Глава 9
Африка снова включается в историю человечества
Новый взгляд на нашу африканскую прародину
Когда мы все уверились, что Африка играла в истории людей центральную роль, то, как ни парадоксально, перестали обращать внимание на последние 50 тысяч лет ее истории. Изучение раннего периода – до 50 тысяч лет назад – двигалось общим желанием понять природу великого скачка к современному человеческому поведению, того самого, который знаменует переход от среднего к позднему каменному веку в африканской археологической летописи, а вне Африки – от среднего палеолита к верхнему. Но что происходило в Африке после этого перехода, ученых уже не слишком интересует. В обиходе ситуация выглядит примерно так: “мы ушли из Африки”, как будто главные герои истории современного человечества – раз! – и переместились в Евразию. Возникает впечатление (ошибочное!), что, когда Африка дала начало будущим неафриканцам, на этом ее история и закончилась, а те, кто остался на континенте, – это не более чем замершие тени прошлого, исключенные из основной сюжетной линии и неизменные на протяжении последних 50 тысяч лет.
В этом смысле поражает контраст между богатством информации о последних 50 тысячелетиях в Евразии и скудостью синхронной информации по Африке. Так, по Европе, откуда происходит львиная доля археологических материалов, задокументированы в деталях серии культурных трансформаций: от неандертальцев к преориньякцам, от них к ориньякцам, к граветтской культуре, к людям мезолитической культуры, затем к фермерам каменной индустрии, бронзовой и железной. Пропасть между нашим пониманием доисторической Евразии и доисторической Африки расширилась еще больше с наступлением революции древней ДНК, ведь материалы по древней ДНК в непропорциональном объеме евразийские, и в особенности европейские.
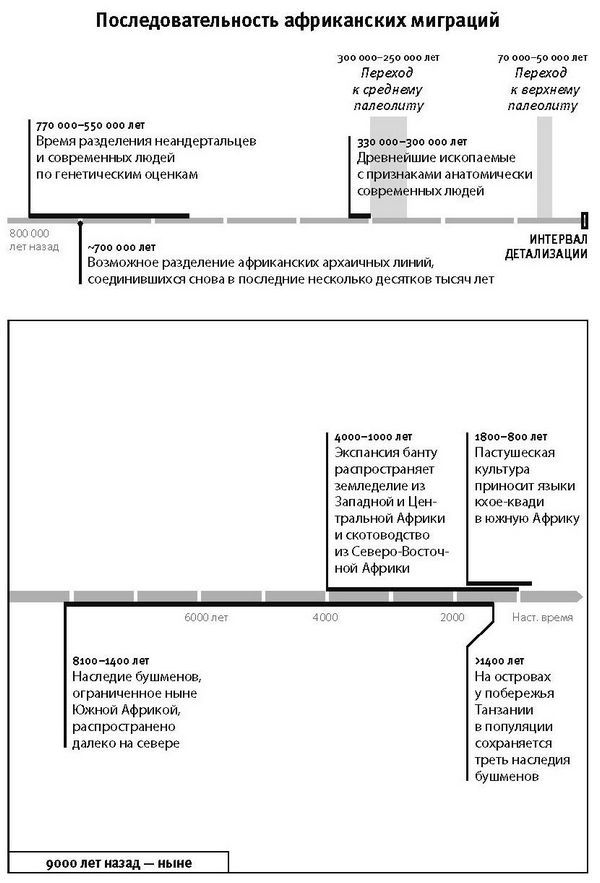
Но если копнуть поглубже, то, как показывают все изыскания на эту тему, “оставленные в африканском тылу” африканцы изменились ничуть не меньше, чем потомки ушедших выселенцев. И основная причина нашей неосведомленности об истории современных африканцев – это просто отсутствие исследований. Последние несколько десятков тысяч лет человеческой истории Африки составляют важную часть повести о нашем виде. Как ни странно, массированные исследования по проблеме происхождения нашего вида, высветившие главную роль Африки в первых актах, сослужили ей дурную службу, потому что отвлекли внимание от ее сегодняшнего населения – что оно собой представляет и как оно сложилось. А эти вопросы можно решать с помощью современной и древней ДНК.
Давнее смешение, давшее современного человека
В 2012 году Сара Тишкофф с коллегами изучали присутствие архаичных примесей в геномах современных африканцев; при этом ученые не имели доступа к архаичным геномам вроде неандертальских и денисовских, позволивших установить факт смешения архаичных и современных людей в Евразии1.
Специалисты из группы Сары Тишкофф отсеквенировали геномы представителей нескольких наиболее разнообразных африканских популяций, а потом в полученных данных попытались выявить признаки гибридизации с архаичными людьми. Предполагалось, что это должны быть относительно протяженные отрезки ДНК с высокой плотностью мутаций – выше, чем в остальных участках генома. Подобная картина гипотетически может получиться при недавней гибридизации популяции с другой группой, изолированной до сравнительно недавнего времени2. Когда такой подход применили к сегодняшним неафриканским популяциям, то те кусочки ДНК, которые отличались по изменчивости, оказались почти нацело неандертальского происхождения. И в африканских геномах, которые изучала команда Тишкофф, тоже выявились подобные выбивающиеся из общего ряда фрагменты, но вовсе не от неандертальцев унаследованные. В африканских геномах нет ничего или крайне мало от неандертальцев, а следовательно, выявленная примесь происходила от каких-то таинственных архаичных африканских людей – популяции-призрака, чьи геномы еще никогда не секвенировали.
Какое родство у этой архаичной популяции-призрака с нынешними африканцами? Этот вопрос поставили перед собой Джеффри Уолл и Майкл Хаммер, взявшись изучать тот же тип генетической изменчивости3. Они рассчитали, что та архаичная популяция отделилась от линии предков нынешнего африканского человечества 700 тысяч лет назад, а примерно 35 тысяч лет назад снова смешалась с нею, и в результате около двух процентов ДНК геномов сегодняшних африканцев составляет эта древняя примесь. Мы, однако, должны к этим процентам и датировкам относиться с осторожностью, потому что они определялись на ограниченном материале и потому что скорость мутирования у людей определяется весьма неточно.
Но выведенная возможность архаичной примеси в южноафриканских популяциях очень впечатляет, особенно если учесть, что в Западной Африке известны ископаемые люди с архаичными признаками и их датировка составляет не более 11 тысяч лет, то есть до сравнительно недавнего времени в Африке наряду с современными людьми сосуществовала и архаичная публика4. Здесь просматриваются богатые возможности для смешения расселяющихся современных людей с древними группами, примерно как это происходило в Евразии.
Если расчеты Уолла и Хаммера близки к истине и вклад архаичных линий составляет лишь 2 %, то этот генетический довесок имеет весьма скромное биологическое влияние, сопоставимое с влиянием неандертальской и денисовской примесей на нынешних неафриканцев. Но вдруг в еще более далекой африканской истории были и другие крупные перемешивания? Мы не можем исключить такой возможности. Лучше всего такие примеси выявляются по частоте мутаций, как это стало понятно на материале популяций Южной Африки.
Представим, что у человека появилась мутация. Она исключительно редкая, поскольку есть лишь у одного человека. В череде поколений частота этой мутации будет случайным образом скакать то вверх, то вниз в зависимости от числа потомков, которым она досталась. Большинство мутаций никогда не достигает значительной частоты, потому что изначально небольшое число носителей может в какой-то генерации вовсе не оставить потомства, и тогда частота данной мутации становится равной нулю, мутация сходит со сцены.
Но в популяцию постоянно закачиваются новые мутации, очевидно, поначалу редкие, и это приводит к тому, что частых (распространенных) мутаций должно быть меньше, чем редких. Соотношение частых и редких мутаций (под мутациями имеются в виду вариабельные позиции в геноме) подчиняется универсальному обратному закону: число мутаций, частота которых 10 % (которые имеются у 10 % индивидов в выборке), в два раза больше, чем тех, частота которых 20 %, а тех, в свою очередь, в два раза больше, чем с частотой 40 %.
Мой коллега Ник Паттерсон взялся проверить, как этот закон выполняется на мутациях в геномах большой выборки людей йоруба в Нигерии; он ограничился теми мутациями, которые имеются и в геномах у йоруба, и в геномах неандертальцев5. Идея с привлечением неандертальцев на редкость уместна – ведь эти мутации почти наверняка были (и имели высокую частоту встречаемости) уже в популяции общих предков людей и неандертальцев, а следовательно, и у их потомков тоже. С математической точки зрения, наше ожидание, что эти мутации должны были иметь высокую частоту встречаемости у предков йоруба, в точности уравновешивается обратным законом, так что в итоге получается, что эти мутации у йоруба должны быть равномерно распределены по всем частотам.
Но реальные данные показали иную картину. В последовательностях ДНК йоруба по результатам Паттерсона обнаруживаются два краевых максимума: на интервалах с высокой и низкой частотой доля “общих с неандертальцами” мутаций повышена, и нет никакого равномерного их распределения по всем частотам. Подобное U-образное распределение мутаций должно получаться, если имело место древнее смешение. После разделения двух популяций в каждой из них частота каждой мутации флуктуирует случайным образом. При этом некоторые мутации достигают частоты 0 % (утрачиваются), другие – 100 % (фиксируются), однако в двух популяциях такая судьба постигнет разные мутации: если в одной популяции мутация случайно достигла частоты 0 (или 100), то в другой популяции она этого, скорее всего, не сделала. И когда наши две популяции смешиваются снова, то мутации, которые в первой популяции достигли экстремальных частот, а во второй нет, в смешанной популяции окажутся где-то посередине в качестве обычных вариабельных позиций. Это должно привести к появлению двух пиков на графике распределения мутаций по частотам. Первый пик соответствует мутациям, которые в одной популяции зафиксировались, а во второй нет. Второй пик соответствует мутациям, которые приблизились к 100-процентной частоте во второй популяции, но не в первой. Частота таких мутаций в смешанной популяции (то есть положение двух пиков на графике) будет определяться той пропорцией, в которой смешались две предковые популяции. Как раз это Паттерсон и обнаружил, пояснив, что результаты укладываются в схему смешения двух популяций в равных долях.
Паттерсон на модели проверил также гипотезу, что наследниками данного смешения являются только йоруба, а неафриканское человечество не является. Но эта гипотеза не подтвердилась. Получилось, напротив, что неафриканцы вместе с самыми далеко отстоящими ветвями, такими как бушмены, все без исключения оказались потомками данной смешанной популяции. Так что хотя Паттерсон начал с западных африканцев, но событие смешения, которое он выявил, относилось не только к ним. Оно, по-видимому, сказалось на наследии всего сегодняшнего человечества. По времени это смешение должно было произойти близко ко времени, когда в скелетной летописи появились признаки современного человека, то есть около 300 тысяч лет назад6.
Заключение Паттерсона перекликается с работой Хен Ли и Ричарда Дурбина о размере предковой популяции, выведенной из характеристик единственного генома (об этой работе говорилось в первой части)7. Напомню, что сравнивались две родительские последовательности, полученные потомком соответственно от матери и от отца. В результате выяснилось, что те участки, которые попали в геном 400–150 тысяч лет назад, меньше представлены в современных геномах, чем должны быть при постоянстве размера популяции8. Одно из возможных объяснений состоит в том, что предковая популяция всех современных людей была исключительно велика на этом временном интервале, поэтому вероятность, что в двух случайно выбранных сегодняшних геномах будет наследие одного конкретного предка того времени, понижена (иными словами, в каждой генерации множество возможных предков). Но есть и другое объяснение: предковая популяция была не единой и гомогенной группой с равномерным свободным перемешиванием, а подразделялась на множество разнообразных группировок. Тогда линии, ведущие к современным людям, могли принадлежать частями к разным разделенным и изолированным группировкам. В такой ситуации картина частот мутаций по Паттерсону как раз и проявится. Что касается реконструированных временных рамок, то они соотносятся с интервалом, когда в скелетах совмещались архаичные и современные признаки. Взять, к примеру, недавно найденные в южноафриканской пещере человеческие скелеты, отнесенные к новому виду Homo naledi: у них сравнительно современное строение тела, но мозг маленький, меньше современного диапазона; датировки Homo naledi попадают между 340 и 230 тысячами лет назад9.
Есть и третье доказательство реальности древнего смешения. Обычно считается, что бушмены, ведущие образ жизни охотников-собирателей, являются наследниками особой линии, отделившейся от основного человеческого ствола раньше всех, еще до разделения остальных африканцев на отдельные ветви10. А это бы значило, что у бушменов и любой из неафриканских популяций равное число общих мутаций. Но у Понтуса Скоглунда из моей лаборатории результат получился иным. А именно что у бушменов больше общих мутаций с охотниками-собирателями из Центральной и Восточной Африки, чем с западноафриканскими популяциями, такими как нигерийские йоруба11. Картина вполне логична, если популяции Западной Африки вобрали больше наследия от одной из давно отделившихся групп, чем все остальные. По-видимому, все сегодняшние люди являются смесью двух глубоко различных предковых групп, во всех есть та или иная доля ДНК от обеих, но при этом западным африканцам досталось от одной из них больше всех.
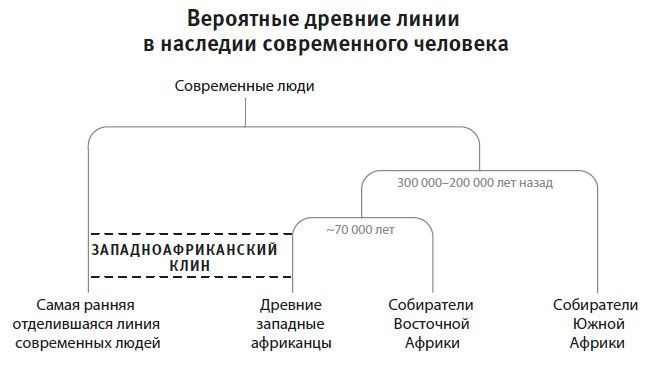
Рис. 25. Формировавшиеся в глубокой древности родственные связи между нынешними африканскими популяциями совсем не простые. Один из вариантов этих связей, который выводится из генетических закономерностей, предполагает раннее – от 300 до 200 тысячелетий назад – разделение линий современного человека в Африке; какая-то из отделившихся ветвей представлена в наибольшей степени у сегодняшних западных африканцев. Время разделения соответствует расхождению восточной и южной ветви популяций охотников-собирателей. Экспансия современных людей в период перехода к позднему каменному веку или к верхнему палеолиту, то есть позже 50 тысяч лет назад, могла снова соединить все африканские популяции.
Такие результаты позволяют предположить, что в Африке задолго до рубежа в 50 тысяч лет назад, то есть до появления симптомов современного поведения в археологической летописи, произошло крупное популяционное смешение. Этот эпизод гибридизации никак нельзя причислить к скромным по эффекту событиям, подобным 2 % неандертальской добавки у неафриканцев или такому же по размеру довеску от архаичной популяции-призрака, выведенной Уоллом и Хаммером. Потому что это не примесь, а настоящая смесь примерно в равных – 50 на 50 – долях. И нельзя сказать, какой из источников считать современным, а какой архаичным. Возможно, никакой из них не был современным, и никакой – архаичным. Может, само смешение и явилось той квинтэссенцией, которая породила современного человека: соединенные и по-новому перетасованные биологические черты двух популяций дали преимущество новообразованной общности людей.
Как сельское хозяйство спрятало прошлое Африки
С какой стороны приступать к исследованию африканского прошлого, того, которое стало разворачиваться уже после становления современного человека и его расселения за пределы Африки и Ближнего Востока, то есть позже 50 тысяч лет назад? Информации на этот счет немало, так как африканские последовательности ДНК обычно на треть более разнообразны, чем любые неафриканские. Это удивительное разнообразие свойственно не только каждой отдельной африканской популяции, но и всему африканскому человечеству, а все потому, что некоторые африканские популяции существовали изолированно от остальных в четыре раза дольше, чем любые взятые наугад пары внеафриканских популяций. Проявляется это, если, например, сравнить число разнящихся мутаций у любых индивидов в парах африканских популяций (к примеру, бушменов из Южной Африки и йоруба из Западной Африки) и в парах любых неафриканских популяций – даже минимальная разница между африканскими индивидами будет выше, чем между неафриканцами12.
Но изучать это разнообразие исключительно трудно, потому что все древние вариации – а они и в самом деле имеются в геномах нынешних людей – здорово перемешаны. Близко к современности, то есть в последние несколько тысяч лет, имели место четыре крупные экспансии, связанные с продвижением сельского хозяйства и маркированные распространением языковых групп13. Все это создало хороший генетический микс. В нем утонуло африканское прошлое, ведь экспансии двигали народы на многие тысячи километров от места происхождения, новые пришельцы замещали или перемешивались с местными группами, которые прежде тоже были широко расселены. В этом смысле изучение африканских популяций мало отличается от соответствующих евразийских историй, в Евразии население тоже многажды обновлялось.
Самое сильное влияние на африканское население относится к сельскохозяйственной экспансии банту-говорящего народа14. Археологические факты повествуют, как шло распространение новой культуры: оно стартовало примерно 4 тысячи лет назад с запада Центральной Африки, где сейчас приграничный район Нигерии и Камеруна15. Около 25 столетий назад эта культура достигла области озера Виктория на востоке Африки, принеся технологию железных орудий16, а 17 столетий назад докатилась и до Южной Африки17. В результате подавляющее большинство народов Восточной, Центральной и Южной Африки говорят на языках банту, а самое высокое языковое разнообразие приходится на сегодняшний Камерун. Это согласуется с гипотезой о существовании в районе Камеруна языка протобанту, который вместе с культурой, зародившейся там 4 тысячи лет назад, стал распространяться во все стороны18. Банту входит в более крупное объединение, так называемую нигеро-кордофанийскую языковую семью, вобравшую большинство языков Западной Африки19. Здесь, вероятно, кроется объяснение, почему рисунок генетической изменчивости (то есть частотное распределение конкретных мутаций) в группах из Нигерии и Замбии имеет больше сходства, чем в группах из Германии и Италии, хотя первая пара разделена большим расстоянием, чем вторая.
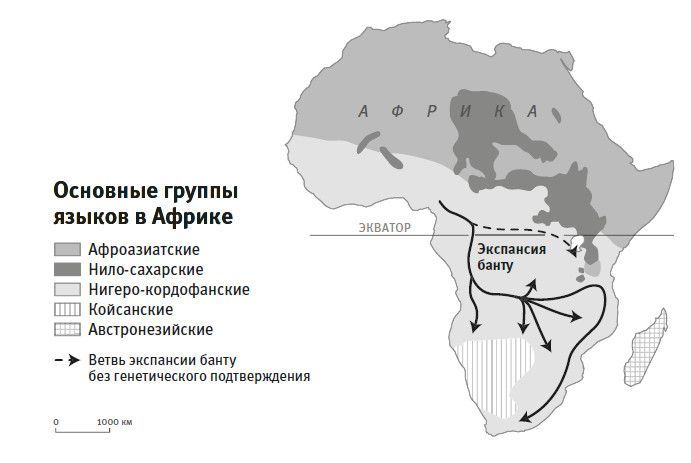
Рис. 26. Вследствие экспансии банту в последние 4 тысячи лет на сегодняшний день западноафриканское наследие доминирует в Восточной и Южной Африке.
Сейчас разработаны сверхчувствительные генетические методы, позволяющие на интервале в несколько последних тысяч лет выявлять наличие общего предка у пары индивидов. С помощью такого метода удалось проследить географический маршрут экспансии банту. Генетическая изменчивость носителей банту на востоке Африки больше схожа с картиной изменчивости в Малави к югу от центральноафриканских тропических лесов, чем с изменчивостью популяции Камеруна20. Это означает, что основная ветвь экспансии сначала двинулась на юг, а уж потом в восточные области, взяв восточный курс уже из южных форпостов. Данная схема противоречит принятой ранее теории о прямой восточной экспансии из Камеруна, которая до получения генетических данных казалась вполне обоснованной.
Другая мощная сельскохозяйственная экспансия соотносится с распространением нило-сахарских языков, на которых говорят от Мали до Танзании. Многие популяции, говорящие на этих языках, занимаются скотоводством, поэтому эту языковую экспансию рассматривают в контексте распространения скотоводства и земледелия в засушливые зоны Сахеля (саванновые полосы к югу от Сахары), когда 5 тысяч лет назад Сахара стала расширяться. Нилотские языки – одна из крупных ветвей нило-сахарских языков, их используют скотоводы долины Нила и Восточной Африки, в том числе масаи и динка. Судя по генетике, пастухи, говорившие на нилотских языках, попадая на территории местных земледельцев, не обязательно занимали социально подчиненное положение. Так, в земледельческой группе луо из Западной Кении (к которой принадлежал отец Барака Обамы) говорят на нилотском языке. Ученый из Кении, Джордж Айодо, работавший в моей лаборатории, сам луо по национальности, выяснил, что профиль генетической изменчивости у луо очень похож на таковой у носителей банту. Объясняется это, по-видимому, тем, что банту-говорящие восточноафриканские земледельцы переняли нилотский язык у высокостатусных соседей-скотоводов21.
Есть в Африке еще одна группа языков – афроазиатских, – чье происхождение совсем неясно. Сейчас они наиболее разнообразны в Эфиопии, так что идея о северо-восточном центре происхождения этой группы приобретает дополнительный вес22. Но афроазиатские языки включают еще и ближневосточную ветвь: арабский, иврит и древний аккадский. С этими языками связана интродукция ячменя, пшеницы и других культур ближневосточного земледелия на северо-восток Африки23, что происходило около 7 тысяч лет назад24. Тут очень вовремя появились данные по древней ДНК – по ним начинают проступать документальные свидетельства о миграциях людей между Северной Африкой и Ближним Востоком, а вместе с ними перемещались языки, обычаи и зерновые культуры. В 2016–2017 годах из моей лаборатории вышли две работы, в которых разбиралась генетическая общность многочисленных восточноафриканских групп, включая носителей и афроазиатских, и других языков. Выяснилось, что их общее наследие в значительной мере происходит от ближневосточной популяции, существовавшей примерно 10 тысяч лет назад25. Также мы нашли хорошее подтверждение еще одному потоку мигрантов из Западной Евразии в Восточную Африку, на сей раз связанных с иранскими фермерами бронзового века, что вполне понятно, учитывая мощные ближневосточные миграционные импульсы того периода. Эта часть ближневосточного наследия широко представлена у нынешних народов Сомали и Эфиопии, говорящих на кушитской ветви (в ранге подсемьи) афроазиатских языков. Так что по генетике вырисовываются две крупных миграционных волны, двигавшиеся с севера на юг как раз во время расползания и диверсификации афроазиатских языков. При этом не нашлось никаких генетических признаков популяций Черной Африки (области к югу от Сахары) в геномах у египтян или жителей Ближнего Востока и Египта, живших до Средних веков26. По генам, естественно, никак не определишь, на каком языке человек разговаривал, так что генетика сама по себе не является доказательством той или иной гипотезы о центре зарождения афроазиатских языков, будь то Черная Африка, Северная Африка, Аравия или Ближний Восток. Но при этом у генетиков нет сомнений, что по крайней мере некоторые афроазиатские языки принесли фермеры с Ближнего Востока. Другой вопрос: на каком языке говорили в Африке до них?
Четвертая сельскохозяйственная экспансия в Африку маркируется языками кхое-квади на юге континента. Подобно койсанским языкам охотников-собирателей Южной Африки – жу (!кунг) и таа (къхонг) – кхое-квади используют щелкающие звуки. Основываясь на сходстве пастушеского словаря, предполагается, что южноафриканские языки кхое-квади были принесены скотоводами из восточных областей примерно 18 столетий назад, а щелкающие звуки новопоселенцы могли воспринять от местных жителей27. Генетические данные согласуются с этой гипотезой: нынешние носители кхое-квади имеют значительную долю наследия восточноафриканцев. Сравнив долю эфиопского наследия в геномах носителей кхое-квади и койсанских языков жу и таа, Джозеф Пикрелл (из моей лаборатории) в 2012 году пришел к выводу, что у кхое-квади оно непропорционально высокое. И что это, если не результат миграции с севера?28 По размеру фрагментов ДНК восточноафриканского происхождения в геномах кхое-квади можно рассчитать время этого генетического смешения; оно примерно соответствует прибытию в южные области 8–9 тысяч лет назад некоей гипотетической популяции скотоводов и последующей паузе перед полным перемешиванием ее с местной популяцией. В этих восточноафриканских фрагментах Пикрелл обнаружил вставки, сходные по своей специфике скорее с ближневосточной популяцией, чем с какой бы то ни было другой. Судя по длине вставок, они попали в геном около трех тысяч лет назад. Как раз в это время в эфиопских областях шло смешивание людей западноевразийского происхождения с народами, несущими наследие Черной Африки29. Так что и эта деталь подтверждает восточноафриканское происхождение кхое-квади.
В 2017 году древняя ДНК подтвердила восточноафриканскую гипотезу. Понтус Скоглунд анализировал древнюю ДНК из скелетов, датированных возрастом приблизительно 31 столетие назад. Это были останки девочки-младенца из Танзании в экваториальной Восточной Африке и двенадцатилетнего подростка из Западно-Капской провинции в ЮАР. Оба были найдены в погребениях с артефактами и костями животных, указывающих на скотоводческие группы30. Танзанийская девочка, как оказалось, происходила из той гипотетической популяции-призрака, предсказанной нами с Пикреллом: из группы с большой долей наследия древних восточноазиатских охотников-собирателей и оставшейся долей с западноевразийской спецификой. И с этой популяцией практически наверняка связано распространение скотоводства из ближневосточных регионов на север Африки и дальше, южнее Сахары. В древней ДНК девочки из Танзании одна треть была от той пастушеской популяции, а две трети от местных групп, близких по генетике к сегодняшним бушменам. А у двенадцатилетнего подростка из Южной Африки геном представлял смесь, такую же, как у скотоводов, говорящих на кхое-квади. Так что все указывает на то, что и ранние языки кхое-квади, и пастушеское хозяйствование, и восточноафриканское генетическое наследие – все перемещалось в южном направлении с миграциями людей.
Перед нами ландшафт биологического и культурного разнообразия сегодняшней Африки, сформированный в ходе сельскохозяйственных экспансий последних нескольких тысячелетий, – он фантастичен. Но в то же время за этой фантастичностью трудно увидеть и понять общую картину событий. В ловушку сегодняшнего впечатляющего разнообразия раз за разом попадают и лингвисты, и археологи – словом, все, кто приступает к изучению Африки; эффект только усиливается, когда перед глазами проходит вереница удивительно непохожих друг на друга африканских лиц, ведь Африка обычно представляется нам совсем другой. Начинаешь думать, что глубокое прошлое Африки не понять, не объяснив всё-всё это разнообразие. И забываешь при этом, что большая часть популяционной структуры сегодняшней Африки сложилась под влиянием сельскохозяйственных экспансий последних нескольких тысяч лет. Поэтому, сосредоточившись на описании гипнотического африканского разнообразия, упускаешь из вида более масштабную, дальнюю, перспективу формирования африканского населения. Так что это разнообразие, как и происхождение современных людей, оказывает Африке в каком-то смысле дурную услугу. Нужно перестать описывать это пестрое верхнее покрывало, снять его совсем – и в этом нам поможет древняя ДНК.
Ищем древних африканских охотников-собирателей
Что за люди жили в Африке до эпохи производительного хозяйства, радикально преобразившего человеческий ландшафт континента? Если основываться на теперешнем африканском разнообразии, то искать ответ на этот вопрос очень и очень непросто. Здесь во введении я рассказал, как Лука Кавалли-Сфорца в 1960 году поставил на то, что по генетической изменчивости сегодняшнего населения можно реконструировать далекую историю популяций31. Но это пари он проиграл, потому что, как выяснилось из исследований древней ДНК, миграции и вымирания популяций были обычнейшим явлением, так что в подавляющем большинстве случаев даже с помощью самой изощренной статистики невозможно по современной канве восстановить картину прошлых демографических событий.
Думаю, читателя не удивит путь, позволивший преодолеть эту преграду. Этот путь – изучение полногеномных данных по древней ДНК в совокупности с данными по нынешним изолированным группам африканцев. Изолированным и в культурном, и в генетическом отношении. К ним относятся, в частности, пигмеи Центральной Африки, бушмены с южной оконечности Африки и хадза из Танзании – всё это носители “щелкающих” языков, кардинально отличающихся от окружающих их языков банту. По генетике группы со щелкающими языками тоже представляют четкий отдельный кластер. Некоторые из этих популяций содержат линии наследия, даже близко несхожие с наследием их соседей, то есть давно с ними разошедшиеся. Мы можем взять генетические данные по этим популяциям и по результатам сравнения постараться вывести события, находящиеся за пределами того, что доступно при анализе ДНК африканских популяций.
До недавнего времени было чрезвычайно трудно добыть древнюю ДНК из образцов с большинства африканских территорий. Дело в африканском климате, влажном и жарком, – в таких условиях ДНК очень быстро разрушается. Но в 2015 году революция древней ДНК докатилась наконец и до Африки: тут помогло увеличение эффективности методов выделения ДНК, а также открытие, что в некоторых костях ДНК сохраняется существенно лучше, чем в других.
Самый первый африканский образец, из которого удалось выделить пригодную для прочтения ДНК, поступил из высокогорной пещеры в Эфиопии, где возраст вмещающих скелетные остатки слоев составлял 45 столетий32. Этот древний индивид оказался родственен одной из сегодняшних эфиопских групп, ари, притом что с другими эфиопскими группами его генетическое родство заметно меньше. В сегодняшней Эфиопии жизнь людей подчиняется правилам изощренной кастовой системы, в рамках которой практикуются строгие ограничения браков между людьми с различными традиционными функциями33. Группа ари включает три подгруппы – агрономы, кузнецы и гончары, и все они отличаются по генетике и друг от друга, и от других не-ари34. Так как те 4500-летние останки похожи по генетическим признакам на ари, становится ясно, что все это время особого перемешивания внутри региона не было, а, напротив, сохранялись строгие местные барьеры, затруднявшие свободную циркуляцию генов. Лучшего примера эндогамии я не знаю – даже в Индии, насколько известно, эндогамия уходит не дальше двух тысячелетий в прошлое35.
Сюрпризы древней ДНК на этом не заканчиваются. В 2017 году Понтус Скоглунд работал с образцами 16 индивидов из Африки: останками охотников-собирателей и скотоводов с территории ЮАР с датировками от 21 до 12 столетий назад, а также охотников-собирателей из района Малави возрастом 81 и 25 столетий; вдобавок были еще останки охотников-собирателей, фермеров и скотоводов из Танзании и Кении, живших от 3100 до 400 лет назад 36 . Все они относительно недавние по сравнению с древнейшими евразийскими образцами ДНК, однако и они дают возможность заглянуть в те африканские времена, когда производительное хозяйство еще не трансформировало популяционную структуру Африки.
Неожиданно для нас из анализа древней ДНК проступили контуры популяции-призрака, доминировавшей в восточной части Черной Африки, а затем практически стертой приходом аграриев37. Мы назвали ее “восточноафриканские фуражиры”: ее наследие целиком вошло в два древних генома охотников-собирателей из Эфиопии и Кении (они имелись в нашей базе данных). Также значительная часть этого наследия обнаружилась у нынешних хадза из Танзании, число которых в настоящее время едва достигает тысячи. Кроме того, выяснилось, что восточноафриканские фуражиры имеют больше родственного с нынешними неафриканцами, чем с любыми сегодняшними группами Черной Африки. Логично предположить, что предки этой популяции-призрака могли существовать во время перехода от среднего к позднему каменному веку, то есть с них могло начаться движение из Африки в Евразию 50 тысяч лет назад. Так что популяция восточноафриканских фуражиров, возможно, играла весьма знаменательную роль в нашей истории.
Сами по себе фуражиры Восточной Африки тоже не были равномерно перемешаны. Мы это знаем потому, что наши данные по восточноафриканским фуражирам разгруппировались в три кластера, представляющих разные субпопуляции. Первая – древние кенийцы и эфиопцы, вторая – древние собиратели Занзибарского архипелага и Малави, а третья – нынешние хадза38. На основе имевшихся данных, рассыпанных по времени и пространству, мы не могли реконструировать время размежевания этих субпопуляций. Но, судя по повсеместному присутствию людей в данном регионе и значительной временной протяженности их существования в одних и тех же местах, было бы неудивительно, если бы различия между субпопуляциями начали формироваться очень давно, десятки тысяч лет назад. Есть и другой пример подобного размежевания. В 2012 году в моей, а также и в других лабораториях изучали популяцию, которую я про себя именовал “южноафриканскими фуражирами”. Эта линия отстоит от восточноафриканских фуражиров настолько же, насколько и любая другая из сегодняшних человеческих групп. Так вот, эти южноафриканские фуражиры подразделяются на две отчетливые субпопуляции, разошедшиеся по крайней мере 20 тысяч лет назад39. В Восточной Африке условия обитания человека не менее разнообразны, чем в Южной, поэтому здесь, наверное, можно ожидать столь же древнего, а может, и более древнего разделения на подгруппы.
Результаты Понтуса Скоглунда преподнесли нам еще один сюрприз. В некоторых образцах древних африканских собирателей обнаружилось наследие и южноафриканских, и восточноафриканских фуражиров. На сегодняшний день наследие линий южноафриканских фуражиров можно обнаружить лишь у населения самой южной части Африки, говорящего на “щелкающих” языках, а в полном объеме оно присутствует только у бушменов. При этом геномы древних охотников-собирателей Южной Африки представлены целиком линией южноафриканских фуражиров.
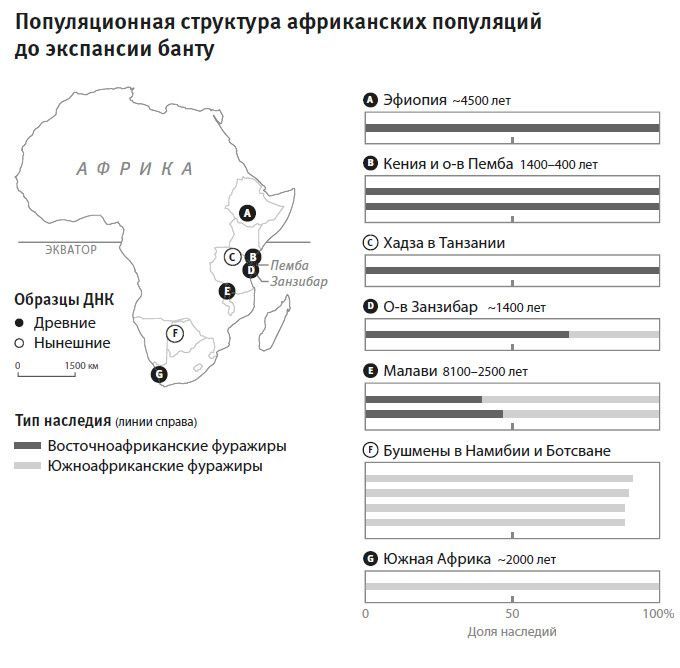
Рис. 27. Южноафриканский тип наследия сейчас в полном объеме присутствует лишь у бушменов из Южной Африки (F), но раньше он был распространен и в Восточной Африке, по крайней мере на территории Танзании. Восточноафриканский тип наследия, тот, что составляет геномы хадза в Танзании (С), раньше тоже был распространен существенно шире.
Сам термин “южноафриканские фуражиры”, как свидетельствуют наши древние образцы, может неверно указывать на место зарождения популяции. У нас есть два образца возрастом около 14 столетий, найденные на островах Занзибар и Пемба к востоку от берегов Танзании. Острова эти входят в островную гряду, которая в процессе повышения уровня моря отделилась от материка около 10 тысяч лет назад и, по-видимому, стала тогда изолированным домом для потомков охотников-собирателей Восточной Африки40. В двух упомянутых образцах геномы представляли смесь наследий: треть принадлежала южноафриканским фуражирам, а остальные две трети – восточноафриканским фуражирам41. В другой серии – 7 образцов из трех местонахождений в Малави – соотношение наследий оказалось обратным: треть восточноафриканского, две трети южноафриканского. Эта серия с юга Центральной Африки датируется между 81 и 25 столетиями назад и представляет выборку большой гомогенной популяции с характерной смесью наследий. Похоже, что южноафриканские фуражиры в прошлом были распространены на обширных территориях африканского континета, поэтому очень трудно понять, где именно сформировалась эта популяция.
Древняя ДНК преподала нам урок: история нынешней Африки уходит корнями в ушедшие популяции, в то, как они подразделялись и смешивались еще до прихода сельского хозяйства. Мы видим, что история африканского человечества сложна на всех временных срезах и на всех уровнях. Так и должно быть, учитывая колоссальные территории и разнообразие ландшафтов Африки и саму древность африканского человечества. Революция древней ДНК только начинает поглядывать на этот континент. В ближайшее время он полностью включится в революционный процесс, оттуда потечет все больше и больше данных из самых разных местонахождений и самых разных временных слоев. И эти данные наверняка изменят и прояснят наши представления о глубоком африканском прошлом.
Что еще нужно для понимания африканского прошлого
Сложность африканской популяционной структуры замечательно видна в картинах естественного отбора населения континента. Возьмем пример с серповидно-клеточной анемией: при этой болезни из-за мутации меняется структура гемоглобина, одного из белков крови, больше всех других молекул контролирующего транспорт кислорода по телу. Люди с западноафриканским наследием ДНК имеют высокий риск этого заболевания. Под давлением естественного отбора мутация серповидно-клеточной анемии достигла значительной частоты среди населения Западной (например, в Сенегале), Западно-Центральной (в частности, в Нигерии) и Центральной Африки (откуда мутация с экспансией банту распространилась по Восточной и Южной Африке). Эта мутация, если она имеется лишь в одной копии, то есть унаследована только от одного из родителей, защищает от малярии – поэтому она и достигла такой высокой частоты в тех регионах. Малярия там настолько опасна, что для популяции в целом оказывается важнее выигрыш выживания, который она дает 20-ти процентам носителей одной копии мутантного гена, чем ранняя смерть (без лечения) от серповидно-клеточной анемии одного процента носителей двух копий этой мутации. Но, что замечательно, данная мутация появилась независимо в трех регионах (западном, западно-центральном и центральном). Мы это знаем, потому что она располагается на разных участках последовательности ДНК. Неподготовленного человека это удивит, потому что раз мутация такая полезная для носителей, то, лишь появившись, она должна была бы, подгоняемая естественным отбором, быстро распространиться по обширной малярийной зоне Африки при условии даже очень редкого скрещивания с соседями42. Такую же картину мы видим для мутации лактазного гена, позволяющей переваривать коровье молоко во взрослом возрасте. Генетический базис для лактазной персистенции у северных африканцев и фульбе из Западной Африки отличается от такового у масаев Судана и Кении: тут совершенно разные мутации, хотя и в одном гене43.
Согласно заключению Питера Ральфа и Грэма Купа, скорость миграции людей по этим регионам была чрезвычайно низкой (что верно даже для областей Черной Африки, удаленных друг от друга не более чем на две тысячи километров). Иначе как бы успели независимо возникнуть мутации и серповидно-клеточной анемии, и лактазной персистенции? Поэтому здесь наиболее эффективным способом распространить полезные мутации оказалось скорее их изобретение, чем импорт из других популяций44. В Африке за последние несколько тысяч лет из-за ограниченной скорости перемешивания некоторых популяций сложилась особая популяционная структура – Ральф и Куп назвали ее “сотообразной”. Этот термин пришел из математики, и обозначает он поверхность, плотно уложенную плитками; иными словами, это генетически гомогенные площади, разделенные четкими границами. Примерно такая картина и должна получиться, если процесс генетического выравнивания за счет взаимоперемешивания соседей конкурирует с процессом появления новых полезных генетических вариантов в каждой из групп. В этом случае размеры областей распространения конкретных мутаций серповидно-клеточной анемии и лактазной персистенции будут отражать скорость генетического обмена между соседними популяциями, какой она была в последние несколько тысяч лет.
Мы преодолели лишь начальные этапы постижения африканской демографической истории, но уже ясно, насколько она сложна: тут и дробление крупных популяционных линий, таких как восточно- и южноафриканские фуражиры, имевшее место далеко в прошлом, тут и многослойные перемешивания, которые перекрыты сравнительно недавними смешениями, вызванными распространением сельского хозяйства. Собрав побольше образцов древней ДНК, мы сможем лучше осмыслить диапазон вариабельности в африканских популяциях последних нескольких десятков тысячелетий и на этой основе построить содержательную модель популяционной структуры.
При этом нет никаких сомнений, что модель эволюционного дерева африканского человечества, как, впрочем, и любого другого региона, нежизнеспособна – откуда бы мы ни брали древнюю ДНК, нигде нет такого, что веточки народов, отделившиеся некогда от общего ствола, до сих пор живут и здравствуют. Вместо этого мы видим картину множественных циклов с популяционным размежеванием и обратным смешением. Кроме того, мы можем быть уверены, что данные древней африканской ДНК опровергнут многие из устоявшихся гипотез, как это происходит с исследованиями других регионов. Что же до того, как новое представление о популяционной сложности приложить к жизни, а также к переосмыслению себя как вида, – этому посвящена следующая часть книги.

