Рассказ Саламандры
Названия – вечный камень преткновения в науке об эволюции. Ни для кого не секрет, что палеонтология противоречива, и в этой отрасли науки нередко вспыхивает вражда. (Есть по крайней мере восемь книг, озаглавленных “Кости раздора”.) Чаще всего палеонтологи ссорятся из-за имен. Что это – ископаемый Homo erectus или архаичный H. sapiens? Ранний H. habilis или поздний Australopithecus? У каждого ученого всегда есть твердое мнение, но чаще всего оказывается, что спорят палеонтологи по пустякам. Это напоминает теологические споры – и неудивительно, что кипят страсти. Одержимость названиями – пример того, что я называю тиранией дискретного мышления.
Калифорнийская долина тянется вдоль почти всего штата Калифорния. На западе она ограничена Береговыми хребтами, на востоке – горами Сьерра-Невада. Эти цепи соединяются на севере и юге долины, так что последняя со всех сторон окружена горами. В этих горах живут саламандры рода Ensatina. Сама Калифорнийская долина (ее ширина достигает 80 км) для саламандр некомфортна, и они там не встречаются. Однако саламандры живут в окружающих долину горах, и их ареал образует вытянутое кольцо, более или менее непрерывное. Ножки саламандры за ее недолгую жизнь не могут унести животное далеко от места рождения. Но гены, которые живут гораздо дольше, – совсем иное дело. Саламандры могут скрещиваться со своими соседями, чьи родители могли скрещиваться с собственными соседями, и так далее. Поэтому потенциально по всему кольцу существует генетический перенос. А то, что происходит на самом деле, изучили мои коллеги из Калифорнийского университета в Беркли. Это исследование начал Роберт Стеббинс и продолжил Дэвид Уэйк.
В районе исследования, в горах к югу от долины (Camp Wolahi), обитает два вида Ensatina, которые не скрещиваются между собой. У одного из них четко выраженные желтые и черные пятна. Другой вид однородно светло-коричневый, без пятен. Район находится в зоне перекрывания ареалов этих видов, однако показано, что пятнистый вид типичен для восточной части Калифорнийской долины (этот район называется долиной Сан-Хоакин). Светло-коричневый вид обитает преимущественно в западной части долины Сан-Хоакин.
Нескрещиваемость – общепризнанный показатель того, заслуживают ли две популяции разных видовых названий. Поэтому логично использовать название Ensatma eschscholtzu для одноцветного западного вида и E. klauberi – для пятнистого восточного. Но есть любопытное обстоятельство.
Если подняться в горы, которые ограничивают Калифорнийскую долину с севера (там она называется долиной Сакраменто), мы найдем лишь один вид Ensatma. Он представляет собой нечто среднее между пятнистым и однотонным видами: в основном коричневый, с нечеткими пятнами. При этом он не является гибридным. Чтобы понять, в чем дело, нужно совершить две экспедиции на юг, собирая образцы саламандр в популяциях, образующих вилку на запад и на восток по обеим сторонам Калифорнийской долины. С восточной стороны саламандры становятся все более пятнистыми, пока не достигают крайней формы – E. klauben – далеко на юге. С западной стороны саламандры становятся все более похожими на одноцветных E. eschscholtzu, которые живут в зоне перекрывания.
Поэтому трудно с уверенностью считать E. eschscholtzu и E. klau-ben отдельными видами. Они образуют “кольцевой” вид. Отдельными видами их можно признать, лишь если собирать образцы на юге. Однако по мере движения на север они постепенно превращаются друг в друга. Зоологи обычно (вслед за Стеббинсом) объединяют их в один вид, E. eschscholtzu, который разделяют на ряд подвидов. На крайнем юге это E. eschscholtzu eschscholtzu (одноцветная коричневая форма). По мере движения по западной стороне долины мы встречаем E. eschscholtzu xanthoptica и E. eschscholtzu onegonensis, которые, как подсказывают их названия, также встречаются на севере, в Орегоне и Вашингтоне. В северной части Калифорнийской долины обитает полупятнистая E. eschscholtzu picta, о которой я упоминал. Двигаясь далее на юг по восточной стороне долины, мы встречаем E. eschscholtzu platensis, у которых чуть больше пятен, чем у picta, затем E. eschscholtzu croceater и, наконец, E. eschscholtzu klauberi (форма со множеством четких пятен, которую мы ранее назвали E. klauberi, считая, что это самостоятельный вид).
Стеббинс полагает, что предки Ensatina возникли в северной части Калифорнийской долины и по мере продвижения к югу по западной и восточной стороне эволюционировали в противоположных направлениях. Альтернативная схема предполагает, что они возникли на юге – в форме, например, E. eschscholtzii eschscholtzii – и эволюционировали, двигаясь по западной стороне долины на север и потом снова на юг с другой стороны. В результате на юге, на другом конце кольца, возникла форма E. eschscholtzii klauberi. Как бы то ни было, сейчас мы наблюдаем гибридизацию по всей длине кольца, кроме точки, где пересекаются два конца линии – далеко на юге Калифорнии.
Ситуация осложняется тем, что Калифорнийская долина, судя по всему, не является непреодолимым барьером для генов. Время от времени саламандры все-таки преодолевают его и некоторые популяции с западной стороны долины скрещиваются с популяциями на восточной стороне – например, xanthoptica на западе и platensis на востоке. Еще одна трудность заключается в том, что на юге есть небольшой разрыв кольца, где, судя по всему, саламандры не живут. Предположительно они там вымерли. Или, возможно, они живут там и сегодня, просто никто их не обнаружил: говорят, местность очень уж трудна для передвижения. И тем не менее, хотя географически кольцо и устроено не слишком просто, все же кольцо непрерывного генетического переноса – это основной паттерн генетики этого вида, как и в известных случаях с серебристыми чайками и клушами вокруг Северного полярного круга.
В Британии серебристая чайка и клуша четко различаются. Различия, которые трудно не заметить, касаются в основном цвета крыльев. У серебристых чаек они серебристо-серые, у клуш – темно-серые, почти черные. Сами птицы отлично различают друг друга и не скрещиваются, хотя иногда образуют смешанные колонии. Поэтому зоологи с чистой совестью дали им разные видовые названия: Larus argentatus и L. fuscus.
Кое в чем эти птицы напоминают саламандр. Если проследить популяцию серебристой чайки на запад к Северной Америке, через Сибирь и снова до Европы, мы заметим, что по мере движения вокруг полюса серебристые чайки все меньше напоминают серебристых чаек и становятся все сильнее похожими на клуш. Западноевропейские клуши – на самом деле продолжение кольца, которое начинается с серебристых чаек. В каждой точке кольца птицы похожи на непосредственных соседей достаточно, чтобы скрещиваться с ними – но лишь до тех пор, пока не достигнута крайняя точка, в которой кольцо замыкается. В Европе серебристая чайка и клуша не скрещиваются, хотя и связаны непрерывным кольцом скрещивания своих родственников.
Кольцевые виды, например саламандры и чайки, демонстрируют в пространственном измерении то, что обычно происходит во временном измерении. Представьте, что мы и шимпанзе – кольцевой вид. В этом нет ничего невозможного: кольцо могло идти на север с одной стороны Восточно-Африканской рифтовой долины и на юг – с другой. При этом на южном конце могли сосуществовать два вида, объединенных непрерывным континуумом скрещивания. Если бы дело и впрямь обстояло таким образом, то как это повлияло бы на наше отношение к другим видам? И вообще на наше отношение к дискретности видов?
Многие правовые и этические нормы основаны на противопоставлении Homo sapiens остальным видам. Многие из тех, кто считает аборты грехом (включая тех, кто убивает врачей и взрывает клиники), без зазрения совести едят мясо и не задумываются о шимпанзе, которых запирают в зоопарках и умерщвляют в лабораториях. Может быть, они призадумались бы, увидев непрерывную цепь звеньев между нами и шимпанзе, где каждое звено скрещивается с соседним (как в случае калифорнийских саламандр)? Однако, к несчастью, все промежуточные формы вымерли. И лишь поэтому ничто не мешает нам воображать, что между нашими видами (и любыми другими видами) лежит огромная пропасть.
Выше я рассказывал об озадаченном юристе, расспрашивавшем меня после лекции. Если вид А эволюционирует в вид Б, рассуждал он, то должен быть момент, когда детеныш принадлежит к новому виду Б, а его родители – к виду А. Представители разных видов не могут скрещиваться друг с другом – но ясно, что детеныш не будет настолько сильно отличаться от родителей, чтобы быть неспособным скрещиваться с представителями их вида. Не опровергает ли это основную идею теории эволюции?
Это все равно, что сказать: “Когда мы разогреваем чайник с холодной водой, нельзя уловить момент, когда вода перестает быть холодной и становится горячей – поэтому мы никогда не сможем заварить себе чаю”. Поскольку я всегда стараюсь сделать беседу конструктивной, я рассказал юристу о серебристых чайках, и, по-моему, это его заинтересовало. Он настаивал, что особи всегда должны относиться к тому или иному виду, и не допускал возможности, что какая-либо особь может находиться на полпути (или десятой части пути) от А к Б. Именно такая ограниченность мышления служит причиной нескончаемых споров о том, в какой момент развития эмбрион становится человеком (и с какого момента, соответственно, аборт приравнивается к убийству). Бесполезно говорить таким людям, что наше отношение к плоду – то, считаем ли мы его “наполовину человеком” или “на одну сотую часть человеком”, – зависит от того, какие признаки человека нас интересуют. Для ума, оперирующего абсолютными понятиями, “человек” всегда подобен “алмазу”. Такой подход не допускает промежуточных форм. Абсолютистское мышление может быть очень опасным и приводить к настоящим страданиям. Это я и называю тиранией дискретного мышления.
В некоторых случаях названия и дискретные категории полезны. Например, юристам они необходимы. Детям нельзя водить машину, взрослым – можно. Закон должен отличать ребенка от взрослого – например, устанавливая возрастной ценз. Очень показательно, что разные страховые компании по-разному определяют этот рубеж.
Некоторые вещи реальны всегда. Вы – один человек, я – другой, и наши имена – это ярлыки, указывающие на нашу самостоятельность. Монооксид углерода на самом деле отличается от диоксида углерода. Здесь нет перекрывания. Молекула состоит либо из одного атома углерода и одного – кислорода, либо из одного атома углерода и двух – кислорода. Нет молекул с полутора атомами кислорода. Один газ смертельно ядовит, а другой нужен растениям, чтобы производить органические вещества, от которых все мы зависим. Золото действительно отличается от серебра. Кристаллы алмаза отличны от кристаллов графита. Оба эти вещества состоят из углерода, но атомы углерода в них расположены совершенно иначе. Здесь нет промежуточных вариантов.
Но часто дискретность совсем не очевидна. Вот что я прочитал в газете во время недавней эпидемии гриппа. Или то была не эпидемия?
Согласно официальной статистике, 144 человека из каждых 100 тысяч болеют гриппом, – сообщила представитель Министерства здравоохранения. Поскольку обычный эпидемический порог – 400 человек на 100 тысяч, правительство не оценивает ситуацию как эпидемию. Однако она добавила: “Профессор Дональдсон придерживается мнения, что это эпидемия. Он считает, что заболевших гораздо больше, чем 144 человека на 100 тысяч. Все это очень неоднозначно и зависит от определения… Профессор Дональдсон, посмотрев на свой график, заявил, что это – серьезная эпидемия”.
Мы знаем, что некоторое количество людей болеет гриппом. Неужели этого недостаточно? Нет, для представителя министерства важен вопрос о том, считать ли это эпидемией. Превысило ли количество заболевших границу – 400 человек на 100 тысяч? Это, конечно, величайшее решение, и принять его может лишь профессор Дональдсон. Можно подумать, что от этого что-нибудь изменится!
На самом деле у эпидемий существует естественная граница: критическое количество зараженных особей, по достижении которого вирус или бактерия резко начинают распространяться с повышенной скоростью. Вот почему чиновники так стараются сделать прививки – например от коклюша – большему количеству людей, чем пороговое значение. Цель не только в том, чтобы обеспечить защиту привитым людям. Это должно помешать патогену достигнуть критической массы. В случае эпидемии гриппа представителя министерства должен волновать вопрос, пересек ли вирус гриппа рубеж и увеличилась ли скорость его распространения. И вопрос этот нужно решать любым способом, но без ссылки на волшебные числа вроде 400 на 100 тысяч. Зацикленность на волшебных числах – признак дискретного мышления. Забавно, что в данном случае дискретное мышление упускает из виду подлинную дискретность – границу, после которой эпидемия набирает обороты. Обычно в природе нет дискретности, которую можно пропустить.
Многие западные страны страдают от “эпидемии ожирения”. Ее проявления очевидны, но мне не нравится манера выражать ее в цифрах. Некоторая часть населения описывается как “клинически тучные” (clinically obese). Тут снова дискретное мышление настаивает на делении людей на тучных и нетучных. Но в жизни все устроено не так. Тучность – не дискретная величина. Мы можем оценить тучность каждого человека и затем вычислить статистику для группы. Подсчет числа людей, которые находятся выше некоего произвольно выбранного порога тучности, совершенно не показателен – хотя бы потому, что для этого нужно устанавливать порог и, возможно, его пересматривать.
То же дискретное мышление лежит в основе официальных данных о количестве людей, живущих “за чертой бедности”. Мы можем оценить бедность семьи на основе данных о ее доходе, предпочтительно в пересчете на вещи, которые эти люди реально могут купить. Можно сказать: “Икс беден, как церковная мышь” или “Игрек богат, как Крез” – и все поймут. Но иллюзорно точный подсчет числа людей, живущих по ту или иную сторону произвольно выбранной “черты бедности”, не имеет смысла: точность, которую предполагает подсчет процентов, противоречит бессмысленной искусственности “черты”. Черта – побочный эффект дискретного мышления. Еще деликатнее вопрос о различении “чернокожих” и “белых” в современном обществе, особенно американском (см. “Рассказ Кобылки”).
Еще пример. Английские университеты присуждают дипломы первой, второй и третьей категорий. В других странах университеты делают примерно то же: A, B, C и так далее. И вот что я хочу сказать. Студенты не делятся на хороших, посредственных и плохих. Не существует таких категорий. Экзаменаторы прилагают немало усилий, чтобы оценить успеваемость по шкале, выставляя оценки или баллы, которые затем добавляют к другим оценкам или обрабатывают иным способом. Баллы несут гораздо больше информации, чем классификация по одной из трех категорий. Однако в итоге оглашаются только дискретные категории.
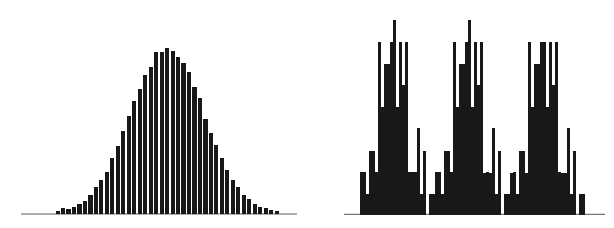
В большой выборке распределение способностей и умений студентов образует кривую нормального распределения, где несколько студентов успевают очень хорошо, несколько – очень плохо, а большинство находится между ними. Эта кривая может не быть симметричной, но чем больше выборка, тем более гладкой будет кривая.
Некоторые экзаменаторы (особенно – надеюсь, меня простят – по гуманитарным предметам), похоже, и вправду верят в существование дискретной сущности “первоклассное мышление”. Они считают, что у студента оно либо есть, либо нет. Задача экзаменатора состоит в том, чтобы отделить первых от вторых, а вторых от третьих, – в некотором роде отделить агнцев от козлищ. Таким людям сложно понять, что в реальности существует непрерывный ряд переходных форм от чистых агнцев к чистым козлищам.
Если бы, вопреки моим ожиданиям, оказалось, что с увеличением выборки студентов распределение оценок приближается к дискретному распределению с тремя пиками, то было бы замечательно. Практика присуждения дипломов первой, второй и третьей степеней была бы оправданной. Но это не подтверждается – что неудивительно, принимая во внимание человеческое разнообразие. Разница между высшей и низшей точками одной категории сильнее, чем разница между высшими точками разных категорий. Куда справедливее было бы оглашать как-нибудь обработанные оценки. Однако дискретное мышление требует, чтобы мы помещали людей в ту или иную категорию.
Вернемся от агнцев и козлищ к эволюции, то есть к овцам и козам. Действительно ли виды дискретны – или же они переходят один в другой? Если рассматривать лишь современных животных, то ответ будет – да, виды дискретны. Исключения, например чайки и калифорнийские саламандры, встречаются редко. Но они очень показательны, потому что переводят понятие непрерывности, которое обычно относится к области времени, в пространство. Людей и шимпанзе, безусловно, связывает общий предок и непрерывная цепь промежуточных звеньев. Однако промежуточные звенья вымерли, а то, что осталось, формирует дискретное распределение. То же верно для людей и нечеловекообразных обезьян, для людей и кенгуру – за тем исключением, что промежуточные звенья вымерли раньше. Поскольку почти все промежуточные звенья уже вымерли, мы обычно с чистой совестью считаем, что виды дискретны. Однако нас интересует эволюционная история не только живых, но и мертвых. Когда мы рассматриваем всех когда-либо живших животных, а не только тех, что живут сейчас, мы видим, что любые два вида связывают непрерывные линии преемственности. Если заглянуть в историю, окажется, что даже такие очевидно дискретные современные виды, как овцы и собаки, связаны через общего предка непрерывной линией преемственности.
Эрнст Майр, выдающийся эволюционист XX века, называл иллюзию дискретности (философы называют ее эссенциализмом) главной причиной того, почему люди так поздно осознали идею эволюции. Платон, которого можно считать вдохновителем эссенциализма, полагал, что реальные вещи – это несовершенные отпечатки вечных идей. Где-то существует совершенный, идеальный кролик, который имеет такое же отношение к реальному кролику, как идеальный круг в математике – к кругу, нарисованному в пыли. И по сей день многие верят, что овцы есть овцы, а козы – это козы, и ни один вид не может дать начало другому, потому что в этом случае они должны были бы изменить свою “сущность”. Однако нет такой вещи, как сущность.
Ни один эволюционист не утверждает, что одни современные виды могут превращаться в другие. Кошки не превращаются в собак, а собаки – в кошек. Просто кошки и собаки произошли от общего предка, жившего десятки миллионов лет назад. Если бы промежуточные формы не вымерли, попытка отделить кошек от собак была бы обречена на провал, как в случае саламандр и чаек. Идеальные сущности тут ни при чем: отделение кошек от собак возможно лишь благодаря тому счастливому (с точки зрения приверженца эссенциализма) факту, что переходные формы вымерли. Платон наверняка посмеялся бы, узнав, что на самом деле именно несовершенство – случайность вымирания – позволяет отделить один вид от другого. То же относится и к отделению человека от наших родственников. Если бы мы обладали полной информацией и об ископаемых, и о современных видах, названия животных стали бы невозможными. Вместо названий нам пришлось бы использовать подвижные шкалы – как в случае слов “теплый”, “прохладный” и “холодный”, которые мы заменяем подвижными шкалами Цельсия и Фаренгейта.
Сейчас мыслящие люди принимают эволюцию как данность, и можно было бы надеяться, что эссенциализм в биологии будет наконец преодолен. Но, увы, пока этого не произошло. Эссенциализм не хочет сдавать позиции. Никто не спорит с тем, что Homo sapiens – это один вид (а многие скажут, что и род), а Pan troglodytes (шимпанзе) – другой. Но никто не спорит и с тем, что, если проследить линии человека и шимпанзе до общего предка, промежуточные формы сформируют континуум, в котором каждое поколение будет способно к скрещиванию со своими родителями и детьми.
Если следовать критерию скрещиваемости, каждая особь относится к тому же виду, что и ее родители. Это заключение кажется ожидаемым, если не сказать очевидным – пока мы не осознаем, что с точки зрения эссенциализма это недопустимое противоречие. Большинство наших предков на протяжении их эволюционной истории по всем критериям принадлежали к отличным от нас видам, и мы, разумеется, не могли бы с ними скрещиваться. В девоне, например, наши предки были рыбами. И, хотя скрещивание между рыбами и нами невозможно, мы связаны непрерывной цепочкой предков.
Теперь становится ясно, насколько бессодержательны споры о названиях ископаемых гоминид. Homo ergaster всеми признается предковым видом H. sapiens, и я буду следовать этой точке зрения. В принципе, есть определенный смысл в том, чтобы считать H. ergaster самостоятельным видом, даже если это невозможно проверить. Это значит, что если мы вернемся в прошлое и встретим H. ergaster, мы не сможем с ним скреститься . Но представьте: вместо того чтобы сразу направиться к H. ergaster или любому другому вымершему предковому виду, мы останавливаемся каждую тысячу лет, подбираем в машину времени молодую фертильную особь и переносим на тысячу лет назад, к следующей остановке. Там мы его (ее) высаживаем. При условии, что этот путешественник (путешественница) приспособится к новым социальным и языковым традициям (а это довольно трудно), он или она вполне смогли бы скреститься с представителем противоположного пола, жившим тысячей лет раньше. Теперь подберем женщину и перевезем ее еще на тысячу лет назад. Наша пассажирка будет способна родить ребенка от мужчины, который жил на тысячу лет раньше нее. Эта цепочка будет тянуться вплоть до времени, когда наши предки плавали в море – то есть до самых рыб. Каждый пассажир, перенесенный на тысячу лет назад, мог бы скрещиваться со своими предшественниками. Однако в какой-то момент – возможно, около миллиона лет назад – мы, современные люди, уже не смогли бы скрещиваться с предками, хотя промежуточный пассажир смог бы. И именно в этот момент мы смогли бы сказать, что встретились с другим видом.
Этот барьер возник не сразу. Нет поколения, в котором ребенок относился бы к Homo sapiens, а его родители – к H. ergaster. Можете считать это противоречием, но не бывает так, чтоб ребенок и родители относились к разным видам – хотя цепочка родителей и детей тянется назад во времени от людей до рыб и дальше. На самом деле противоречие тут видят лишь закоренелые сторонники эссенциализма. Вообще-то это не более парадоксально, чем тот факт, что ребенок, который только что был маленьким, “вдруг” становится большим. Или тот факт, что чайник перестал быть холодным и стал горячим. Юрист, конечно, захочет установить границу между ребенком и взрослым – полночь на восемнадцатый день рождения или что-то в этом духе. Но всем очевидно, что эта граница иллюзорна (хотя и нужна для некоторых целей). Если бы только люди могли понять, что то же самое относится и к, например, эмбриону, который в один прекрасный момент “становится” человеком!
Креационисты очень любят “пробелы” в палеонтологической летописи. Однако им невдомек, что у биологов есть собственные причины любить эти пробелы. Без них систематика просто развалится. Мы бы не смогли давать имена ископаемым, и нам пришлось бы оперировать числами или точками на графике. И вместо споров о том, является ли ископаемое ранним H. ergaster или поздним H. habilis, мы называли бы их H. habigaster. Об этом можно долго говорить. Однако наш мозг эволюционировал в мире, где большинство вещей принадлежат к дискретным категориям – и где большинство промежуточных форм между современными видами уже вымерли. Поэтому нам удобнее наделять каждую вещь своим названием. Я не исключение, и вы тоже. Я не буду лезть из кожи вон, стараясь избегать дискретных названий видов. Но “Рассказ Саламандры” объясняет, почему дискретность – это скорее свойство человеческого мышления, чем свойство природы. Давайте использовать названия так, будто они отражают дискретную реальность, но не забывать, что (по крайней мере в отношении эволюции) это условность, обусловленная ограниченностью нашего сознания.
Назад: Рандеву № 17 Амфибии
Дальше: Рассказ Узкорота

