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Эволюционный стартап у бабочек-подражателей
Дальше: Глава 7 Переходные формы
Эволюция, повернувшая вспять
До какого-то момента процесс видообразования остается обратимым. Изменение условий может привести к тому, что наметившиеся различия сгладятся, а начавшие расходиться популяции снова объединятся. Это может произойти даже с «хорошими» видами, по поводу которых у специалистов нет сомнений, виды это или все-таки разновидности. В наши дни основной причиной таких «обратных эволюционных ходов» является антропогенное воздействие на среду. Например, по словам ихтиолога М. В. Мины из Института биологии развития РАН, под угрозой слияния оказались многие виды цихлид озера Виктория ( Мина, 2001). Причина — в неразумном решении вселить в озеро нильского окуня, крупного хищника, который быстро размножился в озере и поставил многие местные виды под угрозу физического уничтожения. Что касается видов, еще не съеденных окунем, то некоторые из них начали скрещиваться друг с другом, «растворяя» свои уникальные особенности в смешанных генофондах. Дело в том, что нильский окунь — рыба крупная. Его нельзя просто высушить на солнышке, как испокон веков поступали местные рыбаки с мелкой рыбешкой (теми самыми цихлидами). Окуня нужно коптить, а для этого нужны дрова. Вырубка деревьев по берегам озера усилила эрозию почв. Дожди стали смывать в озеро больше глины и песка. Озерная вода, и без того мутноватая, замутилась еще больше. В мутной воде многие цихлиды перестали различать нюансы окраски потенциальных партнеров, служившие изолирующим барьером между видами. Началась гибридизация.
Пассивное накопление генетических различий по модели Добжанского — Мёллера у цихлид требует обычно от 4 до 14 млн лет для развития полной несовместимости. Возраст озера Виктория намного меньше, поэтому большинство местных цихлид еще сохранили способность к гибридизации. Таким образом, вселение одного-единственного вида поставило под угрозу «великий эволюционный эксперимент» природы.
Еще один пример обратного хода эволюции исследовали экологи из Швейцарии и Канады. Ученые воспользовалась превосходными данными по сигам из швейцарских горных озер. В их распоряжении имелись профессионально собранные данные 1940-х годов, а также современные материалы, отражающие разнообразие сигов в начале XXI века ( Vonlanthen et al., 2012). В науке так всегда бывает: чем надежнее данные, тем интереснее результат. Не стало исключением и это исследование. Обычно при анализе эволюции оценивают степень расхождения видов, здесь же наблюдали обратное. Ученые зарегистрировали эволюцию, повернувшую вспять.
Использовались данные по 17 озерам в предгорьях Швейцарских Альп. Это глубокие водоемы с хорошо аэрированной водой, изначально олиготрофные, т. е. с малым содержанием органики. В озерах издавна обитали сиги (род Coregonus). Они заселились после освобождения этой территории от ледника (10–12 тыс. лет назад) и начали осваивать новое экологическое пространство. В ходе специализации и разделения ниш в каждом озере образовались «букеты» симпатрических видов (1–5 в каждом озере), в том числе характерные пары экологических аналогов. В каждой такой паре один вид предпочитает мелководья, питается мелкой добычей, нерестует в зимние месяцы; обычно это крупные рыбы с небольшим числом тычинок на жаберных дугах. Второй вид — его контрагент; обычно это мелкая рыба с большим числом жаберных тычинок, предпочитает жить на глубине, а нереститься летом, питается более крупным зоопланктоном. Число жаберных тычинок отражает пищевую специализацию, четко наследуется и считается хорошим маркером видовой принадлежности. Учитывая разночтения систематики рода Coregonus, в озерах насчитывается всего около 25 видов и подвидов сигов.
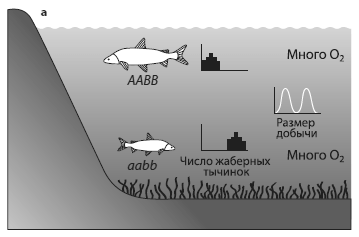
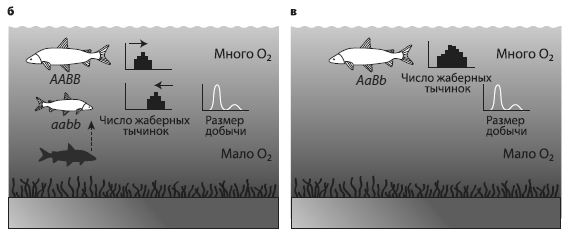 Уменьшение разнообразия сигов в озерах предгорьев Альп. а — ситуация в начале xx века: существует два вида — крупный и мелкий. Крупный обитает в неглубоких водах, мелкий — где поглубже, число жаберных тычинок у крупного меньше (маленькие гистограммы напротив рыбок) в соответствии с размером предпочитаемой добычи (маленькие графики). Кислорода в воде достаточно. б — ситуация после эвтрофикации, когда в придонных слоях воды мало кислорода. Число жаберных тычинок и размер предпочтительной добычи у мелких рыб уменьшается. в — исчезновение глубоководного вида. У мелководного при этом увеличивается генетический полиморфизм и вариабельность по числу жаберных тычинок и размеру добычи. ИзMcKinnon, Taylor, 2012 .
Уменьшение разнообразия сигов в озерах предгорьев Альп. а — ситуация в начале xx века: существует два вида — крупный и мелкий. Крупный обитает в неглубоких водах, мелкий — где поглубже, число жаберных тычинок у крупного меньше (маленькие гистограммы напротив рыбок) в соответствии с размером предпочитаемой добычи (маленькие графики). Кислорода в воде достаточно. б — ситуация после эвтрофикации, когда в придонных слоях воды мало кислорода. Число жаберных тычинок и размер предпочтительной добычи у мелких рыб уменьшается. в — исчезновение глубоководного вида. У мелководного при этом увеличивается генетический полиморфизм и вариабельность по числу жаберных тычинок и размеру добычи. ИзMcKinnon, Taylor, 2012 .
Таково было состояние озерных систем до 1950-х годов, описанное в классических трудах европейских ихтиологов. Каковы же современные данные? А они таковы, что около четверти видов сигов исчезло из видового реестра, причем вымерли в основном мелкие глубоководные виды. Связано это с загрязнением озер и резким повышением содержания органики в воде. Механизм вымирания на первый взгляд прост. Увеличение количества органики вызывает бурный рост микрофлоры, которая окисляет органику растворенным в воде кислородом. В результате содержание кислорода, особенно в придонных слоях, падает, нерестилища глубоководных рыб оказываются непригодными, молодь гибнет и вид исчезает.
Схема как будто проста, но, если поразмышлять о деталях процесса, все окажется интереснее. Вид быстро исчезает… Совсем исчезает? Ведь исчезнуть можно по-разному. В данном случае глубоководный вид слилсяс мелководным. Два вида начали скрещиваться, и в результате уцелевший мелководный вид приобрел некоторые черты исчезнувшего глубоководного. Это было доказано с помощью анализа морфологической (по числу жаберных тычинок) и генетической вариабельности.
В одном случае процесс обратного слияния видов удалось «поймать за хвост», увидеть его в действии. Этот пример исследовали на материале Цюрихского и Валенштадтского озер. Пример хорош тем, что к истории видов прилагается в качестве бонуса еще и история самих озер. Два озера когда-то были одним целым. В это послеледниковое праозеро заселился один предковый вид. Там же он разделился на два эндемичных вида — мелководный C. duplexи глубоководный C. heglingus. Затем в праозере обособились две части. Оба вида оказались в обоих частях озера — теперь в двух отдельных озерах, и генетический анализ действительно доказывает такое их происхождение. Однако генетическая удаленность видов друг от друга оказалась разной в двух озерах. В Цюрихском озере, сильно эвтрофицированном, они генетически ближе друг к другу и явно скрещиваются, а в чистом Валенштадском озере их генетическая вариабельность не перекрывается. Это значит, что в Цюрихском озере вовсю идет процесс стирания границ между видами: глубоководный вид сливается с мелководным.
Не менее интересен пример с Боденским озером. Там еще осталось три вида сигов, в том числе глубоководный и мелководный. Но если у глубоководного вида разнообразие аллелей осталось прежним по сравнению с историческими образцами, то у мелководного помимо своих «родных» аллелей обнаружены почти все аллели глубоководного вида. Мелководный вид постепенно ассимилирует в своем генофонде аллели глубоководного собрата. Так что его вымирание становится неочевидным с философских позиций — вид исчезнет, но гены его останутся.
Все эти озера образовались при таянии ледников около 10 тыс. лет назад. Вскоре эволюция развела специализированные формы по разным экологическим нишам. На этом этапе симпатрического видообразования сформировалась презиготическая изоляция: формы не скрещивались в силу различий в сроках и местах нереста. Этого оказалось достаточно, чтобы появились морфологически различимые виды. Но, как только неблагоприятная обстановка подорвала ресурсы одной из ниш — глубоководной, виды начали обратное схождение. Десяток тысяч лет естественной эволюции перечеркнут полувековым антропогенным вмешательством. Соотношение скоростей естественной и антропогенной эволюции поражает и настраивает на невеселые размышления. Как быстро можно разрушить долгую и кропотливую работу природы!
Но эмоции не должны умалить двух замечательных выводов швейцарско-канадского исследовательского коллектива: эволюция до какой-то степени обратима, и вымирание видов не всегда означает их совершенное исчезновение. Эволюция оказалась податливым процессом, а вид, точнее его генетические рудименты, может продолжить существование в генофонде близкого вида. И опять мы возвращаемся к началу этой главы — к рассуждению о сущности биологического вида, о зыбкости и неопределенности формального определения, хотя именно вид, дискретный и стабильный, составляет основу биологического разнообразия природы.

