Грибы разных лесов
Как формируется видовой состав грибов, соотношение экологических групп, от чего зависит этот процесс? Попробуем ответить на эти вопросы. Молодняки (ельники, сосняки, березняки) в возрасте до 10—15 лет представляют собой такие растительные сообщества, в которых конкуренция за элементы минерального питания, влагу, свет выражена максимально. Практически все деревья здесь находятся в неблагоприятных условиях обитания, за исключением тех, которые растут на опушках. Экологические условия внутри таких массивов относительно однородны. Колебания факторов местообитания по сравнению с лесами более старого возраста незначительны, об этом свидетельствуют данные о суточной динамике освещенности, температуры воздуха и почвы на различных уровнях в 15- и 60-летних ельниках, представленные на рис. 4. В связи с этим относительная влажность воздуха и влажность почвы в молодняках более или менее стабильны в течение суток и вегетационного периода в целом. Соответственно особенностям экологических факторов формируются видовой состав и экологическая структура макромицетов. Последняя чрезвычайно проста и составлена обычно двумя группами — микоризообразователями и подстилочными сапротрофами, приуроченными в своем развитии к слаборазложившейся хвое, листьям, мелким ветвям. Распределение плодовых тел симбиотрофов характеризуется большой скученностью (пятнистостью) и тяготением к границам массивов. Площади пятен не велики (до 10—15 м2), но на каждое плодовое тело в них приходится не более 0,25 м2. Часто наблюдается явление «убегания» плодовых тел симбиотрофов из-под полога насаждений на расстояние до 5 м и более. Чрезвычайно беден и видовой состав микоризообразующих грибов в молодняках, обычно это один, два вида: в березняках — подберезовик, в ельниках — белый, в сосняках — масленок. Деревья, растущие на границах молодняков, гораздо лучше обеспечены элементами почвенного минерального питания, влагой, светом, чем те, которые находятся внутри массивов. С увеличением возраста последние чаще подвергаются ветровалу, снеголому, что в конечном итоге приводит к их выпадению (гибели) и образованию окон в древесном пологе. Это незамедлительно вызывает перестройку структуры лесных сообществ, вносит разнообразие в условия обитания для грибов. В связи с увеличением запасов подстилки усложняется структура напочвенных сапротрофов, а на выпавших из древостоя породах и больных деревьях появляются ксилотрофы. На место погибших деревьев внедряются другие породы и, естественно, грибы, связанные с ними. Усложнение структуры макромицетов ведет к увеличению их видового разнообразия, но численность представителей всех групп при этом резко падает. Групповое распределение плодовых тел, характерное для молодняков, сменяется рассеянным.
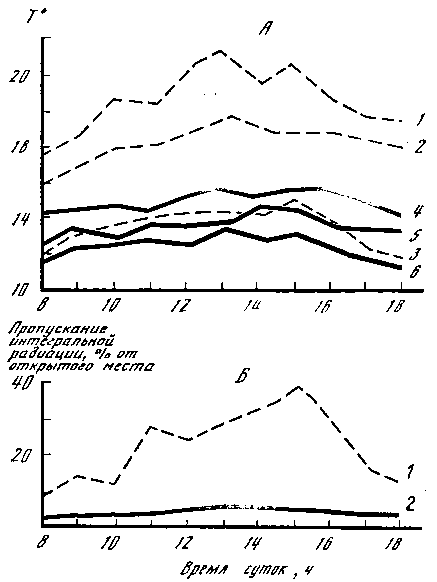
Рис. 4. Варьирование температуры и освещенности в 15- и 60-летних ельниках
А — температура в 60-летнем (1 — на высоте 1,5 м; 2 — на поверхности почвы; 3 — на глубине 5 см) и 15-летнем ельнике (4 — на высоте 1,5 м; 5 — на поверхности почвы; 6 — на глубине 2—3 см). Б — пропускание интегральной радиации (в % от открытого места): 1 — в 60-летнем; 2 — в 15-летнем ельнике
Максимальное количество видов грибов наблюдается в средневозрастных (60—90-летних) лесах: ельниках, дубравах, березняках, сосняках. Это можно проследить на примере ельников: в 10-летних общее количество видов макромицетов — 6, в 20-летних — 16, в 30-летних — 27, в 60-летних — 90. Дальнейшее увеличение возраста лесов приводит к уравниванию экологических условий под пологом леса, уменьшению видового разнообразия и численности грибов до определенного уровня, который остается более или менее стабильным продолжительное время.
Мы рассмотрели в общих чертах закономерности формирования структуры группировок макромицетов в различных по возрасту и составу лесах умеренного климата.
Существенную поправку в эту модель вносят почвенногрунтовые условия, в которых находятся леса. Для древесных пород, занимающих более или менее сходные по обеспеченности элементами минерального питания и влажностью местообитания, эти поправки не велики, в противном случае они достигают огромных величин. Так, например, из найденных в четырех типах сосняков Прикамья 170 видов макромицетов общими для них оказались всего 8, а для такого же числа типов ельников из 180 видов выявлено 83 общих.
В формировании видового состава макромицетов играет основную роль не только (и даже не столько) состав древостоя, сколько условия, в которых находятся лесные сообщества. В географически удаленных (Подмосковье и Эстония) ельниках, находящихся в одинаковых почвенно-грунтовых условиях, видовое сходство грибов составляет 80%.
Рассмотрим влияние условий обитания, в которых может произрастать одна и та же лесообразующая порода, на формирование видового состава, численность и структуру микоризообразующих грибов, энергетически непосредственно связанных с деревьями. Как уже отмечалось, деревья в молодняках находятся в крайне неблагоприятных условиях обитания. Низкая освещенность, острая конкуренция за элементы минерального питания и влагу препятствуют процессу микоризообразования, поэтому симбиотрофы в них характеризуются бедным видовым составом, групповым распределением плодовых тел и большой численностью. Эти особенности микоризообразующих грибов характерны не только для молодняков, но и для лесов более старого возраста, находящихся в чрезвычайно неблагоприятных условиях местообитания. К таковым можно отнести березняки, растущие на скалах Кольского полуострова, березовые полезащитные полосы полупустынь Прикаспия, сильновытоптанные березняки Подмосковья. Показатели видового сходства грибов для таких лесов гораздо выше, чем для лесов, образованных той же породой, но находящихся в более благоприятных почвенно-грунтовых условиях. Переход к более оптимальным условиям произрастания для деревьев сопровождается возрастанием количества видов симбиотрофов, уменьшением обилия плодовых тел каждого вида, отсутствием группового распределения плодовых тел, быстрой сменой во времени грибных слоев.
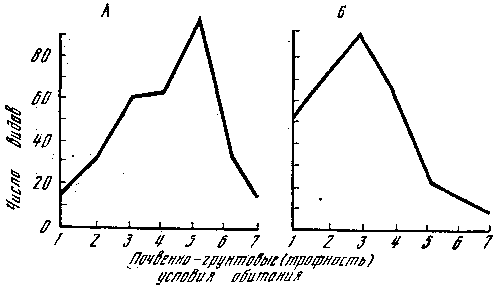
Рис. 5. Распределение симбиотрофов в сосняках и ельниках различного типа
А — сосняки; 1 — лишайниковый; 2 — вересковый; 3 — скальный; 4 — бруснично-зеленомошный; 5 — чернично-зеленомошный; 6 — кисличный; 7 — долгомошный и сфагновый. Б — ельники: 1 — брусничный; 2 — скальный; 3 — черничный; 4 — кисличный; 5 — черничный влажный; 6 — долгомошный; 7 — хвощово-сфагновый
Для лесов Карелии (ельники, сосняки) одинаково низкое количество симбиотрофов отмечено как в условиях недостаточной (сосняк лишайниковый, скальный), так и избыточной (сосняк долгомошный и сфагновый, ельник долгомошный, хвощово-сфагновый) почвенной влагообеспеченности, с которыми связана и обеспеченность растений минеральным питанием (рис. 5). Видовое сходство гораздо выше, например, между ельником и сосняком брусничным (0,74), чем между сосняками лишайниковым и сфагновым (0,37), т. е. в лесах, сложенных одной и той же породой, но находящихся в резко различных условиях обитания. Показательны в этом отношении сосняки, поскольку они занимают самые различные условия обитания и интразональны в своем распространении, но приурочены в основном к бедным песчаным почвам и потому особо нуждаются в симбиотрофных грибах. Обычно с ними связаны 50—70 видов микоризообразующих макромицетов, но в условиях произрастания на богатых меловых почвах их число снижается до 11. В искусственных разновозрастных сосняках, посаженных на богатых суглинистых почвах Подмосковья, встречаются только два вида, верных сосне, — поздний и зернистый масленок, и те в небольшом количестве и не ежегодно, по-видимому, в таких благоприятных условиях (почвенно-грунтовых) сосна не нуждается в грибном симбионте.
Сосняки на песках обычно образуют светлые редкостойные леса. Хорошая освещенность в сосновых лесах стимулирует отток углеводов к корням деревьев, и интенсивность микоризообразования в данном случае находится в прямой зависимости от концентрации в почве азота, фосфора и калия, их связанности почвенным поглощающим комплексом (доступности для растений) и обеспеченности почв кислородом. Известно, что в ряде почв, особенно при недостатке и избытке влаги, фосфор и калий, непосредственно участвующие в превращении энергии и входящие в состав основных продуктов жизнедеятельности деревьев, прочно связываются почвенным поглощающим комплексом и тем самым исключаются из биологического круговорота. Известно также, что периоду всасывания питательных веществ микоризными корнями предшествует процесс обмена веществ, зависящий в первую очередь от температуры и обеспеченности почв кислородом. Поглощенные корнями деревьев доступные соединения элементов питания поступают в гифы грибов. Фосфор переходит в мицелии в связанное состояние и впоследствии транспортируется в ткани дерева. После отмирания микоризных корней фосфор возвращается в почву. Наибольшее варьирование подвижного фосфора (Р2O5) наблюдается в самых верхних горизонтах почвы — зоне максимальной концентрации тонких корней. Другие доступные для растений элементы минерального питания (в частности, обменные основания), как и фосфор, обнаруживают четкую аналогию с распределением по вертикальному профилю почв гифов микоризообразующих грибов. Например, на сухих почвах под лишайниковым бором гифы достигают глубины 25 см, а в сосняках брусничных, занимающих обычно склоны песчаных холмов (транзитные формы рельефа), а именно верхние их части, где резко выражена скорость транзита минеральных и органических веществ с водой, гифы проникают на глубину 160 см и успевают перехватить большую часть этих элементов для своих партнеров-деревьев. Наименьшая обеспеченность растений доступными соединениями фосфора отмечена на песчаных почвах повышенных (лишайниковые боры) и пониженных (сфагновые сосняки) форм рельефа. Однако скорость транспорта питательных веществ из мицелия грибов в ткани растений на бедных почвах гораздо выше, чем на богатых.
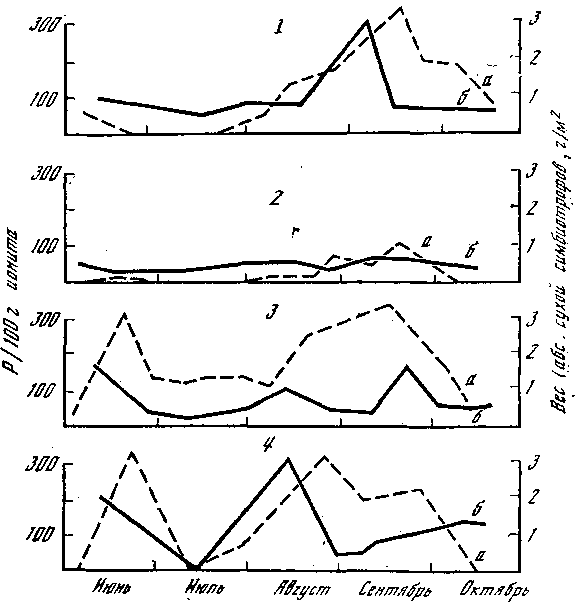
Рис. 6. Динамика фосфора в горизонте А1 дерново-подзолистых почв в массы карпофоров симбиотрофов вблизи эдификатора
1 — ель; 2 — дуб; 3 — береза; 4 — окно; а — фосфор; б — симбиотрофы
Немаловажным фактором для микоризообразования является и обеспеченность почв кислородом, но грибы приспособились к его недостатку путем образования водонепроницаемой пленки на гифах мицелия в условиях почвенного переувлажнения и дефицита влагообеспеченности местообитаний.
Время появления и количество плодовых тел грибов-микоризообразователей, к которым относятся большинство съедобных и ядовитых (березовики, осиновики, грузди, сыроежки, маслята, белые, лисички, мухоморы), в сильной степени зависят от содержания фосфора в лесной подстилке и верхних почвенных горизонтах, а также от его динамики. Максимальные их количества совпадают во времени и в пространстве, о чем свидетельствуют данные, представленные на рис. 6. Работы проводили в смешанном лесу, где отдельные участки находились под влиянием различных пород (эдификаторов), а были такие, где породы древесные выпали из древостоя и образовались окна в древесном пологе. В таких окнах, которые можно считать аналогом лесных опушек, наблюдается два пика максимума, соответствующих раннелетнему и осеннему слоям грибов. Дуб в условиях Подмосковья — маломикотрофная порода, т. е. с ним связано небольшое количество симбиотрофов, и соответственно этому содержание фосфора (Р) под ним имеет вид выровненной кривой.
Способность образовывать микоризу на своих корнях выражена у различных деревьев неодинаково, с одними связаны десятки видов, с другими — единицы. Некоторые виды грибов верны своим партнерам и сопровождают деревья на всем протяжении их географического распространения, что и закреплено в названии грибов (осиновик, березовик, лиственничный масленок, сосновый рыжик); другие не так верны, древесной породе. Совсем нет симбиотрофов у ив, черемухи, рябины, липы, клена, вяза, ясеня. С грабом связаны 2 вида, с ольхой, как и с осиной, — 4, с лиственницей — 8, с пихтой — 27. Иначе обстоит дело с древесными породами, которые в силу своих биологических и экологических особенностей могут занимать крайне различные по почвенно-грунтовым и гидротермическим условиям местообитания. К ним относятся: береза, ель, дуб, сосна. Их способность образовывать микоризу зависит в первую очередь от обеспеченности почв минеральными элементами, доступности для растений питательных веществ и гидротермического режима региона. Показательны в этом отношении березовые леса, распространенные по всей территории СССР от Крайнего Севера до Юга и с Запада до Приморского края. На Севере, где наблюдается лимит или недоступность для корней азота, фосфора, калия, береза чрезвычайно высокомикотрофна и с ней связано около 100 видов симбиотрофов. По мере улучшения почвенно-грунтовых условий ее микотрофность резко падает, достигая минимума на богатых почвах умеренного климата (30—40 видов). С дубом связано от 10 до 100 видов симбиотрофных макромицетов в зависимости от условий обитания, а с елью — 30—70.
Мы рассмотрели, как ведут себя в различных лесах симбиотрофы, непосредственно связанные с корнями древесных пород, а теперь проследим, как ведут себя напочвенные сапротрофы. Формирование их структур в лесных сообществах подчинено закономерностям накопления и разложения мертвого органического вещества, заключенного в лесном опаде и подстилке. Это можно проследить на рис. 7, где представлена схема соотношений различных групп напочвенных сапротрофов в связи с изменениями качественного состава и количества опада и подстилки в возрастном ряду ельников. В молодняках происходит постепенный рост запасов лесной подстилки и процесс ее вертикальной дифференциации (формирование слоев). К 30 годам в ельниках усложняется ее состав, формируется слой погребенных в подстилке трудноразлагаемых фракций (кора, ветки, шишки) и гумусовый слой. С ними связаны соответствующие виды грибов. При внедрении в ельники лиственных пород (дуб, осина, береза) запасы лесной подстилки возрастают. Кроме того, присутствие в опаде листьев существенно меняет темпы разложения хвойной подстилки и влечет за собой появление грибов, специализированных на разрушении мертвого органического вещества лиственных деревьев. Необходимо учитывать и то, что в более старых по возрасту и сложных по набору древесных пород лесах создается огромное количество экологических ситуаций в отношении микроклимата, что вызывает рост видового разнообразия напочвенных сапротрофов.
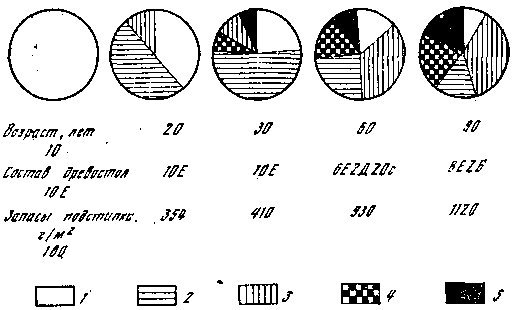
Рис. 7. Формирование структуры напочвенных сапротрофов в возрастном ряду ельников
1 — сапротрофы на опаде; 2 — сапротрофы слоя L; 3 — сапротрофы слоя F; 4 — сапротрофы погребенных фракций опада; 5 — гумусовые сапротрофы; Е — ель; Д — дуб; Ос — осина; Б — береза
Дереворазрушающие сапротрофные макромицеты наиболее независимы в отношении фитоклиматических особенностей определенного лесного сообщества. Количество видов и их численность зависят от запасов отмершей древесины и ее состояния разложения. С увеличением возраста древостоя, количества древесных пород, образующих леса, масса мертвого органического вещества, заключенного в древесине, растет, соответственно увеличивается и видовое разнообразие ксилотрофов.
В среднем для лесов СССР около 40% всех макромицетов принадлежат к группе микоризообразующих, 20% — ксилотрофов и 40% приходится на напочвенные сапротрофы.

