Частная фармакология
Лекарственные средства, влияющие на нервную систему (нейротропные лекарственные средства)
Синаптическая передача является основой деятельности нервных клеток и включает высвобождение нейромедиаторов, которые переносят информацию от пресинаптического – передающего – нейрона к постсинаптической – принимающей – клетке.
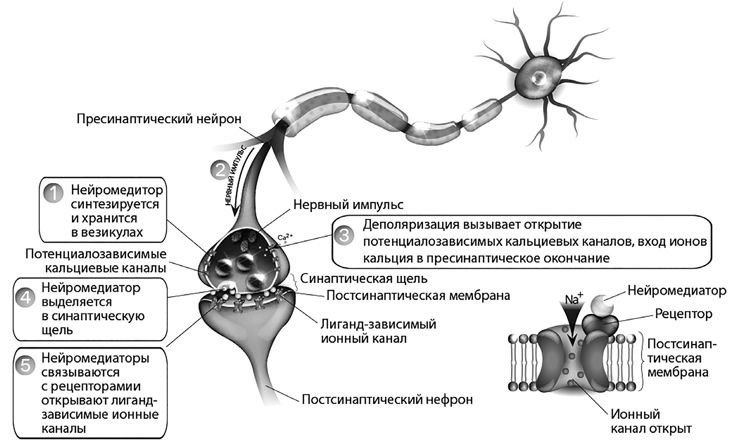
Рис. 11. Факторы, влияющие на проведение возбуждения в синапсе
Синапсы образуются между нервными окончаниями – пресинаптическими окончаниями аксонов (передающего нейрона) и телом клетки или дендритами принимающего нейрона. Внутри окончания аксона находится множество синаптических пузырьков. Это связанные с мембраной сферы, заполненные молекулами нейромедиаторов. Когда потенциал действия или нервный импульс достигает окончания аксона, он активирует потенциал-зависимые кальциевые каналы в клеточной мембране.
Са2+, который присутствует в гораздо большей концентрации вне нейрона, чем внутри, устремляется внутрь клетки.
Са2+ обеспечивает высвобождение нейромедиатора в синаптическую щель. Молекулы нейромедиатора диффундируют через синаптическую щель и связываются с рецепторными белками постсинаптической клетки. Активация постсинаптических рецепторов приводит к открытию или закрытию ионных каналов в клеточной мембране. Это может быть деполяризация или гиперполяризация – в зависимости от задействованных ионов.
В некоторых случаях эти эффекты на поведение канала прямые: рецептор представляет собой управляемый лигандом ионный канал. В других случаях рецептор сам по себе не является ионным каналом, а активирует ионные каналы через сигнальный путь.
Лекарственные средства, влияющие на вегетативную нервную систему
Нервная система анатомически делится на центральную нервную систему (ЦНС: головной и спинной мозг) и периферическую нервную систему (ПНС: нервные ткани вне ЦНС). Функционально нервную систему можно разделить на два основных отдела: вегетативную и соматическую.
Вегетативная нервная система (ВНС) в значительной степени независима (автономна) в том смысле, что ее деятельность не находится под непосредственным сознательным контролем. Эта система связана, в первую очередь, с контролем и интеграцией висцеральных функций, необходимых для жизни, таких как артериальное давление, сердечный выброс, распределение кровотока и пищеварение. Вегетативная нервная система состоит из функционально различных двух частей: симпатической, отвечающей за реакции типа «борись или беги», и парасимпатической, обеспечивающей функции типа «пищеварение и расслабление».
Двигательная часть соматической нервной системы в значительной степени связана с сознательно контролируемыми функциями, такими как движение, дыхание и поза. В соматической нервной системе эта эфферентная часть состоит из одного нейрона, иннервирующего скелетную мышцу.
Вегетативная нервная система, в отличие от соматической, состоит из двух последовательно расположенных нейронов. Тела первых нейронов находятся в ЦНС. Их аксоны, именуемые преганглионарными волокнами, заканчиваются в вегетативных ганглиях, где образуют синаптические контакты со вторыми, ганглионарными нейронами. Передача возбуждения в ганглиях осуществляется с помощью медиатора ацетилхолина, поэтому она называется холинергической, а рецепторы на постсинаптической мембране постганглионарного нейрона называются холинорецепторами. Эти рецепторы, помимо ацетилхолина, возбуждаются также никотином, поэтому называются холинергическими никотиночувствительными рецепторами (н-холинорецепторами).
Аксоны ганглионарных нейронов, называемые постганглионарными волокнами, формируют синаптические контакты с клетками иннервируемых (эффекторных) органов (рис. 12). Исключение составляет вегетативная иннервация хромаффинных клеток мозгового вещества надпочечников, эмбриогенетически родственных нейронам симпатических ганглиев. Эти клетки иннервируются только преганглионарными нервными волокнами.
Центры симпатической иннервации расположены в боковых рогах грудного и поясничного отделов спинного мозга. Ганглии нервной симпатической системы локализованы вне иннервируемых органов – в симпатических стволах, расположенных по обе стороны позвоночника (симпатический ствол) и в симпатических узлах (например, чревное сплетение). Вследствие этих анатомических особенностей строения в симпатической системе преганглионарные волокна короче постганглионарных (см. рис. 12).
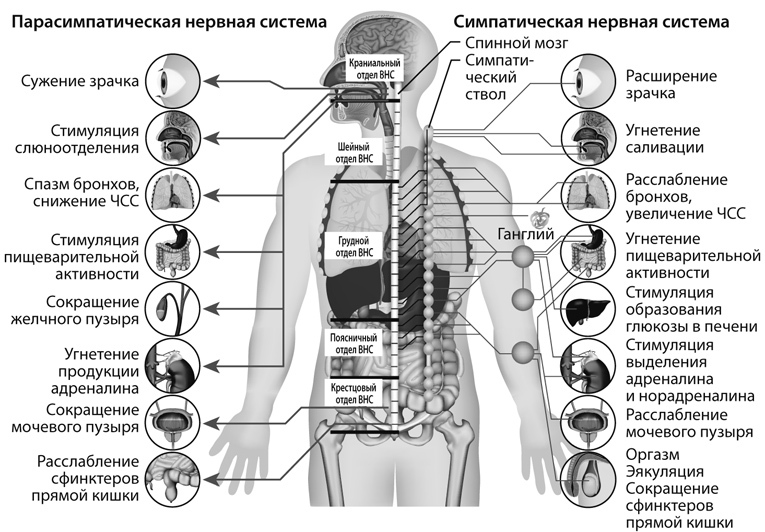
Рис. 12. Схема вегетативной иннервации. (Пояснения в тексте.)
В парасимпатической нервной системе ганглии расположены вблизи органов, поэтому преганглионарные волокна этой системы длинные, а постганглионарные – короткие.
Основное различие между симпатической и парасимпатической нервной системой заключается в том, что в симпатической системе передача возбуждения с постганглионарного волокна на эффекторный орган осуществляется с помощью медиатора норадреналина (адренергическая передача), а в парасимпатической – посредством ацетилхолина (холинергическая передача).
Лекарственные средства, регулирующие функции парасимпатической нервной системы
Центры парасимпатической системы расположены в среднем, продолговатом мозге и крестцовом отделе спинного мозга. Парасимпатические ганглии в основном локализованы в непосредственной близости или внутри эффекторных органов (интрамуральные ганглии), поэтому преганглионарные волокна длиннее постганглионарных (см. рис. 12). В парасимпатической системе медиатором в пре- и постганглионарных волокнах является ацетилхолин.
В холинергическом синапсе синтезированный АЦХ накапливается в везикулах пресинаптических окончаний. Пришедшее по нервному волокну возбуждение вызывает деполяризацию мембраны нервного окончания, открытие потенциал-зависимых кальциевых каналов и вход ионов кальция в пресинаптическое окончание. Ионы кальция в пресинаптическом окончании стимулируют выделение АЦХ в синаптическую щель. После взаимодействия АЦХ с холинорецепторами этот медиатор, в отличие от моноаминов (норадреналин, серотонин, дофамин), не подвергается обратному нейрональному захвату, а разрушается ферментом ацетилхолинэстеразой.
Основные эффекты возбуждения парасимпатической иннервации:
• сужение зрачков (миоз) вследствие сокращения круговой мышцы радужки;
• спазм аккомодации (зрение устанавливается на ближнюю точку видения) вследствие сокращения цилиарной (ресничной) мышцы;
• уменьшение ЧСС (тормозящее действие блуждающего нерва);
• уменьшение скорости проведения импульсов по атриовентрикулярному узлу (ухудшение атриовентрикулярной проводимости);
• снижение сократимости предсердий (но не желудочков, так как в желудочках нет холинергической иннервации);
• повышение тонуса бронхов;
• повышение тонуса гладких мышц ЖКТ, мочевого пузыря (при этом тонус сфинктеров снижается), миометрия;
• увеличение секреции бронхиальных желез, пищеварительных желез (слюнных желез, желез ЖКТ), слезных и носоглоточных желез.
Таблица 5. Расположенные на эффекторных органах холинергические рецепторы
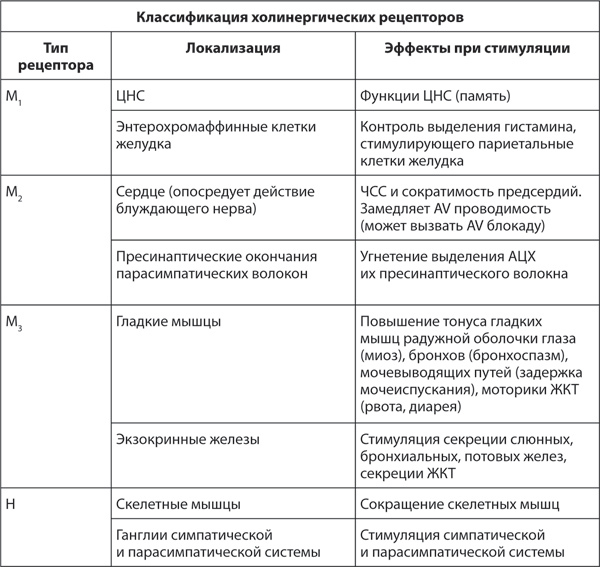
Холинергические рецепторы неоднородны и подразделяются в зависимости от чувствительности к эталонным соединениям (мускарин и никотин) на никотиновые (Н) холинорецепторы, расположенные в ганглиях и нервно-мышечных окончаниях, и мускариновые (М) холинорецепторы, расположенные в постганглионарных синапсах на эффекторных органах. М-холинорецепторы включают 3 подгруппы – М1-М3, – расположенные в различных органах (всего выделено 5 подтипов М-холинорецепторов, однако практическое значение имеют первые три).
Классификация холиномиметиков
Стимуляция передачи возбуждения в холинергических синапсах возможна двумя путями: с помощью прямых агонистов холинорецепторов, называемых холиномиметиками, и за счет ингибирования фермента ацетилхолинэстеразы. Блокада фермента приведет к накоплению собственного ацетилхолина в синаптической щели и стимуляции холинорецепторов.
В соответствии с типами холинорецепторов выделяют следующие группы холиномиметиков:


