Книга: Эмоциональный мозг
Назад: Нейроанатомия эмоции
Дальше: Значение передних отделов новой коры для ориентации поведения на сигналы высоковероятных событий
Морфофизиологические основы потребностей, мотиваций и эмоций. их относительная самостоятельность
На схеме фронтального среза мозга крысы в одной из работ Дж. Олдса (рис. 24) структуры, связанные с биологическими потребностями в пище, воде и сексуальном партнере, выглядят как островки на фоне мозговой ткани, прямое раздражение которой дает эмоционально положительные, эмоционально отрицательные и смешанные эффекты. Раздражение мозга кошки в зоне от заднего ядра до супраоптического в латеральной части гипоталамуса ниже нитевидного ядра провоцирует нападение на крысу без внешних проявлений ярости («холодная» атака). Нападение с яростью наблюдается при смещении электрода в медиальном направлении. Ярость без нападения — при стимуляции свода в точке, лежащей выше нитевидного ядра [Wasman, Flynn 1962; Flynn, 1967].
При одной и той же локализации электродов в латеральном гипоталамусе крыс порог активизации мотивационных структур, о котором судили по влиянию на процессы еды, питья, копуляции, оказался значительно ниже порога эмоционального позитивного подкрепления — самостимуляции [Huston, 1971,1972]. Раздражение латерального гипоталамуса длинными пачками стимулов приводило к большему потреблению молока, чем раздражение короткими пачками, однако крысы предпочитали последний тип раздражения. Следовательно, мотивационные и подкрепляющие (эмоционально положительные) системы разделены в мозге, хотя при самостимуляции наблюдается их одновременное возбуждение [Ball, 1969]. Фармакологический анализ также свидетельствует о различных механизмах потребностей и эмоций. Введение дисульфирама, разно снижающее уровень норадреналина в мозге крыс, устраняет мотивационные эффекты стимуляции (еда, питье, грызение), повышая частоту самораздражений и снижая порог самостимуляции [Михайлова, Черешаров, 1979]. По данным С. А. Борисенко [1977], фенамин и кокаин облегчают самораздражение гипоталамуса и перегородки, одновременно подавляя пищевые и питьевые реакции животного.
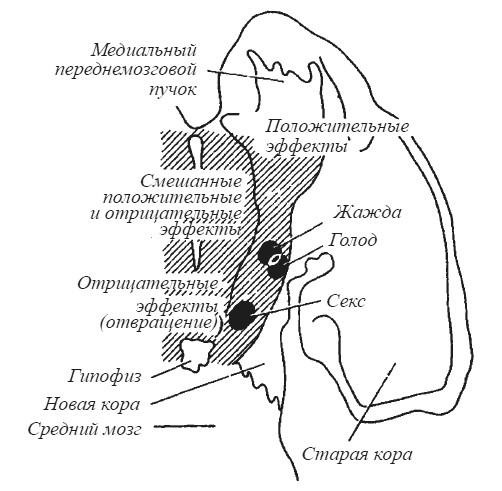
Рис. 24. Схема мозга крысы с обозначением зон, электрическое раздражение которых вызывает мотивационные и эмоциональные эффекты (по Дж. Олдсу)
До сих пор, говоря об анатомическом субстрате потребностей, мы имели в виду актуализацию натуральных потребностей животного в пище, воде и т. п. Что касается феномена самораздражения, то механизм, побуждающий животное вновь и вновь нажимать на педаль, связан, по- видимому не с голодом, жаждой и т. д., но со следовым возбуждением структур эмоционально положительного подкрепления, активированных в момент действия тока. Непродолжительность этого следового возбуждения объясняет и сравнительную легкость угашения реакций при их неподкреплении, и необходимость «затравочных» стимуляций у животных, обученных нажимать на педаль [Deutsh, Howarth, 1963]. Еще в первых своих работах Дж. Олдс сравнивал феноменологию самораздражения не с голодом, а со стремлением к вкусной пище. Крысы могут стимулировать структуры голода и жажды, но только при наличии пищи и воды в экспериментальной обстановке, когда высокая вероятность удовлетворения натуральных потребностей гарантирует возникновение положительных эмоций в процессе еды и питья [Morgan, 1969; Mendelson, 1970]. Цитава обоснованно предлагает различать систему «побуждения» типа голода, жажды, боли и систему «желания» как стремления к повторной активизации структур положительных эмоций. Активность системы «желания» не тождественна механизму самого эмоционально положительного возбуждения («удовольствия»), возникающего в момент подкрепления. По мнению Цитавы, в системе «желания» преобладают допаминергические структуры, а в системе «удовольствия» — норадренергические [Cytawa, 1979]. Что касается прерывания стимуляции, то в зависимости от локализации электрода и параметров тока она может быть обусловлена как привыканием к действию тока, так и вовлечением механизмов отрицательных эмоций [Звартау, Паткина, 1972; Григорьян, 1978].
Мы полагаем, что в пользу относительной самостоятельности механизмов потребностей, мотиваций и эмоций свидетельствуют опыты, поставленные в нашей лаборатории Н. Г. Михайловой и К. Ю. Саркисовой. В своих экспериментах авторы использовали прием градуального нарастания интенсивности стимулирующего тока, давно применяющийся А. В. Вальдманом, М. М. Козловской, Дж. П. Хьюстоном и другими исследователями.
22 белым крысам-самцам были вживлены монополярные стальные электроды с диаметром кончика 70 мкм в латеральную преоптическую область и в латеральный гипоталамус (координаты по атласу Кенига и Клиппель: А — 7,0; L — 1,8; Н — 3,5; А — 3,0–3,5; L — 1,5; Н — 3,5). Во время стимуляций в камере находилась педаль для самораздражения, а также целевые объекты для выявления специфических мотиваций: пищевой (семечки или овес, равномерно рассыпанные на полу), питьевой (поилка с водой), грызения (кусок мела или дерева), сексуальной (самка). Раздражение производили как ритмическим током (прямоугольные монофазные импульсы частотой 100 Гц, длительностью 0,1 мс, интенсивностью от 0 до 1,0 мА), так и постоянным током силой от 0 до 70 мкА. Консуматорные реакции (еда, питье, грызение) регистрировали с помощью записи миограммы жевательных мышц. Вычисляли латентные периоды реакций и вероятность их проявления, то есть отношение числа стимуляций, вызвавших эти реакции, к общему числу раздражений. Подкрепляющий (эмоционально положительный) эффект действия тока оценивали по частоте самораздражений. Перед опытом крысы имели свободный доступ к пище и воде.
Во всех случаях производили стимуляцию только тех зон, которые при воздействии током достаточной силы способны дать реакцию самораздражения. Оказалось, что при поляризации этих зон градуально нарастающим постоянным током также, как и при стимуляции их ритмическим током возрастающей интенсивности, поведенческие реакции всегда имеют одну и ту же последовательность. Слабое раздражение вызывает генерализованную поисковую активность без обращения к находящимся в камере целевым объектам — к пище, воде, особи другого пола и т. д. Только при усилении интенсивности раздражения эти внешние стимулы становятся эффективными: животное начинает есть, иногда пить, грызть и т. п. При дальнейшем усилении ритмического или постоянного тока возникает реакция самораздражения.
Если подвергать стимуляции два различных пункта в гипоталамусе, то наблюдается аналогичная последовательность событий, то есть два раздражения «поисковой интенсивности» дают консуматорные реакции: чаще всего — еды, реже — питья, грызения и т. д., а два раздражения «мотивирующей интенсивности» способны вызвать реакцию самораздражения (рис. 25). Воздействие на один из пунктов, достаточное для того, чтобы вызвать самораздражение, подавляет мотивированное поведение, обусловленное стимуляцией второго пункта. Уайт [White, 1973] наблюдал прекращение натурального пищевого поведения при раздражении миндалины, способном вызвать самостимуляцию. В опытах Н. Г. Михайловой и К. Ю. Саркисовой [1977] выявилось методическое преимущество использования постоянного тока по сравнению с ритмическим. Стимуляция двух пунктов ритмическим током ведет к усилению (учащению) реакций самораздражения. Слабая катодная поляризация одного пункта тормозит самораздражение второго очага, а повышение интенсивности постоянного тока — усиливает самостимуляцию. Отметим, что тормозящее влияние оказывает та интенсивность постоянного тока, которая при изолированном применении вызывает целенаправленное мотивированное поведение.

Рис. 25. Последствия одновременной стимуляции двух пунктов гипоталамуса крысы электрическим током «поисковой» (А) и «мотивирующей» (Б) интенсивности
1 — миограмма жевательных мышц; 2, 3 — стимуляция первого и второго пункта. Отклонение линии 2 на фрагменте в регистрирует самораздражение (по Н. Г. Михайловой и К. Ю. Саркисовой).
Наблюдавшуюся трансформацию эффектов трудно объяснить неспецифической дополнительной активацией «мотивационно-подкрепляющих» структур потому, что стимуляция эмоциально негативных (вызывающих избегание) зон ретикулярной формации среднего мозга оказывает тормозящее влияние на реакцию самораздражения (эффект, ранее подробно исследованный Н. Г. Михайловой [1975]), а стимуляция эмоционально нейтральных зон гипоталамуса вообще не влияет на самораздражение. Только возбуждение эмоционально позитивных структур ретикулярной формации, способных самостоятельно обусловить хотя бы слабое самораздражение, суммируется с «поисковым», «мотивирующим» и «подкрепляющим» возбуждением структур гипоталамуса.
Мы думаем, что модельные эксперименты, о которых речь шла выше, воспроизводят последовательность событий, характерную для организации натурального поведения (рис. 26). Слабое раздражение электрическим током активирует систему мозговых структур, представляющую субстрат потребности. Процесс актуализации потребности, еще не трансформировавшейся в мотивацию, внешне проявляется в виде генерализованного поискового беспокойства. Только усиление возбуждения при нарастании интенсивности постоянного или ритмического тока ведет к активации тех структур, которые хранят энграммы внешних объектов, способных удовлетворить данную потребность. В результате внешние стимулы становятся эффективными, и мотивированное животное начинает есть, пить, грызть и т. п. Однако необходимо дальнейшее усиление тока, чтобы произошла активация структур эмоционально положительного подкрепления и животное перешло к самораздражению мозга в отсутствие естественного удовлетворения какой-либо потребности.
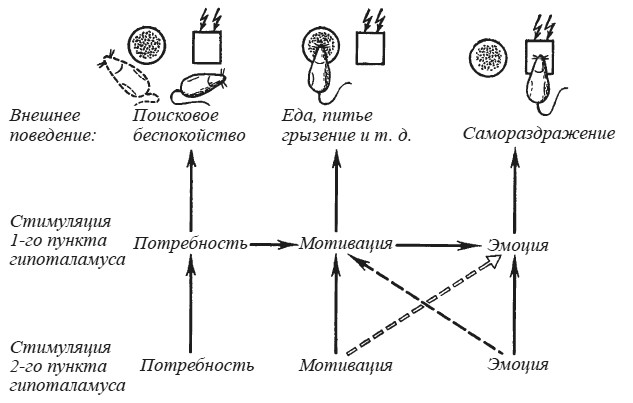
Рис. 26. Схема последствий одновременной стимуляции двух пунктов гипоталамуса
Сплошные стрелки — суммирующиеся эффекты; прерывистые стрелки — тормозящие влияния стимуляции ритмическим и постоянным током (тонкая стрелка) и только постоянным (двойная стрелка) током.
Разумеется, мы не представляем себе дело таким образом, будто структуры потребности, мотивации и эмоции линейно располагаются в соседних участках мозговой ткани и электрический ток нарастающей интенсивности последовательно захватывает эти структуры одну за другой. Речь идет о том, что система структур, необходимых и достаточных для актуализации потребности, более проста, содержит в себе меньше элементов, чем система, обеспечивающая целенаправленное мотивирование животного. А полный комплекс морфофизиологической организации поведения (потребность + мотивация + эмоция) предполагает дополнительное вовлечение еще и нервного аппарата эмоций.
Для того чтобы понять последствия стимуляции двух пунктов гипоталамуса электрическим током различной интенсивности, целесообразно напомнить данные, полученные при исследовании механизмов естественного пищевого поведения К. В. Шулейкиной, Дж. Олдсом и другими авторами.
Как показала К. В. Шулейкина [1971], голодовое возбуждение (актуализация потребности в пище) первично возникает в ретикулярных ядрах моста и продолговатого мозга, охватывает центральное серое вещество среднего мозга, неспецифические ядра таламуса и дорзальный гиппокамп. Для того чтобы актуализированная пищевая потребность трансформировалась в целенаправленный поиск пищи, необходимо вовлечение структур новой коры, миндалины и «положительных элементов» (терминология К. В. Шулейкиной) медиального гипоталамуса.
Заметим, что структуры гипоталамуса начинают активно функционировать на стадии пищедобывательного поиска, инициированного голодовым возбуждением. В латеральном гипоталамусе голодных обезьян можно найти нейроны, которые отвечают изменением своей активности на вид и запах пищи, на условные сигналы предстоящей еды [Mora, Rolls, Burton, 1976]. Там же в латеральном гипоталамусе обнаруживаются нейроны, которые активируются и другими потребностями: жаждой или состоянием морфийной абстиненции у крыс [Олдс, 1977]. Но вот животное входит в контакт с пищей и начинает акт еды, реализуемый синхронизирующей системой нижнего отдела мозгового ствола, таламусом, орбитальной корой и латеральным гипоталамусом. Из полости рта, а затем из желудка поступают нервные импульсы, сигнализирующие о предстоящем утолении голода. Давно известно, что подобная афферентация прекращает акт еды задолго до истинного «тканевого» насыщения. Этот процесс, позднее получивший название «сенсорного насыщения», ярко описан в 1910–1911 гг. И. П. Павловым: «Попадание небольшого количества пищи в желудок временно прекращает или ослабляет действие пищевого центра… Положение дела могло бы быть хуже, если бы раздражимость пищевого центра падала только при полном удовлетворении потребности организма в жидких и твердых питательных веществах, так как постоянным следствием такого условия было бы переедание, чрезмерное переполнение желудка пищей» [Павлов, 1973, с. 108].
В самом начале еды поступление пищи еще продолжает активировать «положительные» нейроны медиального гипоталамуса, однако по мере насыщения происходит активация «отрицательных» нейронов, которые оказывают тормозящее влияние на нейроны латерального гипоталамуса, ранее возбужденные голодом. Прием пищи прекращается.
Мы солидарны с К. В. Шулейкиной в мнении о том, что заключение о локализации «центра голода» в латеральном гипоталамусе, а «центра насыщения» — в медиальном слишком упрощает реальное положение вещей. Мы склонны связывать структуры латерального гипоталамуса преимущественно с процессами пищевой мотивации на всех ее этапах от голодового возбуждения, обеспечивающего реакции нейронов латерального гипоталамуса на условные сигналы, вид и запах пищи, до состояния насыщения. Что касается «положительных» и «отрицательных» элементов медиального гипоталамуса, то изменения их активности дают основание отнести эти элементы к нервным механизмам эмоций — положительных в период подхода к пище и в начале акта еды и отрицательных, которые активируются по мере поступления пищи.
Вернемся теперь к анализу опытов с электрическим раздражением гипоталамуса. Слабое раздражение током, по-видимому, имитирует то голодовое возбуждение, которое в естественных условиях поступает в гипоталамус из ретикулярных ядер моста и продолговатого мозга. Это возбуждение должно быть усилено нарастающей интенсивностью электрического тока или стимуляцией второго «пищевого» (а не всякого, не любого) пункта, чтобы активировать энграммы пищевых объектов и обусловить акт еды. В процессе еды возбуждаются нервные элементы, генерирующие эмоционально положительное состояние, но это возбуждение опять-таки недостаточно для перехода к искусственному раздражению эмоционально позитивных структур электрическим током. Только дальнейшее усиление тока или суммация двух «мотивирующих» возбуждений ведет к замене еды самостимуляцией. Подчеркнем еще раз, что феномен искусственного подкрепления отнюдь не является следствием усиления мотивационного возбуждения. Суммируется не возбуждение мотивационных структур, а возбуждение эмоционально позитивных элементов, поскольку переход от еды к самораздражению происходит только при стимуляции тех пунктов, каждый из которых при достаточной силе тока способен вызвать реакцию самостимуляции.
Переход к самораздражению постепенно прекращает пищевое поведение крысы (соответствующее тормозящее влияние показано на рис. 26 тонкой прерывистой линией). Этот эффект лишний раз свидетельствует о том, что непосредственным подкрепляющим фактором инструментальных рефлексов является не удовлетворение какой-либо потребности, а максимизация положительного (или минимизация отрицательного) эмоционального состояния. Тормозящее влияние стимуляции мотивационных структур на реакцию самораздражения удалось выявить только при действии постоянного тока (на рис. 26 оно обозначено двойной прерывистой линией). Это угнетающее влияние с нашей точки зрения показывает, что в основе феномена самораздражения лежит именно активация структур положительных эмоций. В зависимости от степени голодового возбуждения одна и та же афферентация о предстоящем насыщении (или ее имитация путем раздражения электрическим током соответствующих мозговых структур) может оказаться либо достаточной, либо недостаточной для активации положительных эмоций. В последнем случае усиление мотивации ведет к возбуждению эмоционально отрицательных элементов и угнетает реакцию самораздражения.
Выше мы неоднократно подчеркивали положение о том, что любая эмоция реализуется не точечным «центром», а констелляцией, системой структур, расположенных на различных «этажах» головного мозга. Развивая теоретические концепции Шеррингтона, Магнуса, Павлова и обобщив собственные фактические данные, Э. А. Асратян [1959] сформулировал представление о множестве ветвей центральной части дуги безусловного рефлекса, каждая из которых проходит по различным отделам центральной нервной системы, включая кору большого мозга (рис. 27). Следующим шагом в развитии такого рода представлений будет, по-видимому, уточнение вопроса о специфическом «вкладе», вносимом тем или иным представительством данной рефлекторной системы в осуществление целостной биологически целесообразной реакции. Здесь возможны два варианта, схематически изображенные нами на рис. 28.
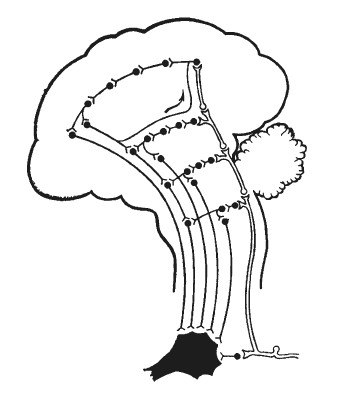
Рис. 27. Схема «многоэтажного» строения безусловного рефлекса по Э. А. Асратяну
Согласно первой точке зрения (рис. 28,I), каждая из потребностей (голод, жажда, секс и т. п.), равно как и каждая из эмоций (страх, ярость, удовольствие), имеют собственные «представительства» на различных этажах центральной нервной системы, в том числе в миндалине, гиппокампе, в новой коре больших полушарий. Согласно второй точке зрения (рис. 28, II), интеграция соматических и вегетативных компонентов, специфических для данной эмоции, обнаруживается только на определенном сравнительно низком уровне (в гипоталамусе?). Что же касается таких мозговых образований, как гиппокамп, миндалина и тем более новая кора, то в них представлены не отдельные потребности и эмоции, но операции, необходимые для генеза самых различных эмоциональных состояний.
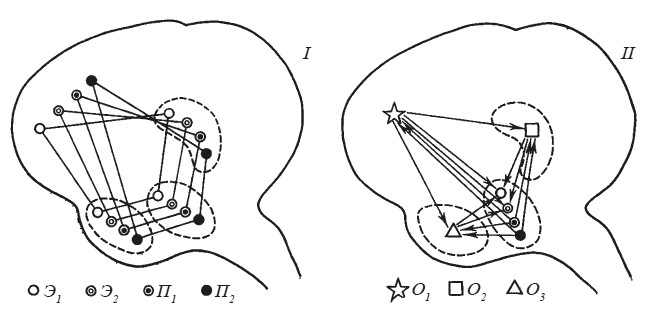
Рис. 28. Два возможных варианта функциональной организации мозговых механизмов потребностей и эмоций (I и II)
Э1 Э2 — эмоции; П1 Л2 — потребности; О1 О2 Оз — операции.
Суть этих операций определяется двумя факторами, имеющими решающее значение для организации любого поведения: наличием актуальных потребностей и возможностью их удовлетворения благодаря взаимодействию с внешней средой. Значимость стимулов, поступающих из внешней среды, зависит от их отношения к потребностям, имеющимся у организма, причем все эти стимулы можно разделить на две основные категории: на стимулы с высокой вероятностью их подкрепления факторами, непосредственно удовлетворяющими ту или иную потребность, и на стимулы с низкой вероятностью подкрепления. Среди актуальных потребностей, в свою очередь, выделяются наиболее острые доминирующие потребности, требующие первоочередного удовлетворения, и субдоминантные потребности, динамически сосуществующие с доминантной или конкурирующие с ней.
Экспериментальные данные, полученные в нашей лаборатории или почерпнутые из литературы, показали, что именно таким представлениям об организации поведения соответствует взаимодействие четырех мозговых образований, играющих решающую роль в оценке поступающих из внешней среды сигналов и выборе реакций. Мы имеем в виду передние отделы новой коры, гиппокамп, миндалину и гипоталамус.
Назад: Нейроанатомия эмоции
Дальше: Значение передних отделов новой коры для ориентации поведения на сигналы высоковероятных событий

