Книга: Рассказ предка
Назад: Возвращение Хозяина
Дальше: Ценностно-нейтральный и ценностно-нагруженный подход к прогрессу
Перезапуск эволюции
Американский биолог-теоретик Стюарт Кауфман указывал в статье 1985 года:
Один из способов подчеркнуть наше нынешнее невежество – это задать вопрос, как выглядели бы организмы через один-два миллиарда лет, если бы эволюция повторилась от докембрия, когда уже сформировались древние эукариотические клетки. И если бы этот эксперимент был проведен бесконечное число раз, какие свойства организмов появлялись бы снова и снова, какие оказались бы редкими, какие свойства легко возникали бы в процессе эволюции, а какие – с трудом? Основной недостаток современного видения эволюции заключается в том, что оно не привело нас к постановке вопросов, ответы на которые могли бы дать нам глубокое понимание прогнозируемых признаков.
Особенно мне нравится оговорка Кауфмана о статистике. Он предлагает не просто мысленный эксперимент, а ряд экспериментов, направленных на поиск всеобщих законов жизни, а не конкретных проявлений организмов. Вопрос Кауфмана сродни вопросу из научной фантастики о том, какой была бы жизнь на других планетах, – за исключением того, что на других планетах исходные и существующие условия были бы иными. На крупной планете гравитация создала бы совершенно новые направления давления отбора. Там животные размером с паука не смогли бы иметь паучьи ноги (они сломались бы под тяжестью животного) и нуждались бы в столпообразных конечностях, напоминающих ноги земных слонов. И, напротив, на маленькой планете животные размером со слона, однако имеющие более хрупкое телосложение, могли бы носиться и прыгать, как пауки-скакуны. Эти ожидания относительно телосложения применимы к целым статистическим выборкам миров с большой силой тяжести и миров с небольшой силой тяжести.
Сила тяжести – такое свойство планеты, на которое жизнь не может повлиять. К подобным свойствам относятся также: расстояние планеты от звезды и скорость ее обращения, которая определяет длительность суток; наклон ее оси, который на планете, имеющей почти круговую орбиту, является основным фактором, определяющим смену времен года. На планете с орбитой, отличной от круговой, например на Плутоне, резкое изменение расстояния до звезды – гораздо более важный фактор, определяющий сезонность. Наличие спутника (спутников), расстояние до него, его масса и орбита также оказывают трудноуловимое, но заметное влияние на жизнь из-за приливов и отливов. Все эти факторы являются данностью, на которую не может повлиять жизнь, и поэтому при многократных экспериментах Кауфмана их следует рассматривать в качестве констант.
Прежние поколения ученых рассматривали как данность также погодные условия и химический состав атмосферы. Сейчас известно, что состав атмосферы (особенно высокое содержание в ней кислорода и низкое – углерода) определяется жизнью. Поэтому наш мысленный эксперимент должен учитывать возможность того, что при “перезапусках” эволюции атмосфера будет меняться под влиянием той формы жизни, которая эволюционирует в данный момент. Вследствие этого жизнь может влиять на погоду и даже на такие климатические события, как ледниковые периоды и засухи. Мой коллега Билл Гамильтон (который оказывался прав слишком часто, чтобы смеяться над его догадками) предположил даже, что облака и дождь суть адаптация стремящихся к распространению микроорганизмов.
Насколько нам известно, на глубинные механизмы Земли не влияет “пена” жизни на ее поверхности. Однако мысленные эксперименты по “перезапуску” эволюции должны учитывать возможные различия в ходе тектонических событий и, следовательно, в истории движения континентов. Интересен также вопрос о вулканической активности, землетрясениях и метеоритных бомбардировках: должны ли мы считать их одинаковыми при последовательных “перезапусках” эксперимента Кауфмана? Наверное, разумно считать движение литосферных плит и столкновения с космическими объектами такими переменными, которые можно усреднить, взяв достаточно крупные статистические выборки “перезапусков”.
С чего же мы начнем, пытаясь ответить на вопрос Кауфмана? Какой была бы жизнь, если бы мы проиграли “запись” статистически значимое количество раз? Можно сразу выделить целый ряд вопросов возрастающей сложности. Кауфман выбрал точкой “перезапуска” часов момент формирования эукариотической клетки из бактериальных компонентов. Но мы можем начать “перезапуск” двумя или тремя эонами ранее – с возникновения самой жизни. Или, напротив, перезапустить часы гораздо позднее: скажем, в момент возникновения сопредка № 1, на месте нашего расхождения с шимпанзе. Тогда мы сможем проверить, появились бы у гоминид двуногость, крупный мозг, язык, цивилизация и бейсбол при статистически значимом количестве “перезапусков” – при исходном условии, что жизнь достигла сопредка № 1. А в промежутке между этими крайностями останется пространство для вопросов о происхождении млекопитающих, происхождении позвоночных, любых других загадок.
Было ли в истории жизни нечто похожее на кауфмановский эксперимент? Да. Во время путешествия мы увидели несколько таких естественных экспериментов. По счастливому стечению обстоятельств Австралия, Новая Зеландия, Мадагаскар, Южная Америка и даже Африка долгое время провели в географической изоляции, что предоставляет нам нечто вроде “перезапусков” основных эпизодов эволюции.
Эти массивы суши были изолированы друг от друга и от остального мира в течение долгих периодов времени после исчезновения динозавров – как раз тогда, когда группы млекопитающих продемонстрировали свой максимальный творческий потенциал. Изоляция не была полной, но достаточной для радиации афротериев и появления лемуров на Мадагаскаре. В случае Южной Америки мы выделили три этапа эволюции млекопитающих, разделенных длительными периодами изоляции. Австралинея предоставила самые идеальные условия для естественного эксперимента: ее изоляция в течение большей части нужного нам периода была почти совершенной, и началась она с появления там крошечной – возможно, представленной единственным экземпляром – группы сумчатых. Новая Зеландия представляет собой исключение, потому что она – единственная среди наших “экспериментальных площадок” – весь рассматриваемый период была лишена млекопитающих.
Глядя на эти эксперименты, я восхищаюсь главным образом тем, насколько предсказуемо ведет себя эволюция, если запустить ее дважды. Мы видели, как похож тасманийский сумчатый волк на собаку, сумчатый крот – на обычного крота, летающий поссум – на белку-летягу, а сумчатый саблезубый тигр – на саблезубого тигра (и различных “ложных саблезубых” представителей плацентарных хищников). Отличия также любопытны. Кенгуру – это прыгающая замена антилопы. Прыжки на двух ногах, доведенные в ходе эволюции до совершенства, могут быть такими же удивительно быстрыми, как и бег на четырех ногах. Но эти способы передвижения радикально отличаются друг от друга – причем их особенности обуславливают важнейшие различия анатомии. Предположительно на каком-то древнем перепутье любая из этих двух “экспериментальных” линий могла выбрать либо путь усовершенствования прыжков на двух ногах, либо путь развития бега на четырех конечностях. Так уж произошло (возможно, сначала почти случайно), что кенгуру стали прыгать, а антилопы – бегать. И теперь мы с удивлением смотрим на последовательные дивергенции того, что получилось.
Все эти различные по масштабам события радиации млекопитающих произошли примерно в одно и то же время на разных массивах суши. Уход динозавров предоставил им широкие возможности. Но и с самими динозаврами в свое время произошло подобное, хотя и с некоторыми упущениями: так, я не знаю, почему среди динозавров не найдено аналога современного крота. До динозавров тоже было много параллельных линий развития, особенно среди тероморфов, и они тоже приводили к появлению сходных морфологических типов.
Читая публичные лекции, я всегда пытаюсь ответить на вопросы аудитории. Чаще всего меня спрашивают, во что эволюционируют люди в будущем (и каждый вопрошающий, похоже, искренне считает этот вопрос оригинальным). У меня каждый раз сжимается сердце: именно этого вопроса должен избегать эволюционист. Мы не можем предсказать эволюцию видов – мы можем лишь спрогнозировать вымирание большинства видов. Однако, хотя мы не в состоянии предсказать будущее, скажем, на 20 млн лет вперед, мы все-таки можем предугадать общий характер экологических типов, которые будут распространены в то время. Среди них будут травоядные и хищные животные, те, кто питается травой и побегами, мясом, рыбой и насекомыми. Эти прогнозы пищевых предпочтений предполагают, что через 20 млн лет на Земле все еще будет соответствующая пища. Присутствие животных, питающихся побегами, предполагает существование деревьев, а насекомоядных животных – наличие насекомых или мелких беспозвоночных. В пределах каждой категории – травоядных, хищных и так далее – будет наблюдаться диапазон различных размеров. Животные будут бегающими, летающими, плавающими, ползающими и роющими. Виды не будут такими, как сегодня. Но диапазон типов будет сходен с нынешним, и разнообразие образов жизни – тоже.
Если в следующие 20 млн лет произойдет крупная катастрофа и вымирание, сопоставимое по масштабам с вымиранием динозавров, мы можем ожидать, что диапазон экотипов будет получен из новых предковых точек. При этом – вопреки моим рассуждениям о грызунах (рандеву № 10) – непросто угадать, какие из современных животных явятся этими отправными точками. На карикатуре викторианской эпохи изображен профессор Ихтиозавр, читающий лекцию о древнем человеческом черепе. Если бы в эпоху динозавров профессор Ихтиозавр взялся бы обсуждать их катастрофический конец, ему было бы трудно предсказать, что место динозавров займут потомки млекопитающих, которые в те времена были мелкими ночными насекомоядными.
Конечно, это касается недавней эволюции: речь не идет о масштабном “перезапуске” по Кауфману. Но и эти “перезапуски” могут многое рассказать о воспроизводимости эволюции. Если ранняя эволюция шла примерно так же, как поздняя, то информация, которую мы получим, может дать нам общие принципы. Предполагаю, что принципы, которые мы обнаруживаем в недавней (после исчезновения динозавров) эволюции, вероятно, воплощаются по меньшей мере до кембрия и, возможно, до момента появления эукариотической клетки. Я думаю, что параллелизм радиации млекопитающих в Австралии, Южной Америке, Африке, Азии и на Мадагаскаре может дать что-то вроде шаблона для ответа на вопросы Кауфмана, относящиеся к гораздо более ранним отправным точкам, например к выбранной им самим – появлению эукариотической клетки. Что касается раннего времени, то в его отношении уверенности нет. Марк Ридли в книге “Демон Ыенделя” указывает, что появление эукариот стало в высшей степени маловероятным событием – возможно, оно еще невероятнее, чем происхождение самой жизни. Держу пари, что большинство мысленных “перезапусков” эволюции с момента возникновения жизни не приведут к “эукариократии”.
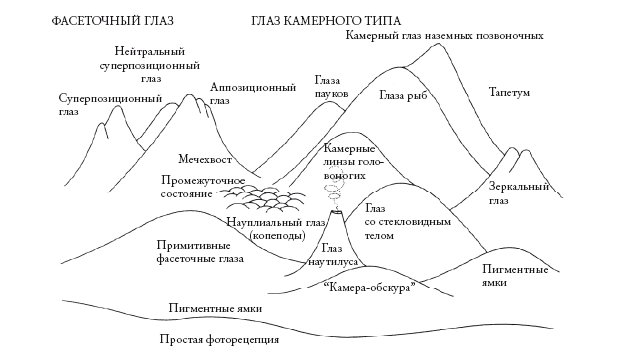
Чтобы изучить конвергенцию, не обязательно полагаться на географическое разделение, как в австралийском естественном эксперименте. Мы можем провести эксперимент, “перезапуская” эволюцию не из одной точки в разных географических областях, а из разных точек в одной области. Ведь конвергенция наблюдается у животных, настолько не связанных друг с другом, что паттерны не имеют отношения к географическому разделению. Согласно некоторым оценкам, “глаз” независимо появился в животном мире 40–60 раз. Этому вопросу я посвятил главу в книге “Восхождение на пик Невероятности” и не буду здесь повторяться. Скажу лишь, что профессор Майкл Лэнд из Университета Сассекса, ведущий специалист по сравнительной зоологии глаза, выделяет девять видов оптического механизма, каждый из которых появлялся в эволюции более одного раза. Профессор любезно подготовил для этой книги схему, где пики соответствуют случаям независимого появления глаза.
Складывается впечатление, что жизнь изо всех сил стремится к эволюции глаза (см. вкладку). Мы можем с уверенностью сказать, что статистически значимая доля “перезапусков” по Кауфману привела бы к появлению глаза. И не просто глаза, а сложного глаза, похожего на глаза насекомых, креветок или трилобитов, и камерного глаза, как у нас или кальмаров, с цветовым зрением и механизмами настройки фокуса и диафрагмы. Также с высокой долей вероятности появились бы глаза, похожие на рефлектор радиолокационной антенны, как у морских блюдечек, и глаза-обскуры, как у наутилуса – современного, похожего на аммонита моллюска с плавучей спиральной раковиной (см. рандеву № 26). И если на других планетах есть жизнь, можно смело ставить на то, что и там есть глаза, работающие по знакомым нам оптическим принципам. Есть не так много способов получить глаз, и жизнь наверняка перепробовала их все.
Такого рода оценки допустимы и для других адаптаций. Эхолокация появлялась не меньше четырех раз: у летучих мышей, усатых китов, птиц гуахаро и пещерных стрижей. Это происходило не чаще эволюции глаза, но все же достаточно часто, чтобы счесть, что при подходящих условиях она появится снова. Вполне возможно, что “перезапуски” эволюции снова и снова открывали бы одни и те же приемы преодоления трудностей. Здесь я тоже не буду повторяться (см. мою книгу “Слепой часовщик”), а подведу итог. Чего стоило бы ожидать от “перезапусков”? Многократно эволюционировала бы эхолокация с использованием высокочастотных сигналов (которые обеспечивают лучшее разрешение, чем низкочастотные). Сигналы, по крайней мере у некоторых видов, вероятно, будут частотно-модулированными, понижающимися или повышающимися (это обеспечивает точность распознавания благодаря разнице между первым и последним участками). Вычислительный аппарат, который используют для анализа эха, вполне мог бы проводить (конечно, подсознательно) подсчеты, основанные на допплеровском смещении частоты эха. Ведь эффект Допплера универсален для любой планеты, где есть звук, и летучие мыши в совершенстве им владеют.
Как узнать, что глаз или эхолокация эволюционировали независимо? Изучая генеалогическое древо. Родственники гуахаро и пещерных стрижей не пользуются эхолокацией. Гуахаро и пещерные стрижи независимо перешли к жизни в пещерах. Мы знаем, что эхолокация развилась у них независимо, потому что никто из их родственников это не практикует. Возможно, у разных групп летучих мышей этот навык эволюционировал независимо. Мы не знаем, сколько раз появилась эхолокация. У некоторых землероек и тюленей эта способность имеется в рудиментарной форме (она есть и у некоторых слепых людей). Пользовались ли эхолокацией птеродактили? Вполне возможно: ночью можно много чем поживиться, а летучих мышей в то время еще не было. То же относится и к ихтиозаврам. Внешне они напоминали дельфинов и, вероятно, вели сходный с ними образ жизни. Поскольку дельфины интенсивно используют эхолокацию, логично спросить, делали ли это ихтиозавры. Прямых доказательств нет. Существует один довод против: у ихтиозавров были очень большие глаза (одна из их самых заметных черт). Это, возможно, свидетельствует о том, что они полагались в основном на зрение, а не на эхолокацию. У дельфинов, напротив, глаза не очень крупные, а одна из их самых заметных особенностей – шишка (“дыня”) над рылом, которая работает как акустическая “линза”: она собирает звук в узкий, направленный вперед пучок.
Как любой зоолог, я могу просмотреть свою мысленную базу данных царства животных и приблизительно ответить на вопросы вроде: “Сколько раз в эволюции независимо появился признак икс?” Если провести подсчеты более систематично, из этого мог бы получиться хороший исследовательский проект. Оказалось бы, что некоторые признаки появились “много раз” (как глаз) или “несколько раз” (как эхолокация). Некоторые – “лишь однажды” или даже “никогда”, – хотя такие примеры найти неожиданно трудно. Различия могут быть весьма любопытными. Я подозреваю, мы открыли бы эволюционные пути, которыми сильнее всего “желает” следовать жизнь. Другие пути имеют более сильное “сопротивление”. В “Восхождении на пик Невероятности” я привел аналогию: огромный музей жизни (реальной и потенциальной), в котором коридоры во многих измерениях представляют собой эволюционные изменения (реальные и потенциальные). Некоторые из коридоров – широкие, манящие. Другие перегорожены барьерами, которые трудно или даже невозможно преодолеть. Эволюция многократно устремляется широкими коридорами и время от времени неожиданно преодолевает один из барьеров.
А теперь рассмотрим несколько признаков, для которых стоит провести подсчет. Ядовитое жало, впрыскивающее яд под кожу, независимо эволюционировало не менее десяти раз: у медуз и их родственников, у пауков, скорпионов, многоножек, насекомых, моллюсков (конусы), змей, акул (хвостоколовые скаты), костистых рыб (бородавчатка), млекопитающих (самцы утконосов) и растений (крапива). Держу пари, что яд, в том числе его подкожное впрыскивание, не раз появлялся бы в экспериментах по “перезапуску” эволюции.
Производство звуков в социальных целях независимо развилось у птиц, млекопитающих, сверчков и кузнечиков, цикад, рыб и лягушек. Электролокация – использование слабых электрических полей для ориентации в пространстве – эволюционировала несколько раз (см. “Рассказ Утконоса”). Несколько раз независимо эволюционировало использование электрического тока в защитных целях – вероятно, возникшее на основе электролокации. Мы довольно уверенно можем делать ставку на повторную эволюцию существ, использующих электричество для ориентирования и в качестве оружия.
Истинный машущий полет, в противоположность пассивному планированию или парашютированию, эволюционировал, судя по всему, четыре раза: у насекомых, птерозавров, летучих мышей и птиц. Парашютирование и планирующий полет различных типов независимо развился, вероятно, не менее ста раз и, возможно, является эволюционным предшественником истинного полета. Он встречается, например, у ящериц, лягушек, змей, “летучих” рыб, кальмаров, шерстокрылов, сумчатых и грызунов (дважды). Я бы мог поставить крупную сумму на то, что в условиях “перезапуска” эволюции по Кауфману планирующие животные появлялись бы снова и снова. (Что касается истинного машущего полета, то я ограничился бы средней суммой.)
Реактивная тяга, вероятно, появилась дважды: у головоногих моллюсков (причем кальмары развивают высокую скорость) и гребешков. Последние обитают преимущественно на морском дне, но могут и плавать. Они ритмично открывают и закрывают створки раковины, щелкая ими, как кастаньетами. Можно было бы предположить (я, например, так и делаю), что они при этом двигаются “назад”, в направлении, противоположном щелчку. Но нет: гребешки движутся “вперед”, как бы прогрызая путь сквозь толщу воды. Дело в том, что щелкающие движения прокачивают воду через пару отверстий, расположенных позади замка раковины. Две образующиеся струи и толкают моллюска “вперед”.
А как насчет признаков, которые эволюционировали лишь однажды или не появились вообще никогда? Мы узнали из “Рассказа Ризобии”, что колесо с настоящими свободно вращающимися подшипниками, похоже, появилось однажды – у бактерий. Позднее его изобрели люди. Язык, судя по всему, есть лишь у нас: это событие, по меньшей мере в 40 раз более редкое, чем эволюция глаза. Вообще же, привести пример хорошей идеи, которая реализовалась бы всего один раз, оказалось неожиданно трудно.
Я предложил эту задачу оксфордскому коллеге – энтомологу и натуралисту Джорджу Макгэвину. Он составил любопытный список, который, однако, оказался короче списка признаков, эволюционировавших много раз. Жуки-бомбардиры из рода Brachmus, согласно Макгэвину, уникальны тем, что смешивают химические вещества с образованием взрывчатой жидкости. Ее компоненты синтезируются в отдельных (разумеется!) железах. Когда жуку угрожает опасность, эти вещества впрыскиваются в камеру около заднего конца тела и взрываются, выстреливая во врага едкой кипящей жидкостью. Этот пример любят креационисты. Они считают, что такая способность не могла эволюционировать у жуков, поскольку все промежуточные формы должны были якобы взорваться. Я с удовольствием продемонстрировал ошибочность этого воззрения на Би-би-си в 1991 году, во время Рождественской лекции для детей Королевского института. Нацепив каску времен Второй мировой и предложив всем желающим уйти, пока не поздно, я смешал гидрохинон и перекись водорода – компоненты взрывчатой смеси жука-бомбардира. Но ничего не случилось, жидкость даже не нагрелась. Дело в том, что для этой реакции нужен катализатор. Я добавил катализатор, постепенно увеличивая концентрацию, и получил разные результаты, от тихого шипения до удовлетворительного бурления. Жук делает то же самое, и я не вижу ничего трудного в постепенном, эволюционном решении этого затруднения.
Следующий пункт в списке Макгэвина – рыба-брызгун из семейства Toxotidae. Она плюет в добычу. Рыба поднимается к поверхности воды и, выстреливая в сидящее насекомое водой изо рта, сбивает его и съедает. Другой кандидат на роль “сбивающего” хищника – муравьиный лев. Это личинки насекомых отряда сетчатокрылые (Neuroptera). Как и многие другие личинки, они не похожи на взрослых особей. У них огромные челюсти, достойные фильма ужасов. Муравьиный лев прячется под поверхностью песка, у основания конической ямки-ловушки. Эту ловушку он устраивает сам, отбрасывая песок наружу из центра: это вызывает миниатюрные оползни по краям ямки, а остальное довершают законы физики, формируя аккуратный конус. Добыча муравьиного льва (это, как правило, муравьи) падает в ямку и скатывается ему прямо в рот. Некоторое сходство с брызгуновой рыбой проявляется в том, что жертвы падают в ямку не совсем сами: иногда их сбивают в яму падающие песчинки. Хотя, конечно, песчинки не сравнить с меткими плевками рыбы-брызгуна, направленными с помощью бинокулярных глаз.
Также уникальны шипящие или плюющие пауки из семейства Scytodidae. Не обладающие проворством пауков-волков и не умеющие плести паутину, шипящие пауки выпрыскивают в сторону добычи ядовитую клейкую жидкость, которая пригвождает ее к земле. После этого паук убивает жертву укусом. Этот способ охоты отличается от тактики рыб-брызгунов. Некоторые животные, например плюющиеся ядом кобры, делают это не с охотничьими целями, а для защиты. Еще один уникальный случай – пауки-боладоры из рода Mastophora. Они мечут в добычу (ночных бабочек, привлеченных запахом поддельного полового феромона, который синтезирует паук) комочек паутины, привязанный к нити, подобно шару-бола.
Замечателен и следующий кандидат на эволюционную уникальность: паук-серебрянка (Argyroneta aquatica). Он живет и охотится под водой, но, подобно дельфинам, дюгоням, морским черепахам, пресноводным улиткам и другим наземным животным, вторично приспособившимся к жизни в воде, дышит атмосферным воздухом. В отличие от остальных “перебежчиков”, серебрянка сооружает “водолазный колокол” из паутины (которая у пауков служит универсальным решением любой проблемы) и крепит его к водорослям. На поверхности паук набирает воздух тем же способом, что и некоторые водяные клопы: воздух задерживается между волосками, покрывающими его тело. Однако дальше, в отличие от клопов, которые просто носят воздух на себе как водолазный баллон, серебрянка тащит его в “водолазный колокол”. Здесь паук ждет добычу, хранит ее и поедает.
Однако чемпион, по Макгэвину, – это личинка африканского слепня из рода Tabanus. Когда лужи, в которых живут и питаются личинки, пересыхают, личинка закапывается в грязь и окукливается. Позднее из засохшей грязи выходит слепень. Он питается кровью и в конце жизненного цикла, во время нового сезона дождей, откладывает яйца в лужи. Закопанным в грязь личинкам грозит очевидная опасность: высыхая, грязь трескается, и трещина может пройти прямо через убежище личинки. Теоретически у нее есть способ избежать этого, перенаправив трещину в обход своей камеры. Так личинка и делает. Прежде чем закопаться и окуклиться, личинка, подобно штопору, ввинчивается в грязь, а затем таким же образом прокладывает путь на поверхность, двигаясь по встречной спирали. После этого она ныряет между двумя спиралями, где и переживает тяжелые времена. Теперь она заключена в грязевый цилиндр, стенки которого предусмотрительно ослаблены двумя спиралями. Поэтому, когда трещина ползет через сохнущую грязь и достигает стенки камеры, она, вместо того чтобы пройти прямо через нее, идет по спиральному пути. Личинка спасена. Это приспособление работает по типу перфорации почтовой марки. Макгэвин считает, что эта хитрость уникальна для данного рода слепней.
А есть ли хорошие идеи, которые никогда не эволюционировали под влиянием естественного отбора? Да. Насколько я знаю, ни у одного животного на планете никогда не было органа для передачи или приема радиоволн для коммуникации. Другой пример – использование огня. Человеческий опыт показывает, насколько полезным может быть огонь. Известны растения, которым для прорастания семян нужен огонь, однако не думаю, что они пользуются им в том же смысле, в котором, например, электрические угри пользуются электричеством. Использование металла для формирования скелета – другой пример хорошей идеи, ни разу не осуществившейся на практике, за исключением предметов, изготовленных человеком. Предположительно, дело в том, что эту идею трудно реализовать в отсутствие огня.
Подобные сравнения, в которых мы подсчитываем, какие признаки эволюционируют часто, а какие – редко, удобно совмещать с географическими сравнениями. Это может позволить нам предсказать особенности внеземной жизни, а также предугадать вероятный результат мысленных экспериментов по “перезапуску” эволюции. Мы, безусловно, можем делать ставку на глаза, уши, крылья и электрические органы – но вряд ли на взрывчатку жука-бомбардира или плевки рыбы-брызгуна.
Биологи, которые, если можно так сказать, последовали примеру Стивена Джея Гулда, считают, что все этапы эволюции, включая эволюцию после кембрия, в немалой степени были основаны на случайностях, которые вряд ли можно воспроизвести в “перезапусках” эволюции по Кауфману. Гулд независимо от Кауфмана придумал мысленный эксперимент, назвав его “перемоткой эволюционной ленты”. Как правило, вероятность появления при “перезапуске” эволюции животного, хотя бы отдаленно напоминающего человека, считается исчезающе малой. Гулд убедительно показал это в книге “Чудесная жизнь”. Именно эта общепринятая точка зрения побудила меня написать осторожное самоотречение во вступительной главе. Она же заставила меня совершить наше путешествие в обратном направлении. И теперь она вынуждает покинуть моих спутников в “Кентербери” и возвращаться в одиночестве. И все же… Я думал, не зашла ли эта грозная ортодоксальность слишком далеко. В рецензии на книгу Гулда “Фулл-хаус” (Full House), перепечатанной в “Капеллане дьявола”, я защищал непопулярную идею прогресса в эволюции. Причем не прогресса в сторону человека (Дарвин меня упаси!), а в направлениях, которые, как минимум, достаточно предсказуемы для того, чтобы оправдать употребление этого слова. Ниже я покажу, что кумулятивная эволюция таких сложных адаптаций как, например, глаза, дает все основания предполагать наличие прогресса – особенно в сочетании с некоторыми замечательными плодами конвергентной эволюции.
Кембриджский геолог Саймон Конвей-Моррис в провокационной книге “Решение жизни: неизбежные люди в одинокой Вселенной” приводит аргументы, противоположные “случайности” Гулда. Значение, которое Конвей-Моррис вложил в подзаголовок своей книги, недалеко от буквального. Он действительно считает, что “перезапуск” эволюции привел бы к второму пришествию человека или кого-то очень близкого к человеку. Это непопулярное утверждение он подкрепляет вызывающе смелыми доказательствами. Свидетели, которых он неоднократно вызывает: конвергенция и ограничивающие факторы.
Сходные проблемы требуют сходных решений, и происходит это не дважды и не трижды. Я всегда считал, что был весьма категоричен в отношении конвергентной эволюции, однако в лице Конвея Морриса я встретил достойного соперника. Он приводит впечатляющий список примеров, и о многих я прежде не знал. Но если я, защищая конвергенцию, обычно прибегаю к сходным направлениям давления отбора, Конвей-Моррис добавляет к этому показания своего второго свидетеля – ограничивающие факторы. Средства, которыми располагает жизнь, и процессы эмбрионального развития допускают ограниченный диапазон решений проблемы. Для любой эволюционной отправной точки существует ограниченное число решений. Поэтому, если условия давления отбора в двух экспериментах по “перезапуску” окажутся сходными, ограничения развития заставят жизнь искать сходные решения.
Вот так ловкий адвокат мог бы использовать этих “свидетелей” в защиту смелой теории, согласно которой “перезапуск” эволюции с высокой вероятностью приведет к появлению прямоходящего существа с крупным мозгом, двумя ловкими руками, направленными вперед камерными глазами и другими человеческими признаками. К сожалению, на нашей планете это произошло лишь однажды. Но, думаю, все когда-то случается впервые.
Признаться, меня впечатлили доводы Конвея-Морриса в пользу предсказуемости эволюции насекомых. Среди определительных признаков насекомых следующие: членистый экзоскелет; фасеточные глаза; характерный способ передвижения на шести ногах, причем три всегда оказываются на земле, образуя треугольник (две ноги с одной стороны, одна с другой); дыхательные трубки-трахеи, переносящие кислород к внутренним органам через отверстия (дыхальца) по бокам тела. Завершая список эволюционных особенностей, нужно упомянуть многократное (одиннадцать раз независимо!) появление у насекомых высокоорганизованных общественных колоний, например у медоносных пчел. Вам кажется, что все это необычно? Уникально в великой лотерее жизни? Вовсе нет: эти признаки конвергентны.
Конвей-Моррис последовательно показывает, что каждый из пунктов списка неоднократно эволюционировал в разных областях царства животных, зачастую несколько раз, в том числе среди самих насекомых. А если природе настолько легко дается эволюция отдельных признаков, не так уж невероятна повторная эволюция совокупности этих признаков. Меня очень привлекает та идея Конвея-Морриса, что не стоит считать конвергентную эволюцию чудом. Возможно, мы должны считать нормой именно ее, а не удивляться исключениям. Одно из них – синтаксический язык, который, судя по всему, уникален для нашего вида. Возможно (к этому я вернусь), это единственное, чего недоставало бы повторно эволюционировавшему прямоходящему животному с крупным мозгом.
Я предупреждал: не стоит искать в эволюции паттерны, рифмы или причины. Однако я оговорился, что иногда все же буду возвращаться к этим идеям. Глава “Возвращение Хозяина” позволила мне пробежаться по эволюции в прямом направлении и посмотреть, какие паттерны мы можем найти. Идея того, что эволюция была нацелена на создание Homo sapiens, разумеется, благополучно опровергнута, и ничто из увиденного нами не смогло восстановить ее в правах. Даже Конвей-Моррис не идет дальше утверждения о том, что одним из результатов “перезапусков” может быть нечто, напоминающее человека. Другим результатом могут явиться, например, насекомые.

