Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Насекомые — для цветов, цветы — для насекомых
Дальше: Видообразование — личное дело каждого
Как отличить своих от чужих?
Смена кормового растения у насекомых, хозяина у паразитов, смена опылителя или сдвиг сроков цветения у растений — примеры простых и удобных способов быстрого развития презиготической изоляции. Есть и другие пути ускоренного видообразования. Например, многие виды растений возникли в результате отдаленной гибридизации с последующей полиплоидизацией гибридов. Гибридные растения, появившиеся в результате скрещивания давно разошедшихся видов, обычно бесплодны, потому что хромосомные наборы их родителей сильно различаются. Из-за этого хромосомы не могут найти себе пару во время мейоза, их расхождение по дочерним клеткам нарушается, и эти клетки (микро- и мегаспоры) оказываются нежизнеспособными. Гибрид, однако, может размножаться вегетативно. А если у него произойдет полногеномная дупликация (полиплоидизация) — удвоение всего химерного генома, — то способность к нормальному мейозу может восстановиться. В результате получится полноценный новый вид, представители которого могут скрещиваться друг с другом, но не с родительскими видами. Полиплоидизация может создать новый репродуктивно изолированный вид и без предварительной гибридизации. Возможно, 2–4 % видов цветковых и 7 % папоротников возникли таким путем ( Otto, Whitton, 2000).
Но подобные способы все же производят впечатление неких частностей, которые не могут объяснить все случаи симпатрического видообразования. Возможно, существует какой-то более общий механизм быстрого формирования презиготической изоляции?
На такую возможность указывают результаты опытов по «искусственному видообразованию». Главный вывод, который можно сделать из этих опытов, состоит в том, что интенсивный разнонаправленный отбор иногда способствует развитию не только постзиготической изоляции (как в эксперименте с дрожжами), но и презиготической. Формируется положительная ассортативность скрещиваний, или эндогамия: брачные предпочтения смещаются в сторону выбора своих и избегания чужих.
В одном из ранних экспериментов мух D. melanogasterподвергали дизруптивному отбору(отбору «на расхождение», когда репродуктивное преимущество имеют особи с крайними значениями признака, а особи с промежуточными значениями отбраковываются). Мух с максимальным и минимальным числом щетинок сохраняли, с промежуточным — отсеивали. В результате между расходящимися частями популяции (много- и малощетинковыми мухами) возникла частичная презиготическая изоляция ( Thoday, Gibson, 1962).
Такие же результаты получены при выращивании D. melanogasterв контрастных условиях («холодно-сухо-темно» и «тепло-влажно-светло»). Здесь тоже изоляция (презиготическая и отчасти постзиготическая) возникла между популяциями, приспособившимися к разным условиям, тогда как между популяциями, независимо друг от друга приспособившимися к одинаковым условиям, изоляция не возникла ( Kilias et al., 1980).
Явления, обнаруженные в этих экспериментах, наблюдаются и в природе: например, у дрозофил, обитающих на разных склонах так называемого каньона Эволюции на горе Кармел (Израиль). Популяции D. melanogaster, живущие на разных склонах каньона на расстоянии всего 100–400 м друг от друга (в нескольких минутах полета!), проявляют статистически достоверные предпочтения при скрещивании, избегая спариваний с обитателями другого склона. Условия на склонах сильно различаются, поскольку склон, обращенный к югу, получает в шесть раз больше солнечного излучения ( Korol et al., 2000).
Еще один эксперимент был проведен на замечательных мухах Cyrtodiopsis dalmanni. У самцов этого вида глаза расположены на длинных стебельках (у самок — на коротких). Признак поддерживается половым отбором: самцы, конкурирующие за самку, меряются глазными стебельками. Тот, у кого они оказались короче, улетает пристыженный. Что касается самок, то они однозначно предпочитают длинноглазых кавалеров.
В эксперименте производился искусственный дизруптивный отбор: в одних линиях отбирались самцы с максимальным расстоянием между глазами (т. е. с самыми длинными глазными стебельками), в других — с минимальным. Удалось вывести линии, сильно различающиеся по этому признаку. Затем были исследованы половые предпочтения самок в разных линиях. Оказалось, что в контрольных (не подвергавшихся отбору) и длинностебельковых линиях самки по-прежнему предпочитают самцов с наибольшим расстоянием между глазами. Однако в короткостебельковых линиях предпочтения самок изменились на противоположные: они стали предпочитать самцов с самыми короткими глазными стебельками. Из-за изменившихся женских предпочтений между линиями возникла частичная презиготическая изоляция ( Wilkinson, Reillo, 1994).
Почему в «короткоглазых» линиях изменились брачные предпочтения самок? Ведь отбор шел не по брачным предпочтениям, а по длине глазных стебельков, да и подвергались отбору только самцы. Вопрос этот очень важен, а ответ на него очень прост. Если отбор поддерживает какой-то признак у самцов, то он при этом автоматически и неизбежно поддерживает также и склонность самок предпочитать самцов с данным признаком. В самом деле, короткоглазый самец — это, скорее всего, сын короткоглазого самца и самки, которая его выбрала. Длинноглазый самец, соответственно, сын длинноглазого отца и матери, которая любила длинноглазых кавалеров. Поэтому, когда ученые отсеивали длинноглазых самцов, они одновременно отбраковывали и «гены длинных глаз», и «гены любви к длинноглазым самцам». Скорее всего, в исходной популяции существовала изменчивость по женским предпочтениям. В линиях, отбиравшихся на короткоглазость, получили преимущество самки-«извращенки», предпочитавшие короткоглазых самцов вопреки «общему мнению».
Это рассуждение применимо к любым вариантам и способам выбора полового партнера. Всегда, когда отбор поддерживает какой-то признак, он автоматически поддерживает и склонность выбирать особей с этим признаком в качестве партнеров. Возможно, это один из универсальных механизмов формирования эндогамии (предпочтения «своих») под действием отбора. Правда, не всегда понятно, как выбирающая особь может определить, имеется ли у претендента нужный признак. Если речь идет о длине рогов или глазных стебельков, тут все ясно, эти признаки и так находятся под действием полового отбора, они на виду, и самки миллионы лет отбирались на способность их различать. Ну а если отбор идет на способность переваривать какой-нибудь полисахарид или, скажем, на устойчивость к холоду? Как в этом случае отличить своего от чужого?
—————Брачного партнера выбирают по микрофлореОдин из самых ярких экспериментов по «искусственному видообразованию» провела в конце 1980-х годов Диана Додд из Йельского университета ( Dodd, 1989). Она взяла восемь линий мух Drosophila pseudoobscura, происходящих от одной природной популяции, и поместила их в стрессовые условия: четыре линии были посажены на диету из мальтозы, четыре — на крахмал. Затем были проведены тесты на избирательность при спаривании. Оказалось, что у мух всего за один год (не более 2–3 десятков поколений) сформировалась сильная положительная ассортативность (предпочтение своих) при выборе партнера: мальтозные мухи предпочитали спариваться с мальтозными, крахмальные — с крахмальными. Как и почему сформировались такие предпочтения, было неясно, ведь мух отбирали не на привередливость в отношении брачного партнера и все восемь лабораторных линий жили в изоляции друг от друга. Избирательность сформировалась автоматически, как побочное следствие адаптации к разным средам.
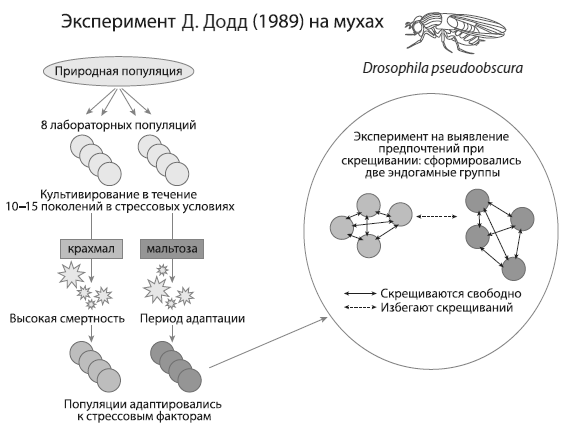 Схема эксперимента Додд (1989). МухиDrosophila pseudoobscura , в течение нескольких поколений привыкавшие к жизни на двух нестандартных средах (крахмале и мальтозе), в дальнейшем предпочитали скрещиваться только со «своими», т. е. с мухами, приспособившимися к такому же корму. Из:Куликов, Марков, 2009 .
Схема эксперимента Додд (1989). МухиDrosophila pseudoobscura , в течение нескольких поколений привыкавшие к жизни на двух нестандартных средах (крахмале и мальтозе), в дальнейшем предпочитали скрещиваться только со «своими», т. е. с мухами, приспособившимися к такому же корму. Из:Куликов, Марков, 2009 .
Этот и другие подобные эксперименты поставили перед исследователями два основных вопроса.
1. Почемупосле адаптации к жизни в контрастных условиях насекомые начинают предпочитать «своих» в качестве брачных партнеров? Может быть, это адаптация, позволяющая избежать «размывания» полезных комплексов аллелей, благодаря которым родители сумели выжить в стрессовых условиях?2. Как, по каким признакамони отличают своих от чужих? Нужно заметить, что в таких экспериментах последнее поколение перед тестами на избирательность выращивают на одинаковой стандартной среде. Это важный момент. Из-за этой предосторожности мухи не могут различить друг друга ни по запаху корма (например, крахмала или мальтозы), ни по состоянию здоровья, которое сигнализировало бы о плохой или хорошей приспособленности, если бы мухи развивались на своих средах. Но мухи все равно различают своих и чужих — по каким-то изменениям, которые произошли с ними в ходе адаптации к новым условиям.
Естественно предположить, что речь идет о генетических изменениях, произошедших под действием отбора. Это могло быть, например, изменение частот каких-то аллелей, снижение генетического полиморфизма и рост гомозиготности. Однако новые эксперименты, проведенные учеными из Израиля и США, показали, что дело в данном случае не в генах самих мух, а в изменениях микрофлоры — комплекса микроорганизмов, живущих на теле и в кишечнике насекомых ( Sharon et al., 2010).Авторы повторили эксперимент Додд с небольшими изменениями. Они работали не с D. pseudoobscura, а с более изученным видом D. melanogaster. Исходную лабораторную линию разделили на две: одну выращивали на стандартном корме, основанном на кукурузной муке и патоке, другую — на крахмале, как в опытах Додд. По прошествии определенного числа поколений часть мух из каждой линии пересаживали на стандартную среду и ждали еще одно поколение, а затем проводили тесты на сексуальную привередливость.Сначала авторы проверили, каких брачных партнеров будут выбирать мухи после 11 поколений жизни на контрастных средах. Результат получился такой же, как у Додд: мухи продемонстрировали четкую положительную ассортативность. Из 38 спариваний в 29 случаях образовались пары своих со своими (гомогамные спаривания), и только девять раз «крахмальные» мухи спарились с «паточными» (гетерогамные спаривания).После этого эксперимент был повторен, но теперь избирательность проверяли в каждом поколении. Результаты оказались неожиданными. Положительная ассортативность не развивалась постепенно по мере адаптации мух к разным условиям. Уже после первого поколения наблюдалось предпочтение своих, и в дальнейшем оно оставалось таким же, не увеличиваясь, но и не снижаясь вплоть до 37-го поколения, когда эксперимент был прекращен. Ранее никто не наблюдал появления таких предпочтений всего за одно поколение.Авторы провели контрольный эксперимент: две популяции мух выращивали в течение 27 поколений изолированно, но на одинаковой (паточной) среде. В этом случае ассортативность не возникла: мухи одинаково охотно спаривались как со своими, так и с чужими. Значит, дело не в изолированном развитии популяций, а именно в разных условиях жизни. Такой же результат был получен и Дианой Додд: ее мальтозные мухи не делали различий между партнерами из своей линии и тремя другими мальтозными линиями; то же наблюдалось и в четырех крахмальных линиях.Затем авторы еще раз повторили исходный эксперимент, помещая последнее поколение мух перед тестированием в пробирки со средой, содержащей антибиотик. Оказалось, что эта процедура ведет к полному исчезновению избирательности. Теперь мухи спаривались с кем ни попадя, не делая различий между своими и чужими.Этот результат уже позволял предположить, что мухи различают своих и чужих по микрофлоре, однако его можно было интерпретировать и иначе (может быть, сами антибиотики каким-то образом делают мух неразборчивыми). В следующем эксперименте мух, обработанных антибиотиком, повторно «заражали» той или иной микрофлорой. Из крахмальной и паточной сред, в которых жили мухи, выделили бактерий и заразили ими пробирки со стерильной паточной средой. В пробирки посадили «вылеченных» антибиотиками мух, и уже через одно поколение у них восстановилась положительная ассортативность. Мухи опять стали предпочитать своих, т. е. живших на среде с такой же микрофлорой (хотя сама среда на этот раз была одинаковая).Микрофлора крахмальных и паточных мух оказалась разнообразной. Главное различие в том, что в микрофлоре крахмальных мух примерно в десять раз больше бактерий рода Lactobacillus. Особенно бурно расплодились в крахмальных мухах микробы Lactobacillus plantarum(которые в небольшом количестве есть и у мух из исходной популяции). Эти бактерии живут в кишечнике мух, где они вырабатывают фермент амилазу, тем самым помогая хозяевам переваривать крахмал. Именно эти микробы и оказывают самое сильное влияние на выбор мухами брачных партнеров.По-видимому, микрофлора влияет на количество и состав углеводородных феромонов, выделяемых мухами. Впрочем, не исключено, что мухи могут непосредственно детектировать присутствие бактерий или продуктов их жизнедеятельности на теле партнера. Известно, что млекопитающие определяют зараженность сородичей теми или иными бактериями при помощи специальных «иммунологических» рецепторов . Может быть, и у насекомых имеется такая возможность.Итак, выбор брачного партнера у мух зависит не только от их собственного генома, но и от симбиотических бактерий. Учитывая, что алгоритмы выбора брачного партнера могут оказывать самое радикальное влияние на ход эволюции, приходится признать, что в такой ситуации уже трудно рассматривать эволюцию мух отдельно от эволюции их бактериальных симбионтов.Изменения среды могут приводить к быстрым изменениям состава микрофлоры у насекомых. У насекомых ассортативность вырабатывается за одно поколение, но мы должны учитывать, что у бактерий это десятки поколений. Так что времени для образования приспособленного к конкретной среде микробного сообщества предостаточно. Положительная ассортативность, в свою очередь, способствует быстрому формированию частичной презиготической изоляции, что может стать первым шагом на пути к разделению исходного вида на два.Теперь мы знаем, что именно меняется у мух при выращивании на контрастных средах и по какому признаку они отличают своих от чужих. Правда, по-прежнему неясно, почемуони предпочитают скрещиваться с партнерами, обладающими сходной микрофлорой, и каковы молекулярные и нейрологические механизмы различения особей с разными наборами симбионтов.—————

