Книга: Мутанты. О генетической изменчивости и человеческом теле.
Назад: Ближе – дальше
Дальше: Кисти, стопы и предки
Стремление к десятичности
Метрическая система, имеющая в своей основе десять чисел, существует лишь потому, что у ученых мужей из Французской академии, разработавших эту систему, было на руках по десять пальцев, на которых они, предположительно, учились считать. Если бы математикой занялись свиньи, они, наверное, измеряли бы свои помои в единицах Международной системы, которая основывалась бы на восьми числах, так как у них имеется по четыре пальца на каждом копыте. У лошадей приходится один палец на конечность, У верблюдов – два, у слонов – пять, у морских свинок, однако, имеется по четыре пальца на передних конечностях и по три – на задних. У кошек и собак по пять пальцев на передних и задних ногах, но один из них очень маленький, рудиментарный. За исключением некоторых разновидностей лягушек и дельфинов, называемых вакита, у большинства позвоночных редко встречается на одной конечности больше пяти пальцев.
Почему так происходит, абсолютно неясно. Вовсе не потому, что лишние пальцы невозможно создать. У млекопитающих всех видов иногда встречаются дополнительные пальцы, но как исключение, а не правило. Сенбернары, пиренейские овчарки, ньюфаундленды и другие крупные собаки особенно подвержены появлению дополнительных пальцев на каждой ноге, причем обычно у них удваивается тот самый рудиментарный палец. У Эрнеста Хемингуэя все кошки были с полидактилией, и их многопалые потомки до сих пор живут на территории его дома в Ки-Уэсте. Пятнадцать процентов уличных кошек в Бостоне характеризуются полидактилией (у некоторых насчитывается до десятка лишних пальцев), а вот в Нью-Йорке среди бродячих кошек вы таких не встретите. Существует немало линий мышей с полидактилией: одну из них назвали "сасквоч" в честь снежного человека, у остальных более прозаичные наименования, вроде "двоеногих" или "лишнепалых". Американский генетик Сьюэлл Райт однажды вывел морскую свинку с сорока четырьмя пальцами на передних и задних конечностях, но долго она не протянула.
У многих людей также имеются лишние пальцы. Примерно 1 на 3000 европеоидов рождается с дополнительными пальцами на руках или ногах (либо и на тех и на других); у африканцев это соотношение составляет 1:300. В принципе может удваиваться любой палец, но у африканцев им обычно бывает мизинец, а у европейцев – большой палец. Полидактилия – это, как правило, генетическая аномалия, часто передающаяся по доминантному типу и встречающаяся в некоторых семьях во многих поколениях. Задолго до появления Грегора Менделя французский математик Пьер-Луи Моро де Мопертюи (1698-1758) описал наследование полидактилии у предков и потомков берлинского врача по имени Якоб Руэ. У его бабушки было по шесть пальцев на каждой руке и ноге, равно как у его матери, у него самого, у троих из его семи братьев и сестер и у двух из его пяти детей. Другие ученые приводили примеры еще более внушительных родословных с полидактилией. Так, в 1931 году русский генетик Е.О. Манойлов опубликовал сообщение об одном грузине с полидактилией, Вячеславе Михайловиче де Камио-Сципионе, который, по его словам, мог проследить свое происхождение от многопалых предков на протяжении шести поколений.
Если апикальный эктодермальный гребень обеспечивает рост наших конечностей в пространстве, за пределами тела, то другая, в равной мере незаметная часть почки конечностей следит за тем, чтобы у нас было нужное число и правильный набор пальцев. Это открытие опять-таки было сделано Джоном Сондерсом вместе с его сотрудницей Мэри Гасселинг. Они обнаружили, что если пересадить кусочек мезодермы с хвостового конца одной из куриных почек конечностей на головной конец другой (так, чтобы почка имела два хвостовых конца с противоположной ориентацией по отношению друг к другу), то в результате у курицы появится крыло с двойным набором обычных пальцевых структур. Самое замечательное в этом было то, что полученные в результате эксперимента крылья были похожи на одну, особенно редкостную разновидность полидактилии, встречающуюся у людей. Они напоминали конечности людей, которые не просто характеризовались добавочными пальцами, а практически обладали полным дубликатом пальцев руки или ноги, так что общее их число на одной конечности могло доходить до десяти. Крылья с полидактилией имели своеобразное зеркальное расположение пальцевых структур, точно такое же, как и на продублированных кистях у людей. Если пронумеровать каждый палец, так что большой палец будет по порядку 1-м, указательный – 2-м, средний – 3-м, безымянный – 4-м и мизинец – 5-м, то формула для нормальной пятипалой кисти будет "12345", а для руки с двойным набором пальцев: "5432112345". Это совершенно удивительное явление, своеобразный анатомический палиндром.
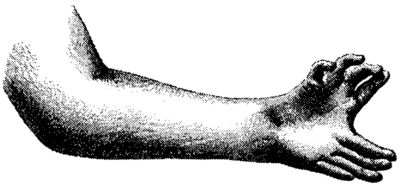
Зеркальная полидактилия.Из книги Уильяма Бейтсона "Материалы для изучения изменчивости", 1894 (Империал-колледж, Лондон).
Сондерс и Гасселинг назвали открытый ими кусочек мезодермы, обладающий столь могучими свойствами, зоной поляризующей активности, или ЗПА (ZPA). Считают, что это источник морфогена. В самом центре, где концентрация его наиболее велика, этот морфоген заставляет необученную мезодерму становиться мизинцем; чуть поодаль более низкие концентрации индуцируют безымянный, средний и указательный пальцы, поочередно, а на противоположном конце конечности будет сформирован большой палец.
Этот отчет о том, как большинство из нас получает свои пять пальцев, наводит на мысль о существовании организатора. Подобно ему, ЗПА обладает необыкновенным талантом призывать к порядку свое окружение. И, как в течение многих лет продолжались поиски морфогена организатора, так в последние годы ищут морфоген ЗПА. Почти наверняка это будет сигнальный белок, вероятно – какой-нибудь из уже известных, член большого семейства сигнальных белков, повсеместно функционирующих в эмбрионе. Но почки конечностей содержат множество таких белков, и пока трудно установить, который из них является собственно морфогеном. За последние годы появилось несколько кандидатов на эту роль, полностью отвечающих всем необходимым требованиям. Один из них – это соник-хеджхог.
Соник-хеджхог появляется в почке конечности там и только там, где и должно находиться морфогену: в мезодерме хвостовой части, что в точности совпадает с положением ЗПА Сондерса и Гасселинг. Он также занимается тем, что и должен делать морфоген: формированием конечностей. Крыльям цыпленка можно придать новые, невероятные формы, в том числе и такие, которые удваивают нормальный набор пальцев и дают его зеркальное отражение, – все это за счет изменения концентрации соника в почке. И наконец, существуют мутанты. Мутации по меньшей мере в десяти генах приводят к полидактилии у людей, и все они как будто бы влияют тем или иным способом на роль соника в конечности.
Но, как мы уже говорили в предыдущей главе, соник-хеджхог не просто определяет, сколько именно пальцев у нас вырастет на руках и ногах. Он также разделяет наш мозг, решает, как широко будут расставлены наши глаза, и регулирует много других важных деталей. Это безнадежно неразборчивая молекула. Если бы можно было пронаблюдать за активностью гена соник-хеджхога в течение некоторого времени, как при замедленной фотосъемке, мы бы увидели, что его деятельность охватывает весь развивающийся организм зародыша или плода и что он включается или выключается то в одном формирующемся органе, то в другом.
Перед механизмами, ответственными за эти процессы, стоит грандиозная по сложности задача, и они практически нигде, учитывая могущество соника в определении судьбы клеток, не имеют права на ошибку. Эти механизмы – транскрипционные факторы, или "молекулярные переключатели". Некоторые из них осуществляют контроль за соником. Если мутации выводят их из строя, соник начинает включаться в тех частях почки конечностей, где обычно не должен этого делать, в результате чего у нас появляются лишние пальцы на руках и ногах. Другие мутации не выводят из строя транскрипционные факторы, а скорее уничтожают регуляторные элементы, с которыми они связываются. Результат, однако, будет тем же самым: смешение градиентов морфогена и ошибки в формировании пальцев.
Мутации, приводящие к полидактилии, ослабляют контроль со стороны соник-хеджхога, изменяя баланс сил в пользу его повсеместного распространения. Другие мутации обладают прямо противоположным эффектом и вообще не дают сонику появляться в зачатке конечности. Самый яркий пример такой мутации – это выведение из строя самого гена соника. Мыши, у которых отсутствует соник, в добавление к другим многочисленным дефектам, вообще не имеют лап. Это удивительно напоминает нарушение, о котором уже говорилось выше: ахейроподию, или ту аномалию, которой страдают алейжадиньос. Действительно, есть доказательства, хотя и не бесспорные, что приводящая к ахейроподии мутация выводит из строя регуляторный элемент, необходимый для присутствия соника в конечности.
Это перечисление мутаций дает лишь очень отдаленные представления об истинной сложности процессов генной регуляции у зародыша. Например, факт включения или невключения гена в данной клетке зависит от того, какие транскрипционные факторы содержит ее ядро, а их присутствие, в свою очередь, зависит от еще одних транскрипционных факторов и так далее. На первый взгляд, подобного рода иерархии как будто вовлекают нас в бесконечную цепь таких зависимостей, когда задача по организации порядка попросту перекладывается на вышестоящие структуры, которые, в свой черед, тоже подчиняются чьим-то приказам. Но дилемма эта скорее кажущаяся, нежели реальная. Формирование эмбриона – не однократный процесс. Конкретное присутствие соника в ЗПА определяется отчасти активностью Hox-генов в мезодерме туловища, из которой вырастают конечности. Но приказы относительно пространственного расположения частей, отдаваемые этими генами, довольно расплывчаты; в задачи соника входит их дальнейшее уточнение. Помимо соника, конечно же, существуют и другие детализирующие уровни, которые осуществляют упорядочивание структуры в более мелких масштабах, причем каждый из этих уровней должен вести постоянные, подробные переговоры, природа которых нам пока что не вполне понятна.
Эта картина последовательных уровней переговоров и контроля может показаться невообразимо сложной. Но на деле она недостаточно сложна, так как ей не удается покорить одну из наиболее общих характеристик программы развития зародыша: его нелинейность. Выше я утверждал, что приводящая к ахейроподии мутация препятствует появлению соника в конечности. И тем не менее я начал эту главу с предположения, что внутриутробные ампутации конечностей разной степени тяжести, проявляющиеся у новорожденных, вызваны нарушениями в апикальном эктодермальном гребне и продуцируемых им фибробластных факторах роста. В этом можно было бы усмотреть противоречие, но оно возникает только в том случае, если мы думаем, что сигналы различных конечностей не зависят друг от друга, тогда как в действительности это не так. Ибо одна из наиболее важных ролей соник-хеджхога состоит в том, чтобы поддерживать и формировать апикальный эктодермальный гребень с его фибробластными факторами роста, а одна из самых важных ролей апикального эктодермального гребня – поддерживать и контролировать продукцию соник-хеджхога в зоне поляризирующей активности. Этот двусторонний обмен информацией не менее рискован, чем перекидка предметами между двумя жонглерами, стоящими на разных концах сцены. Подобное взаимовлияние повсюду присутствует в развивающемся эмбрионе и в корне меняет наши взгляды на процессы роста и развития. Мы начинаем с представлений о линейных путях приказов и контроля и о простом плане строения, а потом пытаемся распутать узел. И когда, как в случае с конечностями, мы действительно начинаем видеть очертания программы развития, она однозначно напоминает запутанный клубок сетей, совершающих головокружительные круговороты во времени и пространстве. Сети в данном случае обеспечивают одно простое правило: если мы начинаем пересчитывать пальцы на руках и ногах, то обычно заканчиваем числом двадцать.
Назад: Ближе – дальше
Дальше: Кисти, стопы и предки

