Звероящеры
После того, как к нам присоединились однопроходные, мы отправляемся в путь – и преодолеваем без остановки 130 млн лет. Это самый длинный переход. Далее мы встретим группу пилигримов более многочисленную, чем наша. Это завропсиды: рептилии и птицы, то есть почти все позвоночные, которые откладывают на суше крупные яйца с водонепроницаемой скорлупой. “Почти” – это потому, что однопроходные тоже откладывают яйца. Даже черепахи (во всех других отношениях морские животные) выходят на берег, чтобы отложить яйца. Плезиозавры, возможно, делали то же самое. Ихтиозавры же были настолько специализированными пловцами, что, по-видимому, не могли выбираться на берег – как и дельфины, их современные двойники. Они независимо “изобрели” живорождение – об этом мы узнали благодаря матерям-ихтиозаврам, окаменевшим в процессе родов.
Я сказал, что мы прошли без остановки 130 млн лет. Но, разумеется, “без остановки” – это допущение, поскольку остановкой мы признаем только встречи с ныне живущими пилигримами. Линия наших предков в течение этих 130 млн лет бурно развивалась, давая ветви. Об этом мы знаем из богатой палеонтологической летописи. Но ни одну из этих ветвей нельзя считать “рандеву”: представителей этих ветвей, которые могли бы стать пилигримами и “отправиться” из современности в прошлое, не осталось. Столкнувшись с подобной проблемой в отношении гоминид, мы решили наделить некоторые ископаемые почетным статусом “призрачных пилигримов”. И, поскольку мы ищем предков и хотим знать, как выглядел наш прародитель в 100-миллионном поколении, мы не можем игнорировать звероящеров и перейти прямо к сопредку № 16. Он был похож на ящерицу. Промежуток между ним и сопредком № 15, который был похож на землеройку, слишком велик, чтобы можно было оставить его без внимания. Нам придется встретиться со звероящерами, “призрачными пилигримами” – хотя рассказов от них мы, конечно, не дождемся. Но сначала – справка об интересующем нас отрезке времени.
Он охватывает половину юрского периода, весь триас, всю пермь и последние 10 млн лет каменноугольного периода. Сначала пилигримы перемещаются из середины юрского периода в сухой мир триаса – один из самых жарких периодов в истории планеты. Тогда вся суша соединилась, образовав Пангею. Здесь мы наблюдаем массовое вымирание конца триаса, во время которого исчезли три четверти видов. Но это ничто по сравнению со следующим переходом, в конце которого мы перемещаемся из триаса на один период назад: в пермь. На границе пермского и триасового периодов вымерло 90 % видов. Все они, включая трилобитов и несколько других крупных групп животных, не оставили потомков. Справедливости ради заметим, что трилобиты довольно давно находились на грани исчезновения. Но все же вымирание конца перми было самым разрушительным в истории. В Австралии сохранились некоторые свидетельства того, что это вымирание, как и меловая катастрофа, было вызвано столкновением массивного небесного тела с Землей. Даже насекомые тогда понесли серьезные потери – единственный раз в истории. В море сообщества придонных организмов были уничтожены почти полностью. На суше роль Ноя среди звероящеров досталась листрозавру (Lystrosaurus). Сразу после катастрофы приземистый короткохвостый листрозавр расселился по миру, быстро захватив освободившиеся экологические ниши.
Неизбежно возникают ассоциации с апокалипсисом. Но не стоит драматизировать. Вымирание неизбежно случается почти со всеми видами. Вероятно, 99 % видов, когда-либо живших на Земле, вымерли. Однако доля вымираний видов на миллион лет непостоянна и лишь иногда превышает 75 %: это значение принято считать границей массовых вымираний. Таким образом, массовые вымирания – это пики на графике скорости вымираний.
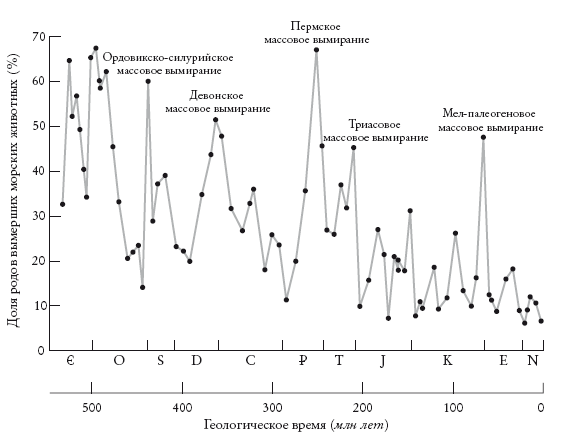
Доля родов морских животных, вымерших в фанерозое. Sepkoski [260].
На графике показано количество вымираний на миллион лет. Видно, что пикам соответствуют явно не самые приятные события. Возможно, это катастрофы – как, например, столкновение с небесным телом 65 млн лет назад, которое привело к гибели динозавров. Вероятно, в некоторых случаях вымирание не было внезапным. Ричард Лики и Роджер Левин говорят о “шестом вымирании”, причиной которого стал Homo sapiens или, как говорил Уильям Картрайт, мой учитель немецкого, Homo insipiens [человек неразумный].
Прежде чем перейти к звероящерам, разберемся с терминами. Термины “рептилия” и “млекопитающее” могут относиться как к кладам, так и к градам – это не взаимоисключающие понятия. Клада – это группа, состоящая из всех потомков одного предка. Например, птицы – это клада. “Рептилии” в традиционном понимании не являются настоящей кладой, потому что в эту группу не входят птицы. Поэтому биологи называют рептилий парафилетической группой. Некоторые рептилии (например, крокодилы) ближе к нерептилиям (птицам), чем к другим рептилиям (черепахам). То общее, что есть у всех рептилий, объединяет их в граду – но не в кладу. Града – это группа животных, которые достигли сходного уровня эволюционного развития.
Еще один пример грады – так называемый герп. Герпетология – это наука о рептилиях (кроме птиц) и амфибиях. “Герп” – необычная грамматическая конструкция: это сокращение, для которого нет полной формы. Герп – это животное, которое изучает герпетолог. Разумеется, это не самый правильный способ описания животного. Самое близкое по значению слово – библейские “гады”.
“Рыбы” – это акулы, различные вымершие животные, костистые рыбы (форель, щука и другие) и целаканты. Форель на самом деле ближе к человеку, чем к акулам (а целаканты – еще более близкие родственники людей, нежели форель). Таким образом, “рыбы” – это не клада, потому что в нее не включен человек (и вообще все млекопитающие, птицы, рептилии и амфибии). Рыбы – это града, объединяющая животных, похожих на рыб. Но предложить точное определение грады почти невозможно. Например, ихтиозавры и дельфины похожи на рыб и, вполне возможно, напоминают рыб по вкусу. Но они не считаются членами грады “рыбы”, поскольку вернулись к “рыбьему” образу жизни, пройдя “нерыбью” стадию эволюции.
Понятие грады очень полезно, если вы твердо верите, что эволюция идет только в одном направлении – в сторону прогресса, следуя параллельными путями от одной отправной точки. Если вы, например, считаете, что все родственные линии независимо и параллельно эволюционировали от амфибийных форм к рептильным и затем к млекопитающим, можно говорить о том, что после грады рептилий животные попадают в граду млекопитающих. Возможно, подобное действительно происходило. Именно эта точка зрения стала первой, с которой я познакомился, – благодаря моему уважаемому преподавателю палеонтологии позвоночных Гарольду Пьюзи.
Если мы перейдем к другой крайности и будем мыслить в терминах строгой кладистики, то слово “рептилия” будет иметь право на существование лишь в том случае, если мы включим в эту группу птиц. Этой точки зрения придерживаются участники проекта “Древо жизни”, который был организован братьями Мэдисон. Об этом проекте можно говорить долго – как и о замене термина “звероящер” на “ящерозверь”. Но слово “рептилии” настолько привычно, что, боюсь, попытка что-то изменить приведет к неразберихе. Кроме того, иногда строгие кладистские принципы могут приводить к нелепым результатам. Вот пример. У сопредка № 16 должны быть прямые потомки и на ветви млекопитающих, и на ветви ящериц, динозавров, крокодилов. Эти потомки неизбежно были практически идентичны друг другу. То есть в течение некоторого времени они были способны скрещиваться друг с другом. И все же строгий кладист будет настаивать, чтобы одного из этих потомков называли завропсидом, а другого – млекопитающим. К счастью, на практике редко приходится сталкиваться с такими крайностями. Этот пример можно использовать в беседах с кладистами-радикалами, когда они задирают нос.
Мы привыкли к тому, что млекопитающие – преемники динозавров. Поэтому тот факт, что звероящеры процветали и до появления динозавров, может показаться удивительным. Звероящеры занимали те же ниши, которые позднее захватили динозавры, а после тех – млекопитающие. Фактически они занимали эти ниши не однажды, а в несколько подходов, разделенных вымираниями. В отсутствие рандеву с ныне живущими пилигримами я буду говорить о трех встречах с “призрачными пилигримами”. Эти встречи заполнят промежуток между сопредком № 15 (который похож на землеройку и объединяет нас с однопроходными) и сопредком № 16 (который похож на ящерицу и объединяет нас с птицами и динозаврами). Здесь я привожу рисунок, выполненный моим коллегой Томом Кемпом, одним из ведущих специалистов по звероящерам. На рисунке изображены некоторые из этих животных и их родственные связи.
Наш прародитель в 150-миллионном поколении, возможно, немного напоминал тринаксодона (Thrmaxodon), который жил в середине триасового периода. Его окаменелости найдены в Африке и Антарктиде – изначально частях Гондваны. Конечно, не стоит надеяться, что нашим предком был сам тринаксодон или другое конкретное ископаемое, которое нам посчастливилось найти. Тринаксодона, как и любое другое ископаемое, стоит считать скорее родственником нашего предка. Он относился к ционодонтам – одной из групп звероящеров. Цинодонты были настолько похожи на млекопитающих, что трудно удержаться от соблазна назвать их млекопитающими. Впрочем, не все ли равно? Цинодонты – почти идеальный пример промежуточного звена. А поскольку эволюция не стояла на месте, было бы странно, если бы не существовало промежуточных звеньев вроде цинодонтов.
Цинодонты – одна из нескольких групп, которые произошли от ранних звероящеров – терапсидов. Наш прародитель в 160-миллионном поколении был, вероятно, терапсидом и жил в пермском периоде. К сожалению, довольно трудно выбрать конкретное ископаемое, которое можно считать представителем терапсидов. Эти животные господствовали на суше до триаса, когда на смену им пришли динозавры. Но и в триасовом периоде они составляли серьезную конкуренцию динозаврам. Среди терапсидов были огромные животные: травоядные трехметровой длины и охотящиеся на них крупные хищники.
Наш предок-терапсид вряд ли был таким устрашающим животным. Как правило, крупные специализированные животные, например горгонопсиды с клыками-саблями или травоядные цинодонты (см. вкладку), не имеют эволюционного будущего. Чаще всего они относятся к 99 % видов, обреченных на вымирание. А 1 % видов – Нои, от которых происходят все остальные (независимо от того, какими крупными или необычными они становятся), – чаще всего маленькие.
Ранние терапсиды были не так похожи на млекопитающих, как их преемники цинодонты. Но все же они напоминали млекопитающих сильнее, чем их предшественники пеликозавры, которые обеспечили раннюю радиацию звероящеров. Предшественником терапсид в ряду предков – нашим прародителем в 165-миллионном поколении – почти наверняка был пеликозавр. Правда, в этом случае выбрать конкретный вид в качестве предка также не представляется возможным. Пеликозавры были древнейшей группой маммалоподобных рептилий. Они процветали в каменноугольном периоде. Самый известный из пеликозавров – это диметродон, тот, что с большим “парусом” на спине. Никто не знает, зачем диметродону был нужен этот “парус”. Возможно, он был чем-то наподобие солнечной батареи, которая помогала животному нагреваться до температуры, необходимой для работы мышц. А может, это был радиатор. Или же он мог использоваться для брачной демонстрации, выступая аналогом павлиньего хвоста. Почти все пеликозавры вымерли в пермском периоде – кроме пеликозавра-Ноя, который дал начало второй группе звероящеров – терапсидам. И все начало триасового времени переоткрывали формы, утраченные в поздней перми.
Пеликозавры были гораздо меньше похожи на млекопитающих, чем терапсиды. Например, пеликозавры ползали на брюхе, как ящерицы, с расставленными в стороны ногами. Походка у них, скорее всего, была виляющая, с рыбообразными движениями тела из стороны в сторону. Терапсиды, цинодонты и млекопитающие постепенно поднимались с земли: постановка ног становилась вертикальной, а походка все меньше напоминала движения рыбы на суше. Среди других признаков “маммализации” (которые, возможно, кажутся прогрессивными лишь самим млекопитающим) – следующие. Нижняя челюсть уменьшилась до единственной нижнечелюстной кости, а остальные ее кости перешли в ухо. В некоторый момент у наших предков возникли шерсть и способность к терморегуляции. Кроме того, появились молоко и родительская забота о детенышах, а также сложные специализированные зубы.
Рассматривая эволюцию звероящеров – “призрачных пилигримов”, – мы обсудили три группы этих животных, сменивших друг друга: пеликозавров, терапсидов и цинодонтов. Млекопитающие – четвертая группа. Но знакомые нам экотипы в группе млекопитающих появились гораздо позднее: спустя 150 млн лет. А до тех пор на Земле властвовали динозавры, которые прожили вдвое дольше, чем три волны звероящеров вместе.
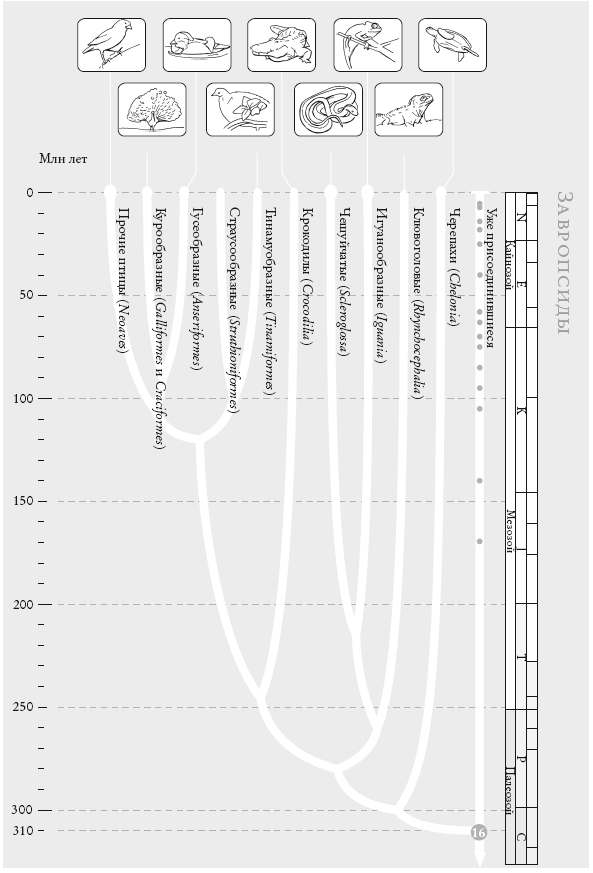
Завропсиды. Революцией в развитии наземных позвоночных стало появление амниона – непроницаемой для воды, но проницаемой для воздуха оболочки яйца. Имеются две рано разошедшиеся линии амниот: синапсиды (представлены млекопитающими) и завропсиды (17 тыс. живущих ныне видов «рептилий» и птиц). Филогения здесь достаточно устойчива.
На рис. (слева направо): средний земляной вьюрок (Geospiza fortis), обыкновенный павлин (Pavo cristatus), мандаринка (Aix galericulata), тинаму-пустынник (Tinamus solitarius), нильский крокодил (Crocodylus niloticus), африканский коралловый аспид (Thamnophis sirtalis parietalis), обыкновенный хамелеон (Chamaeleo chamaeleon), гаттерия (Sphenodon punctatus), зеленая черепаха (Chelonia mydas).
Назад: Что сказал Звездонос Утконосу
Дальше: Рандеву № 16 Завропсиды

