Книга: Эмоциональный мозг
Назад: Подражательное поведение как пример компенсаторной функции эмоций на популяционном уровне
Дальше: Анализ критических замечаний в адрес информационной теории эмоций
Подкрепляющая функция эмоций на популяционном уровне: феномен эмоционального резонанса
Многократно показано, что животные способны реагировать на внешние проявления эмоционального состояния другой особи своего, а подчас — и другого вида. Более того, эти сигналы могут играть роль подкрепления при выработке инструментальных условных рефлексов, в том числе у крыс [Rice, Gainer, 1962; Greene, 1969].
Реакцию избегания при болевом раздражении другой особи мы вырабатывали у 267 взрослых белых крыс-самцов. В этих опытах было использовано экологически характерное для крыс предпочтение находиться в ограниченном пространстве. В установке, сконструированной инженером В. А. Пучковым (рис. 7), пол плексигласового «домика» представлял педаль, автоматически включавшую счетчик времени и болевое раздражение электрическим током лап другой крысы, находившейся за тонкой прозрачной звукопроницаемой перегородкой. Исследуемое животное один раз в день помещали в открытую, относительно просторную часть установки на 5 мин и регистрировали время его пребывания на педали. В течение 10 (в некоторых сериях опытов 5) дней вход в «домик» не сопровождался болевым раздражением второй крысы, в то время как на протяжении следующих 10 дней каждое появление исследуемого животного в «домике» приводило к включению тока силой 1–2 мА. Раздражение «жертвы» продолжалось 3–5 с, с пятисекундными интервалами, до тех пор пока исследуемая крыса находилась на педали.
Рис. 7. Установка В. А. Пучкова для изучения реакции избегания у крыс при болевом раздражении другой особи
1 — свободное пространство; 2 — «домик» с педалью; 3 — помещение «жертвы».
Специально поставленные опыты со звуковой и зрительной изоляцией крыс показали, что в комплексе сигналов оборонительного возбуждения «жертвы» решающее значение для исследуемого животного имеет крик. Вот почему мы меняли партнеров в случайном порядке, а крыс — «жертв», склонных к пассивно-оборонительной реакции «замерзания», с хриплой или беззвучной вокализацией выбраковывали и заменяли другими. При угашении выработанной условной реакции избегания ток выключали, однако крыса — «жертва» по-прежнему оставалась в камере.
Оказалось, что на протяжении 2–3 (редко 6–8) опытов время пребывания на педали у 85 (то есть примерно у 32 %) исследованных животных становилось короче — 1 мин 30 с, а среднее число появлений в «домике» не превышало 3,1 (см. крысу № 13 на рис. 8). Некоторые крысы этой группы вообще перестали заходить в «домик», хотя большую часть времени проводили около входа на педаль. Среди остальных 182 крыс можно было выделить животных, находившихся на педали почти все 5 мин экспозиции, а также крыс, непрерывно перебегавших из открытого пространства в «домик» и выбегавших обратно при крике раздражаемой током «жертвы».
Нас интересовали три вопроса:
1) какие характерные черты присущи другим формам зоосоциального поведения крыс, различающихся по их реакциям на болевое раздражение партнера?
2) каким образом и в каких пределах можно изменить реакцию крыс на крик боли, издаваемый другой крысой?
3) повреждение каких отделов головного мозга и в каком направлении влияет на изучавшуюся нами реакцию избегания?
Ниже приводятся результаты опытов, призванных ответить на эти вопросы.
Ранее [Симонов, 1976] мы показали, что крысы, выработавшие реакцию избегания крика боли без предварительного знакомства с болевым раздражением, характеризуются высоким уровнем исследовательской активности в «открытом поле», отсутствием дефекаций и уринаций в ситуации «открытого поля» (показателя пассивно-оборонительной реакции страха), а также низкой агрессивностью при раздражении лап двух крыс электрическим током. Крысы, не обнаружившие способности к выработке условной реакции избегания, напротив, оказались малоактивными в «открытом поле», трусливыми (если судить по дефекациям и уринациям) и агрессивными. Сочетание этих трех качеств наиболее неблагоприятно для выработки условного рефлекса.
В опытах, поставленных Д. З. Партев, у каждой из 10 крыс отдельно вырабатывали условный оборонительный рефлекс на звонок, подкрепляемый болевым раздражением лап. Камера с решетчатым полом была разделена на две половины перегородкой, имевшей дверцу Электрический ток подавали на решетку пола одной половины камеры после 5–8 с изолированного действия звонка, который продолжался еще 10 с вместе с болевым раздражением.
После того как у всех 10 крыс был выработан стабильный оборонительный условный рефлекс (80-100 % реакций на условный сигнал), в камеру помещали одновременно от 3 до 5 животных. Если теперь включить звонок, то крысы, ранее быстро перебегавшие на безопасную половину камеры, будут вести себя по-разному. Одни из них по-прежнему устремляются на безопасную половину, в то время как другие начинают драться. Некоторые из крыс, перебежав на безопасную половину, встают в дверях и атакуют крыс, пытающихся спастись от тока. Подчеркнем, что убегают или дерутся одни и те же животные. Даже в том случае, если условный сигнал (звонок) застает агрессивную крысу рядом с открытой дверцей, крыса не убегает от тока, а направляется в прямо противоположном направлении, чтобы атаковать другое животное. При выработке условной реакции избегания крика боли (до начала экспериментов с групповым поведением) у крыс, находившихся в «домике» в среднем 2,13 мин, в групповом эксперименте после 120 применений условного сигнала (звонка) в 80 % случаев зарегистрирован уход на безопасную половину и только в 14 % —драки. У тех же крыс, которые находились в «домике» в среднем 4,03 мин, количество драк достигло 65 %, а уход от раздражения током наблюдался лишь в 46 % случаев. Что касается «защиты безопасной территории», то эта форма поведения наблюдалась у обеих категорий крыс в одинаковом числе случаев (6 %). Опыты Д. P. Партев показали, что характер реакции на болевое раздражение партнера представляет устойчивую черту индивидуальных особенностей данного животного, закономерно коррелирующую с его поведением в ситуации угрозы собственного болевого раздражения, причем крысы малочувствительные к крику боли «жертвы» более склонны к агрессивному поведению.
Можно ли изменить реакцию крыс на крик боли другой особи и в каких пределах?
182 крысы, находившиеся на педали свыше 1 мин 30 с, мы от 3 до 10 раз использовали в качестве «жертв» при выработке условной реакции избегания у их партнеров. Эта процедура привела к тому, что у 14 животных время пребывания в «домике» резко сократилось, хотя у 68 оно осталось без изменений (см. крыс № 11,12 на рис. 8). Можно было предположить, что применение тока ведет к выработке обычного оборонительного рефлекса, где ограниченное пространство становится условным сигналом болевого раздражения. Экспериментальные факты противоречат подобному объяснению. Как только начинается угашение условного рефлекса (то есть крысу-«жертву» перестают раздражать током), крысы уже в первой пробе находятся на педали свыше 4 мин. Значит, использование подопытного животного в качестве «жертвы» ведет не к выработке обычного оборонительного условного рефлекса, а повышает чувствительность к сигналам оборонительного возбуждения другой особи. Аналогичные результаты ранее получил Р. Чёрч [Church, 1959].
Впрочем, процедура воздействия электрическим током дает этот эффект только у 42 % от общего количества исследованных животных. 68 из 267 крыс (или 26 %) продолжали находиться на педали свыше 3 мин (в среднем 3,77 мин) и после ознакомления с током. Возник вопрос: не объясняется ли эта «нечувствительность» к крику боли другой особи затруднением в выработке условной реакции избегания, то есть дефектом механизмов научения, приобретения новых навыков? Известные основания к такому предположению у нас возникли во время опытов Д. 3. Партев, где выяснилось, что крысы, находившиеся на педали меньше 2 мин, реагируют на сигнал раздражения током условным оборонительным рефлексом в 81 % всех применений этого сигнала на протяжении 11–14 дней, а крысы, находившиеся на педали свыше 3 мин, — только в 61 % случаев. Вот почему нас заинтересовало предложение д-ра В. Вецель применить оротовую кислоту — вещество, способствующее консолидации вновь выработанных условных связей.
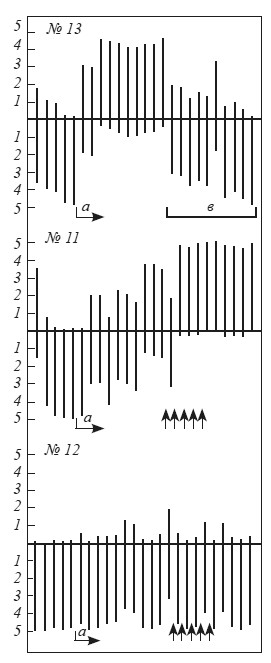
Рис. 8. Динамика выработки реакций избегания у крыс № 11, 12, 13
Абсцисса — последовательные пробы. Ниже оси ординат — время пребывания крысы в «домике» (мин), выше — время пребывания вне «домика»; а — начало выработки условного рефлекса; b — угашение. Стрелки обозначают использование данной крысы в качестве «жертвы».
Опыты с оротовой кислотой были поставлены совместно с д-ром В. Вецель на 10 крысах. В 30 мл воды растворяли 300 мг оротовой кислоты и 375 мг метилглюкамина и вводили этот раствор внутрибрюшинно из расчета 1 мл на 100 г веса животного. Контрольным 10 крысам вводили 0,9 %-ный раствор хлористого натрия также 1 мл на 100 г веса. Инъекции растворов производили за один час до начала опытов с выработкой условной реакции избегания.
У всех крыс на протяжении первых 5 дней вырабатывали условную реакцию избегания крика боли. В течение следующих 5 дней в отсек для «жертвы» помещали двух крыс, поскольку ранее нами было показано, что борьба крыс, спровоцированная болевым раздражением лап, является более сильным стимулом по сравнению с криком боли одной крысы. С 11-го по 15-й день производили угашение выработанной условной реакции.
Статистическая обработка результатов показала неправомерность объединения всех подопытных и всех контрольных крыс только в две группы. Достоверные различия можно получить, лишь разделив каждую группу на две подгруппы животных, чувствительных и нечувствительных к крику боли партнера. В этом случае разница между крысами, которым вводили оротовую кислоту, и контрольными животными оказалась достоверной у крыс, реагирующих на крик боли при раздражении одной «жертвы». Усиление стимула путем одновременного раздражения двух «жертв» маскирует эффект оротовой кислоты, который снова становится статистически значимым в опытах с угашением. Утешение условной реакции избегания происходило достоверно медленнее у крыс, получавших оротовую кислоту и обнаруживших ранее достаточно высокую реактивность к крику боли партнера.
Таким образом, мы убедились, что введение оротовой кислоты способствует выработке и закреплению условной реакции избегания только у тех животных, для которых крик боли партнера является достаточно эффективным стимулом. Ни усиление сигналов оборонительного возбуждения путем использования двух «жертв», ни применение болевого раздражения током, ни химическое воздействие оротовой кислотой не влияют на крыс, исходно не реагирующих или слабо реагирующих на крик боли другой особи.
Совместно с М. Л. Пигаревой и Ф. А. Бразовской мы исследовали изменение реакций избегания крика боли у крыс после повреждения различных образований головного мозга. Для повреждения фронтальной и цингулярной областей коры удаляли соответствующую вышележащую пластинку черепной кости и производили термокоагуляцию мозговой ткани электродом из нихромовой проволоки. Коагуляцию энторинальной области коры и подкорковых образований осуществляли с помощью стереотаксически введенных стальных электродов, изолированных лаком за исключением кончика диаметром 0,1–0,2 мм (ток 2,5–3 мА в течение 20–30 с). Индифферентный электрод прикрепляли к хвосту животного. Стереотаксические координаты определяли по атласу Е. Фифковой и Дж. Маршала. К опытам приступали через 10–12 дней после операции. По окончании опытов головной мозг подвергали гистологическому контролю (окраска по методу Ниссля). Каждый 20-й срез толщиной 20–40 мкм окрашивали крезилвиолетом.
Все исследованные нами отделы головного мозга можно разделить на три основные категории:
1) мозговые образования, повреждение которых не оказывает достоверного влияния на реакцию избегания у крыс — цингулярная и энторинальная кора, гиппокамп, перегородка и маммилярные тела;
2) структуры, эффект повреждения которых отчетлив и вместе с тем не зависит от индивидуальных особенностей животного — фронтальная область коры;
3) структуры, последствия повреждения которых определяются индивидуальными особенностями животного до операции — миндалина, центральное серое вещество и гипоталамус.
На рис. 9 представлены последствия повреждения фронтальной области коры, центрального серого вещества и миндалины у 30 крыс. Каждая группа из 10 крыс разделена на две подгруппы I и II в зависимости от чувствительности к крику боли партнера до операции. Удаление фронтальной коры уменьшило время пребывания на педали и в первой и во второй подгруппе животных практически одинаково (44 и 41 % соответственно). После разрушения центрального серого вещества время пребывания на педали резко возросло у животных, высокочувствительных к крику боли (325 %). У крыс, находившихся на педали в среднем 2,65 мин, оно также возросло, но только на 121 %. Двустороннее повреждение миндалин сократило время пребывания на педали почти вдвое (53 %) у крыс первой подгруппы и практически не изменило его во второй подгруппе (106 %).
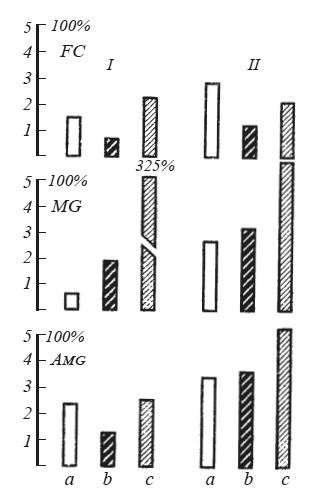
Рис. 9. Среднее время (мин) пребывания на педали двух (/ и //) групп крыс при болевом раздражении другой особи а — до операции; b — после операции; с — изменение времени пребывания на педали в процентах; FC — удаление фронтальной коры; MG — разрушение центрального серого вещества; Amg — повреждение миндалины.
Эффект, противоположный последствиям амигдалоэктомии, наблюдался после повреждения латерального гипоталамуса. Время пребывания на педали сократилось у крыс, ранее находившихся в «домике» сравнительно долго, и возросло у тех животных, которые большую часть пятиминутной экспозиции проводили в открытом пространстве. Иными словами, после повреждения латерального гипоталамуса все крысы становятся похожими друг на друга и проводят в «домике» и вне «домика» примерно одинаковое время. Для этих крыс характерно застревание в дверях «домика», когда одна половина тела животного находится на педали, а другая — в открытом пространстве.
Эксперименты с локальным повреждением структур головного мозга позволяют сделать следующие выводы:
Условная реакция избегания крика боли зависит от сохранности не одного какого-либо отдела мозга, но целого ряда мозговых образований: фронтальной коры, центрального серого вещества, миндалины и гипоталамуса.
Вместе с тем неправомерно утверждать, что условная реакция избегания осуществляется «мозгом как целым», потому что имеется ряд структур, повреждение которых заметно не сказывается на изучавшейся форме поведения. К их числу относятся, например, энторинальная и цингулярная кора, гиппокамп, перегородка и маммилярные тела.
Участие мозговых образований в изучавшейся реакции тесно связано с функциональной специализацией этих образований, которая обнаруживается при исследовании других форм поведения: условнорефлекторного переключения пищевых и оборонительных рефлексов по Э. А. Асратяну, при выработке условных рефлексов с частичным (вероятностным) подкреплением и т. д.
Особый интерес представляет наличие структур (центральное серое вещество, миндалина и гипоталамус), эффект повреждения которых зависит от индивидуальных особенностей животного, выявленных до оперативного вмешательства. Можно предполагать, что именно эти структуры представляют нейроанатомический субстрат индивидуально различной реактивности крыс по отношению к сигналам оборонительного возбуждения партнера, равно как и ряда других форм поведения, присущих данному животному.
Среди мозговых образований последней группы особенно демонстративные и однозначные результаты мы получили при разрушении центрального серого вещества. После повреждения этого отдела мозга все крысы, ранее хорошо реагировавшие на крик боли партнера, резко ухудшили реакцию избегания. По данным ряда исследователей, центральное серое вещество связано с проведением болевой афферентации и с интеграцией реакций на эмоционально отрицательные (аверсивные) стимулы: звуки, запахи и т. д. Можно предположить, что в процессе эволюции естественный отбор «использовал» для реагирования на сигналы отрицательного эмоционального состояния другой особи те же мозговые механизмы, которые связаны с восприятием аверсивных, в том числе — болевых стимулов, адресованных самому субъекту. Эти данные, полученные в опытах на животных, помогают понять механизм того явления, когда человек, став свидетелем страданий другого, испытывает почти физическое недомогание: сжатие в области сердца, «комок в горле», тошноту и т. п. соматические симптомы.
Л. А. Преображенская [1973] вырабатывала инструментальный условный двигательный рефлекс избегания у собак при болевом раздражении партнера. Две собаки помещались в одной камере на разных станках, расположенных на расстоянии полуметра один от другого. На заднюю лапу собаки-«жертвы» подавали ток значительной силы, вызывавший не только локальный подъем лапы, но и одышку, скуление, часто визг. У второй собаки — «наблюдателя» левая передняя лапа прикреплялась к рычагу. Подъемом лапы выше определенного уровня животное могло отключить действие тока на первую собаку. Звуковой сигнал (тон 600 Гц) на 10 с опережал действие тока, которое продолжалось 30 с. В случае, если собака поднимала лапу и отключала ток, ток прекращался. У собак регистрировали двигательные реакции, дыхание, электрокардиограмму и электрограмму дорзального гиппокампа.
Условный рефлекс удалось выработать у трех из шести подопытных животных (рис. 10). Заметим, что ранее выработанный «собственный» оборонительный рефлекс не способствует реакции избегания при болевом раздражении партнера, если чувствительность данного животного к сигналам оборонительного возбуждения другой особи низка. На рис. 11 видно, что при первых применениях условного сигнала после того, как электроды перенесли с лапы «наблюдателя» на лапу «жертвы», еще наблюдается короткий условнорефлекторный подъем рычага, отключающего ток. Однако вскоре «наблюдатель» перестает реагировать и на условный сигнал, и на сигналы, исходящие от партнера.
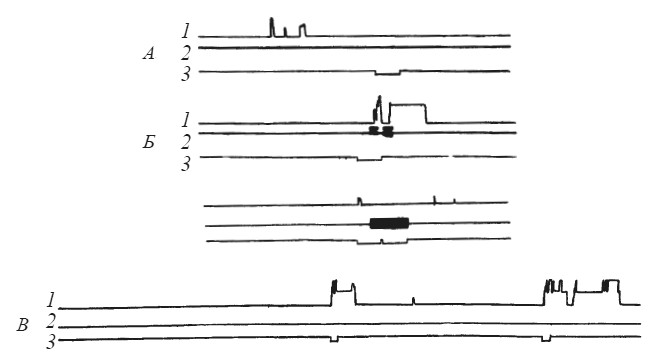
Рис. 10. Динамика выработки реакции избегания у собаки при болевом раздражении партнера
1 — подъем лапы собакой-«наблюдателем»; 2 — действие тока на собаку-«жертву»; 3 — отметка условного сигнала (по Л. А. Преображенской); А, Б, В — последовательные фрагменты опыта
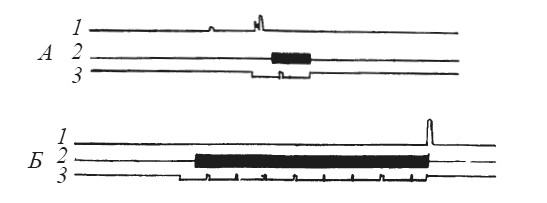
Рис. 11. Исчезновение условной реакции избегания после переноса электродов на собаку-«жертву»
Обозначения те же, что на рис. 10 (по Л. А. Преображенской)
У собак, чувствительных к болевому раздражению партнера, можно зарегистрировать объективные признаки эмоционального напряжения в виде учащения сердцебиений и нарастания тета-ритма в гиппокампе (рис. 12, табл. 2).
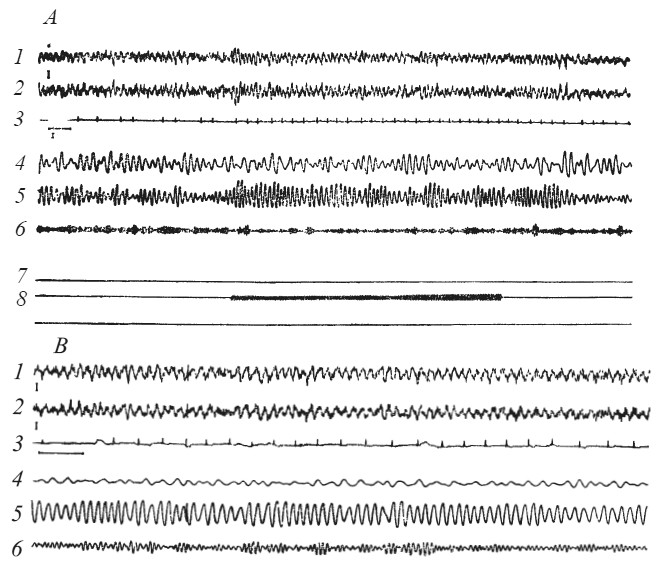
Рис. 12. Изменения ЭГ гиппокампа и ЭГК при болевом раздражении собаки (В) или ее партнера (Д)
1, 2 — электрограмма гиппокампа; 3 — ЭКГ; 4, 5, 6 — дельта-, тета- и альфа-частоты ЭГ, выделенные анализатором; 7 — отметка подъема лапы собакой-«наблюдателем»; 8 — действие тока на собаку-«жертву» (по Л. А. Преображенской).
Статистическая обработка полученных данных показала, что число опытов, в которых наблюдаемые изменения были достоверны, колебалось у разных животных от 66 до 87 %. Эти сдвиги носили весьма кратковременный характер и после прекращения раздражения партнера быстро возвращались к исходным значениям. Поведение собаки-«наблюдателя» в интервалах между применением тока было спокойным, некоторые из них дремали. У двух собак были выработаны два инструментальных условных рефлекса: подъем левой передней лапы прекращал болевое раздражение партнера, а нажим правой лапой на педаль приводил к подаче пищи «наблюдателю». Для определения соотносительной силы двух мотиваций проводили специальные пробы с одновременным раздражением партнера и включением пищевого условного сигнала. Оказалось, что в большинстве случаев собака осуществляет реакцию, прекращающую оборонительное возбуждение партнера. Аналогичные результаты получены в опытах на обезьянах: из 15 макак-резусов 10 переставали нажимать на рычаг, если подача кормушки сопровождалась болевым раздражением другой особи [Masserman, Wechkin, Terris, 1964].
Таблица 2. Средняя величина частоты сердечных сокращений и амплитуды интегрированного тета-ритма (в усл. ед.) у собак-«наблюдателей» (по Л. А. Преображенской, 1973)
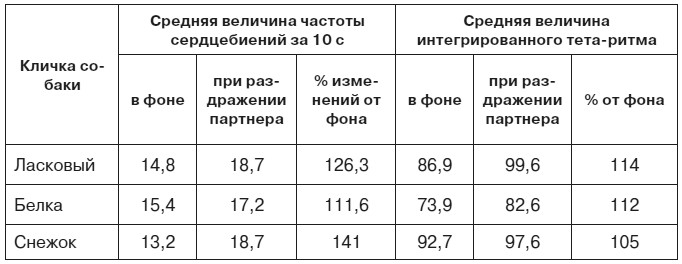
Таким образом, сигналы оборонительного возбуждения другой особи вызывают участи животных-«наблюдателей» состояние отрицательного эмоционального напряжения, которое животные стремятся минимизировать, то есть прервать или предотвратить. Наши собственные эксперименты на крысах и собаках, равно как и результаты других исследователей, в том числе в опытах на обезьянах, показали, что индивидуальные особенности реагирования на крик боли преимущественно зависят от потребностно-мотивационных особенностей данного животного, а не от его способности к обучению. Вот почему среди животных, стоящих на разных ступенях эволюционного развития (крыса, собака, обезьяна), обнаруживаются особи, в разной мере чувствительные к сигналам эмоционального состояния другого существа того же вида.
Качественно новый и более сложный характер реакции на эмоциональное состояние другого приобретают у человека. Трудно согласиться с представлением о сопереживании как простой имитации, прямом воспроизведении эмоций по принципу: встретил радостного человека и обрадовался, увидел печальное лицо — почувствовал грусть. А ведь подобную точку зрения нередко распространяют даже на искусство, рассматривая его в качестве средства «передачи чувств» от художника к зрителю. Экспрессия чувств действует при этом «заражающим» образом, а зритель отвечает актеру «сопереживающим пониманием», в результате чего происходит «эмоциональное понимание происходящего» [Якобсон, 1977].
Но именно искусство, особенно искусство театра, ежедневно опровергает версию о сопереживании как имитации, как следствии пресловутой «заразительности чувств». «Бывает, что плачет актриса, что называется “живыми слезами”, — писал народный артист СССР Н. Плотников, — как и требовал того режиссер на репетициях. Все верно, а зритель толкает товарища в бок, бинокль дает: “Посмотри-ка, плачет! По-настоящему!” Вроде бы, правда жизни на сцене (плачет-то актриса взаправду), а искусства нет» [Плотников, 1971]. Может быть, только в раннем детстве мы встречаемся с механизмом чисто имитационного воспроизведения эмоций группой детей без уяснения причин, почему заплакал этот первый мальчик или почему заливается смехом девочка, вслед за которой захохотала вся группа? Позднее механизмы сопереживания становятся все более сложными.
Для того чтобы сострадать, недостаточно просто быть свидетелем страдания другого человека, как полагают сторонники непосредственной «заразительности» эмоций. Присущие детям в определенном возрасте бесчувственность и жестокость имеют в своей основе неспособность ребенка связать внешние признаки состояния другого с аналогичным собственным состоянием и характерной для него экспрессией. Как мы могли убедиться выше, даже у животных собственный опыт знакомства с болевым воздействием существенно влияет на их чувствительность к сигналам оборонительного возбуждения другой особи. Сочувствию и сопереживанию надо учиться, а развитие этой способности в решающей мере зависит от правильного воспитания. «Труд души, — писал В. А. Сухомлинский, — это значит страдать, болеть страданиями и болями человека — прежде всего матери, отца, сестры, дедушки, бабушки. Не бойтесь открывать юную душу для этих страданий — они благодарны. Пусть девятилетний сын ночь не спит у постели заболевшей матери или отца, пусть чужая боль заполнит все уголки его сердца. Одна из самых мучительно трудных вещей в педагогике — это учить ребенка труду любви» [Сухомлинский, 1971, с. 4]. И в другом месте: «Искусство облагораживания ребенка и подростка высшими чувствами и переживаниями является искусством сопереживания» [Сухомлинский, 1979, с. 273].
Сколь ни важна способность сочувствия (переживания эмоций, сходных с эмоциями другого), сама по себе она еще не определяет характера действий. Л. Н. Толстой и Ф. М. Достоевский отличали любовь пассивную, созерцательную и потому бесплодную от любви активной, действенной. Здесь мы снова встречаемся с правилом, сформулированным информационной теорией эмоций. Сигналы состояния другого человека есть лишь информация, воспринимаемая субъектом. Что же касается действий этого субъекта, то они будут зависеть от потребности, доминирующей в структуре данной личности. Ею может оказаться и родительская потребность помочь своему (или не своему) ребенку, и сложная социальная потребность соответствовать этическому эталону, которая предпишет вести себя строго определенным образом, и эгоистическая потребность остаться в стороне, избавить себя от лишних хлопот. Каждая из этих потребностей вызовет к жизни свой ряд эмоций и свою цепь действий. «Любовь мечтательная, — писал Достоевский, — жаждет подвига скорого, быстро удовлетворимого и чтобы все на него глядели. Тут действительно доходит до того, что даже и жизнь отдают, только бы не продлилось долго, а поскорей совершилось, как бы на сцене, и чтобы все глядели и хвалили. Любовь же деятельная — это работа и выдержка, а для иных так, пожалуй, целая наука» [Достоевский, 1958, с. 75–76]. Спустя много лет об этом же написал Эрих Фромм: «Продуктивная любовь несовместима с пассивностью, с тем чтобы просто созерцать жизнь любимого существа: она включает деятельность, заботу и ответственность за его развитие» [Fromm, 1960, р. 100–101].
Задача воспитания в сущности и состоит в формировании наиболее ценных для данного общества потребностей (мотивов, побуждений). «Труд души» в смысле В. А. Сухомлинского не сводится к тому, чтобы ребенок просто ощущал некий эмоциональный дискомфорт при виде больного или несправедливо обиженного человека, не просто стремился к устранению этого тягостного для него самого «сопереживания», но приходил на помощь и переживал положительные эмоции, которые приносит успех действий, направленных на облегчение участи другого.
Таков самый краткий обзор регуляторных — переключающих, подкрепляющих, компенсаторных — функций эмоций на индивидуальном и популяционном уровнях. На всем протяжении главы мы стремились показать, что эти регуляторные функции эмоций непосредственно вытекают из их отражательно-оценочной функции, обусловлены ею. В отличие от концепций «отношения», «значимости», «смысла» и т. п. информационная теория эмоций точно и однозначно определяет ту объективно существующую реальность, тот «эталон» (если использовать выражение В. К. Вилюнаса), который получает субъективное отражение в эмоциях человека и высших животных: потребность и вероятность (возможность) ее удовлетворения. Именно эти два фактора делают события «значимыми» для субъекта, придают им «личностный смысл» и побуждают субъекта не только переживать, но и выражать, действенно реализовать свое «отношение» к окружающему миру и к самому себе.
Назад: Подражательное поведение как пример компенсаторной функции эмоций на популяционном уровне
Дальше: Анализ критических замечаний в адрес информационной теории эмоций

