Книга: Удивительная эволюция. Биологическая история Земли в невероятных превращениях и мутациях организмов
Назад: Глава вторая Реплицированные рептилии
Дальше: Часть вторая Эксперименты в дикой природе
Глава третья
Причуды эволюции
Пробираясь в зарослях кустарника, ночью по лесу бродит мохнатое маленькое существо, втыкая нос то в одно, то в другое место: оно пытается учуять запах своей мягкотелой добычи. В лесу темно, а зрение у этого эльфа слабое, но длинные усы и острое обоняние позволяют ему ориентироваться в пространстве. В случае опасности он срывается с места с головокружительной скоростью, несясь по лесу и обходя ямки, и вскоре уже исчезает из виду.
Абсолютно уникальный образ жизни. Многие животные ночами бороздят лесную почву похожим образом в поисках мелкой добычи: ежи, землеройки, куницы, а также более крупные животные, такие как опоссумы и даже кабаны. Мир полон ими.
Но это животное особенное. Все другие покрыты шерстью. Его шкурка тоже мягкая, состоящая из миллиона тонких нитей. Но это не шерсть. Все другие животные передвигаются на четырех лапах и производят на свет живое потомство. Но не оно.
Царапая, обследуя, обнюхивая, животное часто действует в паре со своим партнером, подзывая его время от времени и оставаясь с ним в контакте, пока они обходят свою территорию. И когда зовет самец, он выдает себя особенным криком: «ки-ви, ки-ви».
Мы в Новой Зеландии, и это ночное насекомоядное – птица с зачатками крыльев, кошачьими усами, мягкими перьями, и у него, в отличие от всех остальных птиц, на кончике клюва имеются ноздри.
Киви (иллюстрация Дэвида Тасса)
Многие называют их «почетными млекопитающими». Киви – не единственная странная птица из Новой Зеландии. Самыми знаменитыми были гигантские бескрылые птицы моа, некоторые из них возвышались на девять футов над землей и весили шестьсот фунтов. А еще бескрылый попугай; плотоядный попугай, атакующий овец; адзебил, коренастый бескрылый родственник лысухи с массивным хищным клювом и самый крупный за всю историю хищник – орел, размеры которого позволяли ему охотиться на моа. Птицы – не единственные примеры причудливой эволюции. К другим характерным для Новой Зеландии аномалиям относятся вьющиеся кустарники с торчащими наружу ветками и спрятанными внутри листьями; улитки размером с гамбургер и покрытый броней сверчок величиной почти с крысу – пожалуй, самое крупное в мире насекомое. Новая Зеландия изобилует необычными видами.
Но не менее странным является отсутствие там привычных видов животных: млекопитающих. На островах едва ли сыщешь клочок шерсти. Не считая тюленей, лежбища которых можно встретить на уютных новозеландских пляжах, единственным местным видом млекопитающих являются три вида летучих мышей. И даже они необычны. В силу того, что вместо передних конечностей у них крылья, летучие мыши, как правило, довольно неуклюжи на земле. Но не в Новой Зеландии. Самый часто встречающийся наземный вид рукокрылых – короткохвостые летучие мыши – с большим проворством рыскают по лесной почве в поисках насекомых, плодов и нектара. Как точно назвал их знаменитый биолог Джаред Даймонд, это «попытка семейства летучих мышей породить мышь»[41].
Не стоит забывать, что млекопитающие доминировали в наземных экосистемах мира на протяжении последних пятидесяти пяти миллионов лет. Новая Зеландия – это картина альтернативного мира, лишенного млекопитающих. В отсутствии млекопитающих их нишу заняли птицы, взяв на себя те природные функции, которые обычно выполняли млекопитающие, но делая это непривычным образом. При беглом взгляде киви можно перепутать с землеройкой или барсуком. Однако доминировавшие в свое время травоядные – теперь уже исчезнувшие моа и гигантские бескрылые гуси – не шли ни в какое сравнение со стадами антилоп и оленей. И плотоядный попугай, и крупная с тяжелым клювом лысуха – это всего лишь необычная замена привычного нам комплекта хищных видов: кошек, волков, медведей и куниц. На самом деле освобождение от давления хищников, вероятней всего, способствовало увеличению размеров насекомых, улиток и других членистоногих и превращению летучих мышей в грызунов. При повторном прокручивании пленки эволюция мира, где доминируют птицы, может идти совсем иным образом, чем эволюция в условиях доминирования млекопитающих.
НОВАЯ ЗЕЛАНДИЯ не единственное место, где развитие природных форм шло своим особым путем. За исключением карибских анолисов и улиток Mandarina острова изобилуют эволюционными причудами. На Кубе, к примеру, есть свои природные особенности. К сожалению, сова, которая была ростом с первоклашку и могла съесть детеныша гигантского ленивца, исчезла (так же, как и ленивцы, один из видов которых был размером с гориллу). Но остров остается родиной колибри, птички величиной со шмеля; щелезуба, древнего млекопитающего, сошедшего прямо со страниц книг доктора Сьюза.

Щелезуб
У него ядовитая слюна и длинный, гибкий усатый нос. А еще там живут похожие на морских свинок животные размером с бигля, которые лазают по деревьям и оставляют повсюду зеленые, похожие по форме на бананы экскременты.
Даже на крошечных островах есть свои особенные диковины. Остров Лорд-Хоуи площадью пять с половиной квадратных мили, лежащий полумесяцем в Тасмановом море, является родиной шестидюймовых черных «древесных крабов», которые, несмотря на свое название, представляют собой крупных, громоздких представителей семейства палочников. Соломоновы острова, расположенные в южной части Тихого океана, приютили у себя ящерицу, имитирующую поведение обезьяны. Это цепкохвостый сцинк, блестящая, стройная ящерица длиной два с половиной фута с цепким хвостом, которым она страхуется, исследуя кроны деревьев в поисках плодов. Остров Святой Елены, расположенный в южной части Атлантического океана, в первую очередь, известен тем, что там в изгнании жил Наполеон. А несколько десятилетий назад ученые обнаружили на этом острове гигантскую, длиной три с половиной дюйма, уховертку – блестящее черное насекомое с клещами длиной в дюйм, украшающими задний конец ее тела, которое отдаленно напоминает существо из фильма «Звездный путь». И, конечно же, все слышали о дронте, обитавшем когда-то на острове Маврикий в Индийском океане, – бескрылом, бесстрашном, поедающем фрукты попугае размером с индюка ростом в три фута и весом в сорок фунтов.
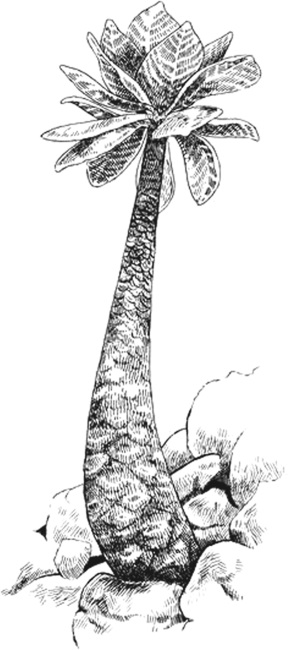
Гавайское растение алула
Однако лидером среди маленьких островов по количеству эволюционных диковинок являются Гавайские острова.
Здесь вы встретите равнокрылых стрекоз, личинки которых живут не в воде, а на суше; прожорливых плотоядных гусениц; плодовых мушек, отказавшихся от своей привычной фруктовой диеты ради гниющей листвы, и другие виды плодовых мушек с головой в форме молота, которые защищают свои территории подобно толсторогим баранам.
Растительный мир Гавайев не менее экстравагантен. И возглавляет список гавайская пальма алула, которая напоминает кеглю, увенчанную салатом-латуком (еще одно ее альтернативное название – «капуста на палке»). Это горное растение высотой в три фута, непохожее ни на одно другое растение в мире[42], как сказал один прославленный ботаник, живет в расщелинах северных склонов островов Кауаи и Молокаи. Благодаря его округлой нижней части растение легко подхватывается сильными океанскими ветрами, а жесткие мясистые листья хорошо приспособлены для сухого и соленого воздуха, в котором обитает растение.
А еще вспомним про Мадагаскар, который иногда называют восьмым материком за специфичность его флоры и фауны. Мы уже обсуждали лягушек и птиц этого острова. Но в этом списке есть и другие необычные виды: карликовый бегемот; адаптивная радиация лемуров, включающая особь весом в семьдесят пять фунтов, которая, вероятно, свешивалась вниз головой, как ленивец. Еще один вид, который выглядел как очень крупная коала; египетская цапля высотой десять футов и весом в полтонны (самые тяжелые птицы из всех когда-либо живущих); хамелеон, который высовывает свой липкий язык на расстояние, в два раза превышающее длину его тела, чтобы захватывать им ни о чем не подозревающее насекомое; ископаемые лягушки размером с большую пиццу; крокодилы-вегетарианцы; жук с длинной шеей.
По своей необычности растения Мадагаскара ни в чем не уступают животным. Например, сухие леса, состоящие из высоких тонких стеблей с выступом на верхушке, или толстый приземистый баобаб, который выглядит так, словно его воткнули в землю вниз головой, и корни торчат сверху. Объединяет животный и растительный мир орхидея с длинной тридцатисантиметровой трубкой в основании цветка и опыляющий ее мотылек с хоботком такой же длины, который в несколько раз превышает длину его тела: он специально приспособлен для того, чтобы втыкать в трубку и дотягиваться до находящегося на дне нектара.
И, наконец, в нашем списке – далеко не окончательном – удивительные виды Австралии: утконос, кенгуру и коала, подобных которым нет нигде в мире.
* * *
О чем же говорят все эти островные диковины? Острова позволяют нам краешком глаза взглянуть на эволюционно альтернативные миры, те миры, которые имели бы право на существование, если бы жизнь приняла иной оборот. Что если бы млекопитающие исчезли с лица земли в конце мелового периода вместе с динозаврами? Новая Зеландия дает намек на то, что могло бы быть. Куда бы привела эволюция приматов, если бы обезьяны и приматы не стали активно развиваться? Достаточно взглянуть на многообразие лемуров, обитающих не где-нибудь, а на Мадагаскаре.
Острова – это гигантская поваренная книга эволюции. И получившееся в итоге «варево» говорит нам о том, что невозможно предсказать, что именно выйдет из печи. Измените ингредиенты или порядок их добавления, подбавьте огня, уберите какой-то продукт, положите одну щепотку соли вместо двух, и вкус получившегося блюда окажется другим. Даже когда используешь один и тот же рецепт, кажущиеся безобидными действия вроде замены одного сорта муки другим или готовки на кухне соседа, а не на своей, могут дать совсем иной результат. Островная поваренная книга изобилует историями вероятностей и случайностей, и многообразие получившихся результатов свидетельствует о том, что предсказать эволюцию кого-то или чего-то на отдельно взятом острове очень сложно.
Нужно просто отправиться туда и убедиться самому, и быть готовым найти практически все.
Конечно, единичные случаи эволюции встречаются не только на островах. Природный мир полон причудливых растений и животных, не имеющих эволюционных параллелей. Возьмем для примера слона: какое еще животное пользуется своим носом, чтобы поднимать с земли предметы, посыпать себя пылью и нежно ласкать члена своей семьи? Или рыба-брызгун, чья система зрения и строение рта позволяют ей точно выстреливать струей воды, способной сбить сидящее на ветке насекомое. Однако в нападении с дальнего расстояния непревзойденными мастерами являются пауки болас, выпускающие длинную шелковую нить с липким комочком на конце, которую они накидывают на свою жертву, как гаучо. Этот шарик приклеивается к любому несчастному мотыльку, в которого он попадет. А теперь перейдем от охотничьей к репродуктивной тактике. Самцы рыбы-удильщика значительно меньше самок. Когда самец кусает самку, он выделяет фермент, сращивая свои губы с ее кожей и сливая их тела в единое целое. В конечном итоге, все остальное тело самца исчезает, оставляя только яички для выполнения главной цели. Ну и, конечно, животные с крупным мозгом, использующие орудия, – двуногие. Биосфера насыщена видами, которые смогли уникальным способом адаптироваться к своему образу жизни.
КОНВЕЙ МОРРИС И ЕГО КОЛЛЕГИ составили длинные списки примеров конвергенции. Но с той же легкостью можно составить сравнительные перечни видов, не имеющих двойников. Нам вполне понятен процесс конвергенции, когда виды адаптируются к похожим обстоятельствам одинаковым способом. Но что же такого особенного в единичных эволюционных случаях? Почему у других видов не развились похожие адаптивные свойства?
Одной из гипотез является то, что эти виды появлялись в уникальном природном окружении. Может быть, они не имели аналогов потому, что никакие другие виды не оказывались в подобных условиях. Возможно, такое объяснение применимо к коалам. Весь образ жизни этих животных связан с обитанием на эвкалиптовых деревьях и поеданием ими листьев, насыщенных ядовитыми составляющими.
Как результат пищеварительная система коал исключительно длинная, обеспечивающая достаточное количество времени для процесса детоксикации и усвоения питательных веществ. Медленное переваривание пищи в сочетании с низкой калорийностью листьев означает, что коалы не жируют, а потому они минимизируют трату энергии, проводя большую часть дня в сонном состоянии. Эвкалиптовые деревья растут только в Австралии, так, может быть, единичность вида коал отражает уникальность их окружения.
Но я подозреваю, что в большинстве случаев не это является объяснением. Утконосы встречаются в реках и озерах Австралии на востоке, где они питаются раками и другими видами водных беспозвоночных, которых они отыскивают, копая речное дно и чуя добычу с помощью электрорецепторов, расположенных на клюве.
Образ жизни утконоса таков, что он вполне возможен во многих других местах помимо Австралии. Речки, в которых они обитают, очень похожи на тот ручей, что протекал за домом моего друга детства в Сент-Луисе. Конечно, в Северной Америке полно кишащих раками речек, многие из которых протекают в тех местах, где климатические условия схожи со средой обитания утконосов. И живущие в них хищники явно не опаснее тех, что обитают в австралийских водоемах. Так где же тогда двойник нашего утконоса?
Почему никого, похожего на утконоса, не появилось в любом другом месте?
Или на кенгуру, или на любое другое животное из перечисленного мной списка, каждое из которых обитает в той среде, которая встречается в других регионах?
Другим объяснением единичных случаев эволюции является то, что естественный отбор либо не такой предсказуемый, либо не такой сильный, как полагают некоторые. То есть даже когда виды находятся в похожих условиях, их развитие может пойти разным путем.
Главной причиной отсутствия конвергенции может быть наличие не одного, а нескольких способов адаптироваться к проблеме, поставленной окружающей средой. Вспомните, как плавают позвоночные животные. Многие используют хвост в качестве опоры, но не все хвосты одинаковы. Рыбьи вертикально приплюснуты и двигаются взад-вперед. Крокодилы плавают так же. Но хвосты китов приплюснуты горизонтально и двигаются вверх-вниз. Другие животные, такие как угри и морские змеи, совершают волнообразные движения всем телом. Отдельные виды птиц, например, бакланы и гагары, способны быстро передвигаться под водой, яростно работая перепончатыми лапами. С другой стороны, некоторые виды плавают, используя видоизмененные передние конечности, такие как ласты у морских львов и крылья у пингвинов. Однако самыми удивительными пловцами являются ленивцы, чьи длинные передние конечности развились у них для долгого свисания вниз головой. Они способны отдаленно имитировать австралийский кроль. Беспозвоночные демонстрируют еще больше способов быстрого перемещения в воде, например, реактивное движение осьминогов и кальмаров.
Этот список различных способов движения в водной среде рождает очевидный вопрос: насколько похожими должны быть признаки двух видов, чтобы считаться конвергентными? У кальмаров и дельфинов очень разное строение тела, помогающее им быстро передвигаться в воде, Тут даже нет сомнений, что они не конвергентны друг другу. Лопастное движение лапами некоторых водных птиц – это еще один неконвергентный способ быстрого перемещения под водой.
Однако другие примеры не настолько однозначны. Допустим, хвостовой плавник у китов и акул похож по внешнему виду и функционированию, но один плавник горизонтальный и двигается вверх-вниз, а другой вертикальный и изгибается вправо-влево. Являются ли эти свойства легкими вариациями на тему конвергенции или же неконвергентными решениями с функционально похожим результатом? Я подозреваю, что большинство людей сочтет горизонтальные и вертикальные хвостовые плавники, по сути, одинаковым решением.
Давайте сейчас вернемся немного назад, к тому признаку, что дает функционально похожий результат, но обнаруживает чуть большие анатомические различия.
Способность летать эволюционировала у позвоночных трижды: у летучих мышей, птиц и птерозавров (крупных рептилий, покоривших небо в эпоху динозавров). У всех трех видов передние конечности трансформировались в крылья, и они летают (в случае с птерозаврами – они летали), по сути, одинаково: взмахивают легкими крыльями, обеспечивая движение вверх и выбрасывание тела вперед.
Но при более внимательном изучении выясняется, что крылья этих летающих позвоночных устроены совсем по-разному. Самым очевидным отличием является сама аэродинамическая поверхность. Птицы используют перья, растущие из костей их крыльев. В противоположность им крыло летучих мышей и птерозавров состоит из тонкой, но очень крепкой кожи, натянутой между костями пальцев и туловищем, а в некоторых случаях крепится даже к задним конечностям. Строение скелета крыльев у этих трех групп летунов также заметно отличается.
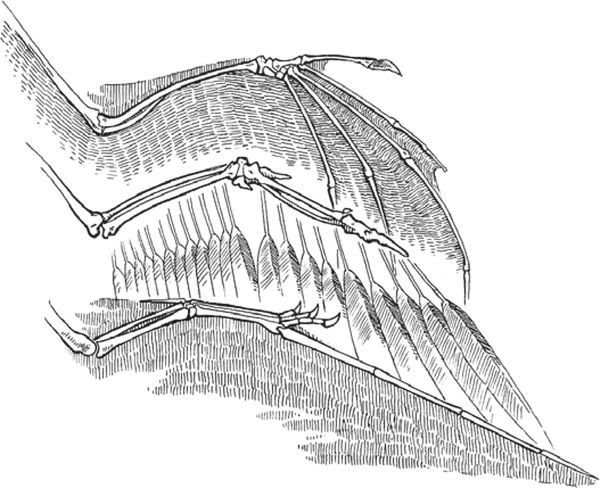
У летучих мышей (сверху), птиц (посередине) и птерозавров (внизу) крылья развились за счет удлинения различных частей передних конечностей. Кроме того, поверхность крыла у летучих мышей и птерозавров состоит из кожи, в то время как птицы пользуются перьями.
Итак, можно ли считать трансформировавшиеся в крылья предплечья птиц, летучих мышей и птерозавров конвергентными адаптациями для управляемого полета, которые только сконструированы разным образом? Или они представляют собой альтернативный, неконвергентный способ эволюции управляемого полета?
Еще один пример. Самой крупной рыбой в океане является шестидесятифутовая китовая акула, названная так потому, что она очень похожа на гладкого кита. Она также является фильтратором, заглатывая своим массивным ртом огромные объемы воды и отфильтровывая крошечный корм. Но на этом сходства заканчиваются. Гладкие киты – голубой, горбатый, серый и другие – процеживают свою добычу, выталкивая воду сквозь жесткие пластинки напоминающих расческу китовых усов, которые образуют завесу, свешиваясь с верхней челюсти. Любые чуть более крупные частицы корма застревают на внутренней поверхности усовой завесы и затем проглатываются. В отличие от них китовые акулы фильтруют свою пищу совсем по-другому. Вода у них выталкивается через жаберные мешки, расположенные с двух сторон на затылке. Фильтрующие подушечки, состоящие из хрящей, размещены в жаберных мешках таким образом, что вода проталкивается между подушечек, проходит через мешки и стекает в океан, а частички пищи продолжают двигаться в противоположном направлении, формируя в глотке массу, которая затем проглатывается. Таким образом, и гладкие киты, и китовые акулы – это крупные морские животные, которые пользуются своим огромным ртом, чтобы набирать воду и отфильтровывать мелкую добычу. Но в то же время сама структура, осуществляющая данную фильтрацию, сконструирована, расположена и функционирует по-разному. Являются ли эти адаптации к фильтрованию пищи конвергентными или неконвергентными?
Где проходит та граница, отделяющая конвергенцию и неконвергенцию, если мы сравниваем структуры в целом похожие и стремящиеся к одинаковому функциональному преимуществу, вопрос субъективный. Я склонен считать крылья птиц, летучих мышей и птерозавров конвергентными друг другу. Точно так же я считаю гладких китов и китовых акул в целом конвергентными, потому что и у тех, и у других крупная ротовая полость, и они отфильтровывают планктон. Но, по моему мнению, их фильтрующие структуры – это неконвергентные, альтернативные адаптации для фильтрации пищи.
Но, на самом деле, в подобных случаях не существует правильного или неправильного ответа.
Тем не менее в других случаях виды способны адаптироваться, развивая явно отличные, неконвергентные фенотипы, обеспечивающие те же самые функциональные способности. Мой любимый пример данного феномена связан с подземным образом жизни грызунов. Свыше двухсот пятидесяти видов крыс проводят большую часть своей жизни под землей, перемещаясь по построенным ими тоннелям. У грызунов подобная модель поведения эволюционировала неоднократно, но совершалось все разными путями. Большинство грызунов роют землю стандартным способом, рыхля ее передними конечностями и отбрасывая назад. Передние конечности у подобного вида крепкие и очень мускулистые, а когти длинные и сильные. Другие виды пользуются зубами, а не когтями, откапывая землю. Как можно догадаться, зубы у этих животных длинные и выступающие, даже по стандартам грызунов, а челюстные мышцы и череп массивные и прочные. Большинство «зубных» копальщиков отбрасывают землю назад передними конечностями. Но некоторые виды плотно утрамбовывают откопанную землю в стену тоннеля, толкая землю вверх своими удлиненными, похожими на лопаты носами. Анатомические различия этих копальщиков четко иллюстрируют неконвергентные адаптации, которые дают функционально одинаковый результат.
Неконвергенция может возникнуть и по другой причине. Зачастую существуют разные функциональные способы адаптироваться к окружающим условиям. Для примера рассмотрим, как потенциальная жертва может адаптироваться к присутствию рядом такого хищника, как лев. Один из вариантов – развить у себя превосходные спринтерские способности, чтобы легко убежать от них. Но существуют и другие альтернативы, такие как маскировка, пассивная или активная защита. Получившиеся в итоге способы адаптации будут решительно неконвергентны друг другу, так как к ним относятся рога африканского буйвола, панцири панголина и черепахи, длинные ноги импалы, иголки дикобраза, яд и точность броска ошейниковой кобры и пестрая шкура бушбока.
Множественные решения одинаковой селективной проблемы не ограничиваются вопросами защиты. Гепарды и гиеновидные собаки охотятся на одинаковых животных, но кошки осуществляют это короткими и быстрыми рывками, в то время как гиеновидные собаки бегают медленнее, но дольше, лишая свою жертву сил и, в конечном итоге, сбивая ее. Соответственно, адаптации двух этих животных отличаются: очень длинные лапы и гибкая спина гепарда позволяют ему развивать скорость до семидесяти миль в час. А высокая выносливость гиеновидных собак помогает им сохранять стабильный темп в тридцать миль в час в течение достаточно продолжительного времени, чтобы измотать свою жертву (гепарды же способны лишь на короткие спринтерские дистанции).
Или рассмотрим, к примеру, способы адаптации животных к добыче нектара. Мы знаем, что растения вырабатывают сладко пахнущую сахаристую жидкость, чтобы приманивать насекомых, птиц и других животных для того, чтобы помочь им осуществить свой репродуктивный процесс. Когда животное засовывает морду или все тело в цветок, чтобы глотнуть нектара, оно покрывается пыльцой. Когда оно переходит к другому цветку, часть пыльцы слетает и оплодотворяет семязачаток растения.
У многих цветов имеются очень длинные трубки с нектаром на дне: так растение может ограничить доступ к пыльце до одного или нескольких определенных видов, хорошо адаптировавшихся к использованию этого растения, таких как мотыльки с их длинными хоботками и колибри с похожими клювами и языками. В силу своей адаптации подобные виды, вероятно, не пользуются многими другими видами цветов, сводя к минимуму вероятность того, что пыльца попадет на растение другого вида и будет зря израсходована.
Но не все нектароядные играют по правилам. Некоторые виды насекомых, птиц и млекопитающих выедают отверстие в основании цветка, обходя лепестки и пыльцу и, таким образом, не доводя до завершения эту совместную эволюционную сделку. Чтобы проделать подобное, воришки нектара овладели самыми разными способами адаптации. У них нет длинных языков и специальных приспособлений, чтобы добраться до дна трубок растения, но зато у данных видов благодаря возникшим адаптивным свойствам развилась способность разрывать стенку цветка.
У некоторых колибри на клювах имеются зазубрины специально для этой цели. У птиц, названных соответственно – крючкоклювы, – имеется острый крючок на кончике, чтобы разрезать цветок.
Во всех этих многочисленных примерах мы видим, что существуют разные эволюционные варианты, формирующиеся в ответ на поставленную окружающей средой проблему. Но сам факт наличия массы возможностей вовсе не означает, что все или хотя бы несколько таких возможностей смогут развиться. Конвей Моррис и команда утверждают, что один такой возможный вариант будет превосходить другие. Вот почему похожий признак эволюционирует конвергентно, снова и снова. И все же конвергенция возникает не всегда. Почему бы естественному отбору не проталкивать каждый раз один и тот же признак?
Бывает так, что два (или больше) признаков эквивалентны. Маскировка или побег на большой скорости помогут одинаково успешно избежать хищников. Или, может быть, один подход эффективнее другого для какой-то конкретной цели, но при иных затратах, которые уравновешивают его преимущество. Стремительно нестись прочь от хищника – возможно, и лучший способ спастись, но маскировка может усилить способность такого животного, как змея, подкарауливать собственную жертву. Когда речь идет о выживании и сохранении потомства, то тех животных, которые замаскированы, может ждать такой же успех, как и тех, которые полагаются на скорость в процессе репродукции и передачи своих генов следующему поколению. В результате естественный отбор не обязательно отдаст предпочтение тому или другому варианту. То, какой признак разовьется, может стать делом случая, благодаря которому мутация происходит сначала у популяции, которая становится объектом нападения хищников.
Или же, как вариант, то, какой признак разовьется, может быть контингентно изначальному фенотипу и генотипу вида. Вид, который был в целом активен, может быть предрасположен к развитию у него тех признаков, которые обеспечивают большую скорость, при условии, что он сталкивается с новым хищником, в то время как у более оседлого вида может развиться лучший способ маскировки. Ни один из вариантов не превосходит другой, но эволюционный итог может сильно зависеть от первоначальных условий.
Может быть и так, что одно решение действительно превосходит другое, но в некоторых случаях легче выработать приближенное к оптимальному решение. Французский ученый Франсуа Жакоб[43], получивший Нобелевскую премию за свое исследование работы ДНК, предложил аналогию, объясняющую, почему естественный отбор не всегда приводит к эволюции идеально созданного организма. Естественный отбор, говорил Жакоб, это не инженер, который вырабатывает оптимальное решение проблемы. В данном случае, продолжал он, лучше представить себе умельца-самоучку, мастера на все руки, который использует любые доступные материалы, чтобы изготовить любой приемлемый вариант – не лучшее, а наиболее достижимое в данных обстоятельствах решение.
Теперь представьте себе вид птиц, который оказывается в местности с озером, кишащим рыбой. Они могут начать нырять в воду за рыбой, а со временем начать адаптироваться к водной среде: у птиц начнут формироваться крупные и мощные задние лапы, как у баклана, или трансформироваться крылья, как у пингвинов. Предположим, что самый лучший способ быстро и проворно плавать – это задействовать в воде сильный и мускулистый хвост, двигая им взад-вперед или вверх-вниз, именно это делают самые быстрые пловцы. Но у птиц нет длинных хвостов: они потеряли их ранее в процессе своей эволюционной истории, более сотни миллионов лет назад, сохранив лишь крошечные остатки сплавленных костей («хвосты» птиц состоят только из перьев, а не из костей). Я не говорю, что повторная эволюция длинного хвоста невозможна, но естественный отбор, этот мастеровой, вероятно, не пойдет таким путем. У птицы уже есть крылья, и ее лапы способны обеспечить ей необходимый проталкивающий импульс. Гораздо более вероятным выглядит тот вариант, что естественный отбор, скорее, будет способствовать улучшению ее плавательных умений за счет уже существующих структур, чем развивать новую способность с нуля, даже если в конечном итоге перекроенная птица с костистым хвостом – что напоминает скрещение гагары и крокодила – может оказаться лучшим пловцом.
Но, все равно, если птица-крокодил будет лучше адаптирована – станет лучшим и быстрым пловцом, – тогда почему бы плавающей птице не продолжать развиваться в этом направлении? Ответом может быть, что переход от одной адаптивной формы к другой может быть трудным, так как промежуточные условия бывают хуже. Длинный сильный хвост может быть великолепен для быстрого проталкивания тела, но короткий шлепок хвоста может просто помешать движению, фактически уменьшая плавательную результативность. У естественного отбора нет дара предвидения: он не будет отдавать предпочтения негативному свойству только потому, что это ранний шаг на пути, ведущему к лучшему в конечном итоге свойству. Напротив, чтобы определенная черта развилась в ходе естественного отбора, каждый шаг на этом пути должен быть улучшением того, что было раньше: естественный отбор никогда не отдаст предпочтение худшему свойству, даже если это всего лишь переходный эволюционный этап.
Как следствие, виды могут в конечном итоге застрять на уровне близких к оптимальным адаптациях. Какой бы ни была причина, но их предки не выбирали лучшую дорогу к адаптации. Естественный отбор заставлял вид развиваться, и он, в конце концов, приспосабливается, но не так хорошо, как мог бы. Такой ход рассуждений подчеркивает роль, которую может играть контингентность в определении направления эволюции и в том, почему в результате виды могут пойти разными путями, столкнувшись с идентичными природными условиями. Различия видов в их родовом генотипе и фенотипе или в том, какая мутация произойдет первой, могут заставить виды адаптироваться по-разному и даже иногда привести к ухудшенным адаптациям.
ПО ТОЙ ЖЕ САМОЙ ЛОГИКЕ можно ожидать, что чем больше похожи два родственных вида, тем вероятнее, что они будут развиваться похожим образом, когда столкнутся с одинаковыми избирательными условиями. Именно это и происходит. И нет никакого совпадения в том, что лучшие примеры повторяющейся конвергенции можно найти среди близкородственных видов. У ящериц анолисов один и тот же набор представителей рода развивался четыре раза, и при этом ни один другой тип островной ящерицы не конвергировал с анолисами. Два практически идентичных вида носатой энгидрины относятся к одному роду.
Приклеивающиеся подушечки пальцев у геккона эволюционировали одиннадцать раз и всего лишь два раза среди более шести тысяч других видов ящериц. Не все случаи конвергенции[44] охватывают ближайших эволюционных родственников, но недавний статистический анализ подтвердил, что конвергенция больше распространена среди схожих видов.
Близкородственный эффект особенно очевиден при сравнении популяций одного и того же вида, который часто повторно развивает один и тот же признак, сталкиваясь с похожими условиями обитания. В первой главе я привел много примеров: мыши в песчаных дюнах, пещерные рыбы, ядовитые тритоны и их хищники, подвязочные змеи, люди. И еще один пример.
Трехиглая колюшка[45] – это маленькая рыбка, обычно около двух дюймов в длину, которая обитает в прибрежных водах северной части Северного полушария. Самой отличительной характеристикой этой тонкой рыбки, как можно догадаться по ее названию, являются три длинных иглы, расположенные в ряд вдоль хребта перед спинным плавником. А снизу, там, где должен быть брюшной плавник, находится еще одна игла.
Вероятно, хищники представляют серьезную угрозу для этих обитателей океана, потому что кроме того, что эти иглы могут вставать дыбом, бока рыбы защищены броней в виде костистых щитков. У некоторых особей их количество достигает сорока штук.
Во время последнего ледникового периода большая часть Северного полушария была скрыта ледниками. Когда примерно десять тысяч лет назад лед растаял, в океан стекли новые потоки рек. Как и лосось, колюшка размножается в пресной воде, и местные популяции быстро использовали преимущества этих новых мест для нереста.
Но потом ландшафт снова изменился. Когда толща льда высотой в милю накапливается на земле, то поверхность прогибается под его весом. Но как только лед исчезает, земля медленно возвращается в исходное положение. И когда подобное произошло на территории нынешней Канады, часть водных потоков оказалась отрезанной от океана, в результате чего образовались озера. В итоге в этих озерах осталась бывшая ранее морской колюшка.
Подобный процесс происходил с бессчетными тысячами рек, ручьев и протоков, в особенности вдоль западного побережья Северной Америки. Эти водоемы были в геологическом отношении новыми образованиями, недостаточно заселенными: лишь несколько других видов океанической рыбы составили компанию колюшке. Как итог эти новые озерные популяции оказались в абсолютно новых для себя условиях обитания там, где практически отсутствовали хищные рыбы.
Как следствие этого, популяции озерной колюшки, каждая из которых была изолирована в своей собственной «лоханке» и эволюционировала независимо от других, менялись параллельно. Зачем терять энергию и ресурсы, строя защиту против несуществующих хищников? Популяции конвергентно потеряли большую часть своей брони, и их иголки усохли. Генетические исследования показали, что эта параллельная эволюция затронула геном. Во всех озерных популяциях одинаковые генетические изменения воздействовали на эволюцию брони и иголок.
Распространенность конвергенции среди близкородственных популяций и видов вполне понятна. Близкие родственники, как правило, похожи генетически, так что естественный отбор, вероятней всего, будет задействовать одинаковые генетические системы. Более того, родственники чаще похожи во многих фенотипических признаках.
Из-за этих похожестей близкородственные виды и популяции имеют одинаковые эволюционные предрасположенности, которые с большой долей вероятности будут развиваться так, а не иначе. Некоторые эволюционные биологи считают эти предрасположенности привязками или эволюционными тенденциями, которые могут действовать несколькими разными способами. Самый очевидный – это генетическая похожесть близких родственников, но при этом могут возникнуть более незаметные качества. Эволюционировавший у предка признак может мешать каким-то эволюционным вариантам, в результате чего развитие среди потомков данного вида произойдет ограниченным количеством других способов. Либо же предок сможет развить у себя признак, который проложит путь для эволюции второго признака.
Подобное потенцирование, как называют сейчас данный феномен молекулярные биологи, будет обладать таким эффектом, что у всех близкородственных видов разовьется второй признак, тот, который вряд ли возникнет у вида, произошедшего не от этого предка.
На основе всех этих факторов родственные виды, вероятней всего, конвергентно разовьют одинаковые качества, столкнувшись с похожими селективными требованиями. Но это вовсе не означает, что дальние родственники не могут конвергировать. Это, конечно же, происходит, только менее часто.
СЕЙЧАС САМОЕ ВРЕМЯ ненадолго отклониться от темы и подчеркнуть, что конвергенция не обязательно отражает адаптацию к одинаковым обстоятельствам или вообще является результатом адаптации. Причина в том, что естественный отбор – не единственный процесс, заставляющий признаки эволюционировать. Периодически характерные черты эволюционируют случайным образом, в частности, в маленьких популяциях. Признак также может развиться, потому что генетически он связан с другим признаком, который предпочел естественный отбор, или как результат устойчивой миграции из другой популяции. Как следствие, конвергентная эволюция может происходить по случайному совпадению, если у двух популяций развились одинаковые свойства, не связанные с адаптацией. Подобная неадаптивная конвергенция может быть наиболее превалирующей среди родственных популяций или видов из-за их общих эволюционных предрасположенностей.
Примером здесь могут служить пальцы саламандры. У большинства саламандр конвергентно эволюционировали пальцы: их четыре вместо стандартного наследственного комплекта из пяти пальцев. Количество пальцев[46] у взрослой саламандры определяется тем, сколько клеток участвует в процессе формирования конечностей на стадии эмбрионального развития. Все, что сокращает число клеток в зачатке конечности – например, увеличение размера клетки или общее уменьшение размера тела, – может привести к сокращению количества пальцев. У нас нет доказательств, что подобное конвергентное эволюционное сокращение было вызвано естественным отбором: четырехпалые виды не появляются в каких-то определенных средах обитания, и нет никакого преимущества в том, чтобы иметь меньше пальцев (насколько нам известно).
Наиболее вероятное объяснение заключается в том, что уменьшение количества пальцев происходило конвергентно по причинам, не связанным с адаптацией. Возможно, у отдельных видов произвольно развились более крупные клетки, а другие были выбраны за маленький размер тела.
В идеале нам следует напрямую проверить гипотезу о том, что естественный отбор направлял конвергенцию[47]. Соответствующие данные можно получить с помощью прямой оценки естественного отбора, детального анализа преимуществ, которые дает признак, если таковые есть, и знания эволюционной истории вида. Даже простое наблюдение, что признак эволюционировал неоднократно в похожих условиях окружения, наталкивает на мысль об адаптационной подоплеке: связь между развитием свойства и окружением не прослеживалась бы без вовлечения естественного отбора. Но, к сожалению, иногда у нас нет никакой соответствующей информации.
Возьмем для примера тираннозавра рекса. Каким бы устрашающим и грозным ни был этот деспотический властелин, у него имелся один недостаток: его лапы. Худосочные, с двумя пальцами: своими передними конечностями он не мог дотянуться даже до собственного рта. Ученые выдвигали самые разные предположения, одно безумнее другого. Может быть, суперхищник ел в такой безумной спешке, что его лапы укоротились в процессе эволюции, чтобы он случайно не откусил их и не съел. Возможно, маленькими конечностями было удобно отталкиваться от земли, чтобы встать после сна. А может быть, самцу тираннозавра более короткие конечности нужны были для того, чтобы щекотать своего партнера. Нет нужды говорить, что ни одна из этих идей не получила поддержки.
Недавно палеонтологи обнаружили новый вид тероподов, Gualicho shinyae, у которого было похожее слабое место в виде двухпальцевых отростков. И хоть мы не знаем истинной причины, почему этот признак развился у обоих видов, один из авторов статьи говорит: «Очевидно, имело место некое адаптивное преимущество[48], потому что мы обнаруживаем данный признак множество раз у разных видов тероподов».
Но, может быть, это не столь ясно. Конвергентная эволюция совсем не обязательно доказывает, что общий признак является результатом естественного отбора. Возможно, у тираннозавра рекса и Gualicho shinyae случайно развились уменьшенные передние конечности.
Если бы мы знали, почему эволюционировали маленькие конечности с двумя пальцами, какое преимущество они давали или почему естественный отбор благоволил им, у нас было бы основание считать, что конвергенция была адаптивной. Но в отсутствии каких-либо данных мы не можем просто допустить, что причиной является естественный отбор.
В ЗАКЛЮЧЕНИЕ хочу привести последний пример исследования, который демонстрирует несколько разных путей к эволюционной неконвергенции. Данный пример касается видов, которые питаются личинками насекомых, живущих в дереве. Всем знаком перестук, когда дятел долбит отверстие в стволе дерева, нанося удары со скоростью до двадцати ударов в секунду. Но чего многие не знают, так это того, как дятел вытягивает личинку из дерева. А делает он это, вставляя свой длинный колючий язык – такой длинный, что когда он им не пользуется, язык лежит в свернутом виде сзади в черепной полости – глубоко в отверстие, цепляя свою добычу колючками и вытаскивая ее.
Это, несомненно, весьма искусный трюк, но оказывается, что представители семейства дятловых не имеют монополии на такое добывание личинок. Дятлы распространены практически по всему свету, но они не смогли пересечь океан, а потому их нет в Австралии и на большинстве островов. В их отсутствие другие виды эволюционировали и заняли нишу охотников на древесных личинок, но никто не делал это так, как дятлы. В данном случае вместо конвергенции мы наблюдаем набор различных решений проблемы извлечения насекомых из дерева.
На Гавайских островах обитает серпоклювка, симпатичная птичка с необычным клювом. Голова у самцов желтого цвета, а у самок – оливкового. Следует отметить, что верхняя и нижняя части клюва совершенно разные. Нижняя – короткая, прочная и прямая и используется для выкапывания отверстия, в стиле дятла. Но вместо того, чтобы пользоваться языком для извлечения личинок, «акиаполаау» имеет тонкий и заметно согнутый вниз верхний клюв, который в два раза длиннее нижнего и может проникать глубоко в отверстие, чтобы выудить оттуда личинку.
В Новой Зеландии подобная многогранность не по силам любой птице, а потому гуйя разноклювая пошла другим курсом, поделив задачи между полами. Самец брал на себя основные дятловские функции: у него крепкий клюв, которым он пользовался для выдалбливания гниющего дерева в поисках личинок. У самки же обе половинки клюва, похожие на верхний клюв серпоклювки, тонкие и сильно загнуты книзу. Ими она извлекала добычу из глубоких трещин. Какое-то время считалось, что пары птиц действовали, как одна команда: самец бурит, а самка достает. Но это предположение оказалось в итоге неверным прочтением исходного научного отчета. Согласно новой версии, самцы и самки охотились по отдельности. Как это ни прискорбно, но данный вид был истреблен в прошлом веке, так что более детальные исследования уже невозможны.
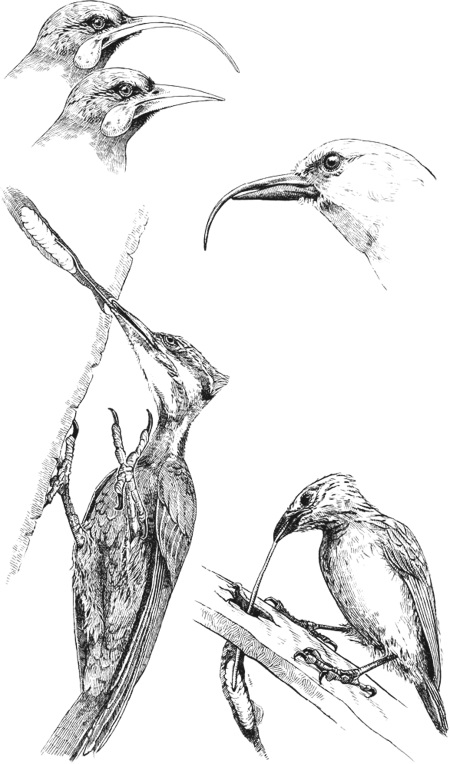
Разные способы адаптации к поеданию личинок (сверху): разноклювая гуйя, серпоклювка, дятел и дятловый древесный вьюрок.
Пожалуй, самый примечательный способ птичьей адаптации к подобному образу жизни демонстрирует птица с наименее примечательным клювом. У дятлового древесного вьюрка с Галапагоса, представителя семейства дарвиновского вьюрка, абсолютно стандартный несогнутый клюв, ни особо тонкий, ни особо прочный, ни длинный, ни короткий, ни достаточно жесткий, чтобы выстукивать дерево, ни достаточно чувствительный, чтобы прощупывать поверхность. Однако это не играет никакой роли, потому что дятловый древесный вьюрок не пользуется клювом для извлечения добычи. По крайней мере, пользуется не напрямую. Он, подобно шимпанзе, что выуживают термитов из гнезда, держит в клюве веточку правильного размера, засовывая ее в отверстие или трещину и раскачивая, выманивая, прощупывая и разведывая, пока, наконец, не уговорит личинку выбраться наружу, и тогда быстро съедает ее. И точно так же, как шимпанзе (а также как вороны Новой Каледонии и, конечно же, мы), эти вьюрки не используют слепо любые веточки, которые подберут. Скорее, они внимательно выбирают орудие, иногда идеально обстругивая и приспосабливая его специально для выполнения конкретной задачи.
Вы, конечно, можете сделать вывод, что выкапывание личинок из деревьев – чисто птичья работа, и ошибетесь. На Мадагаскаре, на этом острове эволюционных чудес, животные, извлекающие личинок из дерева, могут быть самыми диковинными.
Там ниша дятла занята не птицей, а приматом. И каким приматом! Размером с домашнего кота, ведущий ночной образ жизни, ай-ай выглядит, как герой фильма ужасов. Сверкающие желтые глаза, крупные, мягкие, черные кожистые уши на светлой морде, огромный лоб с узким коротким носом, редкая серая шерсть, беспорядочно растущая сверху и сбоку головы, – животное кажется зловещим скрещением Альберта Эйнштейна и Йоды. Но, в отличие от великого физика и мастера джедай, у ай-ай имеется пара огромных, постоянно растущих резцов и свойство, порождающее ночные кошмары и объясняющее веру древних малагасийцев в магические способности этих существ – удлиненный тонкий средний палец, способный вращаться в любом направлении.
То, каким способом ай-ай достает личинку, и вправду поразительно. Он начинает с того, что простукивает ствол дерева своим длинным пальцем. Крупные уши работают, как локаторы, расшифровывающие отражение звука: так животное прислушивается к сигнальной перкуссии пустого внутреннего пространства в дереве. Как только потенциальный тоннель личинки обнаружен, ай-ай задействует свои выступающие вперед резцы, прогрызая ими дерево, чтобы добраться до внутренней пустоты.

Ай-ай
После того, как полость будет открыта, к процессу подключается длинный палец, закручивающийся сначала в одну сторону, а потом в другую до тех пор, пока ноготь не подцепит и не вытянет личинку. Зачем нужен диковинный клюв, когда длинный палец и зубы могут легко проделать эту работу?
Почему естественный отбор предлагал различные решения одной и той же проблемы? По-видимому, личинки насекомых в разных местах разные. И лучшим способом поймать материковых личинок – действовать, как дятел; в то время как галапагосских лучше всего выуживать веточками; а малагасийских проще всего найти приматам с крупными ушами. Мы не можем исключать такой наивной возможности, что у каждого вида развился свой уникальный оптимальный способ ловить местных личинок, но в данном случае более вероятными кажутся два возможных альтернативных объяснения.
Согласно одному сценарию различия эволюционировали случайно. Возможно, у предка дятла случилась мутация, в результате которой сформировался длинный колючий язык, в то время как у прародителя дятлового древесного вьюрка возникло изменение, которое заставила его подбирать веточки и втыкать их в отверстия. Другими словами, ни один из путей не превосходил другой, и то, какая мутация происходила, было лишь делом случая.
Второй сценарий обусловлен эволюцией: то, как вид реагирует на естественный отбор, зависит от того, как он развивался в прошлом. Возьмем для примера ай-ай, представителя семейства лемуров. У приматов, как у всех млекопитающих, есть рты, состоящие из костей, кожи, мышц и, как правило, зубов. Развить жесткий, острый клюв, как у птицы, было бы для млекопитающего настоящим эволюционным подвигом. Это гораздо более трудный генетический процесс, чем видоизменение уже существующих резцов для того, чтобы прогрызать ствол дерева. С другой стороны, у птиц передние конечности трансформировались в пригодные для полета структуры, и им уже недоступны пальцевые кости, работающие как крючки у ай-ай.
_____
ИТАК, К ЧЕМУ ЖЕ МЫ ПРИШЛИ? Является ли конвергенция превалирующим явлением, демонстрацией наследственной структуры в биологическом мире, направляемой предсказуемыми силами естественного отбора к итогам, предопределенным условиями окружающей среды? Или же примеры конвергентной эволюции – это исключения, тщательно подобранные иллюстрации биологической предсказуемости в беспорядочном мире, в котором большинство видов не имеют эволюционных параллелей?
Мы можем спорить на эту тему и так и эдак до посинения. Я бы привел в качестве примера утконоса. Но вы бы парировали конвергентными ежами. Я бы обусловил существование уникального, висящего вниз головой ленивца, а вы бы возразили мне, приведя в качестве доказательства ходящую на двух ногах мышь, эволюционировавшую независимо на трех материках. Вот именно так, по сути, происходил исторический спор с накапливающимися списками и примерами.
Конвея Морриса и его коллег следует похвалить за то, что они сделали актуальной проблему конвергентной эволюции. Все мы знали, что конвергенция – это уловка естествознания, поразительный пример силы естественного отбора. Но Конвей Моррис и компания прояснили тот факт, что эволюционное копирование гораздо более распространенное явление, чем мы себе это представляли. Сейчас мы понимаем, что подобное часто случается в природе, и примеров этого полно вокруг нас. И все же оно далеко не повсеместно. Чаще виды, обитающие в схожих условиях, не адаптируются конвергентным образом.
В этом месте мы должны выйти за рамки описания исторической модели с перечислением дополнительных примеров за и против. Скорее, нам следует задаться вопросом, понимаем ли мы, почему конвергенция происходит в одних случаях, а не в других; как найти ту границу, где конвергенция случается или нет; почему двуногие грызуны эволюционировали независимо в пустынях по всему свету, а кенгуру только один раз. И чтобы сделать это, требуется нечто большее, чем добавление новых примеров к нашим спискам.
Нам нужно проверить гипотезу эволюционного детерминизма напрямую.
На протяжении последнего века экспериментальный подход был стандартным для большинства научных дисциплин, и вполне обоснованно. Аккуратно меняя один параметр и оставляя другие неизменными, мы можем напрямую протестировать причину и следствие. Неэкспериментальные исследования хромают без отсутствия контроля и от того, что любой из многочисленных параметров может быть ответственным за наблюдаемые между объектами исследования различия.
Но эволюционная биология поздно включилась в экспериментальную игру: легендарный медлительный ход эволюции сделал идею экспериментов провальной затеей. Теперь мы знаем, что данная точка зрения ошибочна и эволюция способна двигаться очень быстро. И понимание этого открывает новые двери к ее изучению.
Итак, мы перетрясли ящики естествознания, оглядываясь назад, чтобы понять, что возникало в прошлом. Но теперь настало время заглянуть вперед, чтобы использовать силу экспериментального метода для изучения эволюционной роли контингентности и детерминизма.

