К физиологии гипнотического состояния собаки67
Совместно с д-ром М. К. Петровой
Кроме шаблонного исторического способа гипнотизации животных (опрокидывания на спину и удерживания в этом неестественном положении некоторое время), – гипнотизации, проявляющейся в каталептическом состоянии, наши лаборатории при изучении нормальной деятельности верхних отделов головного мозга получили возможность изучать более подробно разнообразные и очень тонкие проявления гипнотического состояния. Как уже нами установлено, основным условием наступления этого состояния является продолжительность одних и тех же раздражений, приводящих, наконец, соответствующие клетки коры в тормозное состояние, которое, с одной стороны, представляет различные степени напряженности, с другой – различный размер распространенности по коре больших полушарий и дальше вниз по головному мозгу. Относящиеся сюда факты приведены в книге одного из нас (И.П. Павлов. Лекции о работе больших полушарий головного мозга).
Но дальнейшие наблюдения открывают все большее разнообразие симптомов гипнотического состояния, все более тонкие градации его, часто едва отличающиеся от бодрого состояния; все большую подвижность гипнотического состояния в зависимости от мельчайших изменений обстановки, от ничтожной смены действующих на животное внешних раздражений.
В настоящей статье мы остановимся на тех явлениях, которые нам пришлось наблюдать на двух собаках, прежде служивших одному из нас (М.К. Петровой) при изучении условных рефлексов на разные темы, а теперь постоянно приходящих в гипнотическое состояние, как только они ставятся и снаряжаются в нашей обычной экспериментальной обстановке.
Давно и часто сообщалось в работах из наших лабораторий о разъединении, в случае условных пищевых рефлексов, слюнной секреции и пищевой двигательной реакции, когда собака приходит в сонливое состояние. Дело именно происходило таким образом, что или уже и при наших искусственных условных раздражителях, или, что чаще, при натуральном (как доказано, тоже условном) раздражении видом и запахом подаваемой еды слюна обильно начинает течь, а животное еду не берет.
Вот при этом состоянии животных в наших теперешних наблюдениях и выступили очень разнообразные и в высшей степени интересные вариации пищевой двигательной реакции. Те или другие вариации эти, очевидно, как разные степени интенсивности гипнотизации обнаруживаются преимущественно то у одного, то у другого животного. У одной собаки, менее глубоко гипнотизирующейся, резко проявляется только то, что у душевнобольных называется негативизмом. После условного раздражителя, продолжающегося некоторое время, мы подаем собаке еду – она от нее отворачивается, мы отводим от собаки кормушку с едой – она к ней тянется. Мы снова подаем – опять отвертывается, отводим – снова тянется. Реакцию устранения от кормушки мы называем отрицательной или первой фазой негативизма, движение по направлению к кормушке – положительной или второй фазой. И такой негативизм может повторяться много раз, пока собака, наконец, в большинстве случаев не возьмет еду. Степень гипнотизации и выражается в числе повторений этой процедуры. При начинающейся гипнотизации еда берется и съедается при второй подаче. При углублении гипнотизации обе фазы негативизма повторяются все большее число раз. При высшей степени гипнотизации собака еду не берет, сколько бы мы ни повторяли предложение ее. Но стоит только тем или другим способом рассеять гипноз, – снятием ли приборчика, прикрепленного на собаке для собирания слюны, или отвязыванием цепи, на которой водится собака и которая на время опыта завертывается около верхней перекладины станка, или еще каким-либо способом, – чтобы собака сейчас же с жадностью начала пожирать ту же еду.
У другой собаки пищевая двигательная реакция во время гипнотизации представляется в еще более сложном виде. У нее явления, если взять более полный случай, выступают в следующем порядке. При действии наших условных раздражителей (обыкновенно к концу их изолированного действия) собака, если сидела – встает, если стояла – поворачивается всем туловищем в ту сторону, откуда ей подается еда, но при подаче еды двигает головой в сторону или вверх от нее, т. е. наступает первая фаза негативизма. Если теперь кормушка отводится от животного, то, наоборот, голова делает движение в сторону кормушки, следит за кормушкой, т. е. выступает вторая фаза. После нескольких повторений такого негативизма собака, наконец, держит пасть прямо над едой, а пищу не берет, не может ее взять. Она начинает как бы с большим трудом, понемножку и многократно, открывать и закрывать рот, но впустую, не забирая еды (абортивные движения). Затем начинает двигать челюстями все свободнее. Теперь берет еду, но маленькими порциями, и, наконец, хватает ее широко раскрытой пастью и быстро, раз за разом. Таким образом, в этой фазе гипнотизации мы должны различать три разных состояния в трех отделах скелетной мускулатуры, относящейся к акту еды: сильное торможение, скованность ближайшей: к акту еды мускулатуры (жевательных мышц и языка), значительную подвижность, но в виде периодической деятельности, в форме негативизма шейной мускулатуры, и, наконец, нормальную деятельность остальной, туловищной мускулатуры. Чем глубже гипнотизация, тем скованнее, тем более заторможена ближайшая мускулатура: язык высовывается изо рта как парализованный, а челюсти остаются совершенно без движения. На шейной мускулатуре негативизм обнаруживается только в одной первой отрицательной фазе. Затем дальше прекращаются и совсем движения головы, остается только поворот туловища при условных раздражителях. Наконец, при еще более далеко зашедшем гипнозе прекращается и эта последняя двигательная реакция на них и на еду. Все эти явления разом можно рассеять, уничтожить теми же мерами, что были упомянуты при описании первой собаки.
Относительно пищевой двигательной реакции в наших случаях надо прибавить еще следующее. Малейшие изменения в обычном виде еды или даже в способе подачи еды делают то, что отрицательная двигательная реакция сейчас же превращается в положительную, т. е. собака берет только что перед этим отвергаемую еду. Мы подаем нашей собаке еду в обычной чашке с ровно лежащим в ней несколько сыроватым мясо-сухарным порошком. Она еду не берет. Стоит порошок в той же чашке отчасти собрать большим выстоящим куском, как собака его даже жадно хватает, а затем начинает есть и остальной порошок. Также достаточно положить тот же порошок на тарелочку или на бумажку и так подать, чтобы опять получилась положительная реакция. Собака возьмет еду и из руки, вместо чашки, и, наконец, иногда она, после нашего условного раздражителя, отказываясь от порошка в чашке, начинает подлизывать тот же порошок, рассыпанный на полу станка.
Кроме описанных двигательных явлений, относящихся к акту еды, во время наших наблюдений над гипнотическим состоянием выступают и другие двигательные явления особого рода, заслуживающие внимания. Многие собаки в бодром состоянии после того, как съедят небольшую экспериментальную порцию пищи, некоторое время лижут переднюю лапу и переднюю часть груди. У собак в гипнотическом состоянии это лизание обыкновенно очень затягивается, а у одной из наших, теперь описываемых собак оно скоро принимает и некоторую особенную форму. Облизав, ослюнявив лапу, специально мякоть пальцев передней лапы, собака подносит ее к приборчику, который прикреплен (приклеен) на месте слюнной фистулы, и несколько раз проводит по нему пальцами, проделывая эту процедуру много раз, если ей не мешать в этом. В бодром состоянии та же собака этого не делала. В бодром состоянии некоторые собаки борются с этим приборчиком только тогда, когда он впервые прикрепляется, а затем с ним свыкаются, совершенно его игнорируя. Можно с правом догадываться, что у нашей собаки в гипнотическом состоянии это есть обнаружение одного из специальных оборонительных рефлексов. Когда собака имеет поранение на месте кожи, доступном ее языку, она обыкновенно повторно очищает его слюной, как говорится, зализывает рану (терапевтический рефлекс). Очевидно, в данном случае раздражение застывшей замазкой, которой прикрепляется приборчик к коже, и дает повод к этому рефлексу, причем ввиду местонахождения раздражения язык заменяется пальцами лапы.
Многие из описанных вариаций пищевой двигательной реакции обыкновенно наблюдаются разом в одном и том же опыте, быстро сменяясь одни на другие. Эта текучесть, подвижность гипнотического состояния дает себя знать и в других явлениях помимо указанных. Мы и приведем еще несколько случаев колебания гипнотического состояния и изменения эффекта условного раздражения, которые или уже были описаны ранее и повторились у нас, или же впервые отмечены нами при наблюдениях и опытах над нашими собаками. Эти колебания и изменения происходят или от еще неизвестных причин, или приурочиваются к определенным условиям.
Укажем опять, что раз собака начала гипнотизироваться в экспериментальной обстановке, то гипнотическое состояние обычно наступает сейчас же, как только собака помещается в станок, и иногда даже как только переступает порог экспериментальной комнаты, а затем оно постоянно и постепенно нарастает по мере продолжения опыта, если не вводятся условия, его рассеивающие.
Прежде всего остановимся на факте разъединения секреторной и двигательной реакций пищевого рефлекса. Это разъединение принимает часто вид как бы взаимного антагонизма. То при раздражении наступает слюноотделение при отсутствующей двигательной реакции, т. е. собака не берет еду, как указано выше, то, наоборот, собака быстро берет и жадно ест подаваемую еду, а на хорошо выработанные условные раздражители слюна не течет.
Вот пример этого. У одной из наших собак, Бека, два дня подряд так начинается опыт.
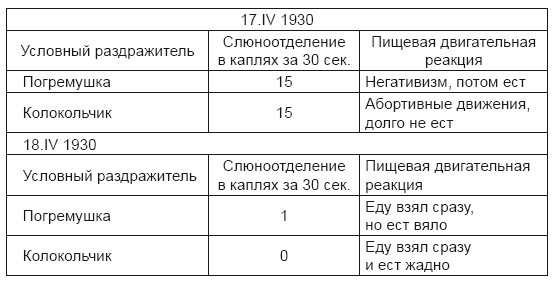
Иногда эти, как бы антагонистические, отношения между секреторной и двигательной пищевой реакциями быстро изменяются в противоположные в течение опыта.
Вот пример на другой нашей собаке «Джон».

В ранних работах из наших лабораторий много раз сообщалось, что хорошо выработанный тормозной раздражитель, обыкновенно дифференцировочный, может изменять данное гипнотическое состояние в противоположных направлениях, то усиливая, то ослабляя его. То же часто видели и мы на описываемых гипнотизирующихся животных.
Наконец, следует упомянуть о факте, что среди наших обыкновенных условных сильных раздражителей чрезвычайно сильный, условный раздражитель часто устраняет или ослабляет гипнотическое состояние, когда обыкновенные сильные раздражители или оставляют его без изменения, или даже его усиливают.
Вот пример этого, из того же опыта на Беке, начало которого было приведено выше. При продолжении опыта и после применения дифференцировки умеренно сильные условные раздражители – погремушка, бульканье и колокольчик – оставались без секреторного эффекта, п собака при подаче еды долго не брала ее при абортивных жевательных движениях. Трещотка – очень сильный условный раздражитель – вызвала слюноотделение, и собака взяла еду после короткого негативизма.
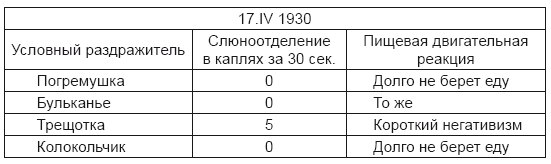
Как понять, представить себе физиологический механизм приведенных фактов? Конечно, при настоящем состоянии наших сведений о физиологии высших отделов головного мозга было бы слишком большой, не соответствующей положению дела претензией дать вполне обоснованный и ясный ответ на все при этом возникающие вопросы. Но попытки сведения частных явлений на более общие черты деятельности этих отделов должны быть постоянно делаемы в видах осуществления новых вариаций опытов с надеждой все большего приближения к пониманию чрезвычайно сложных в данном случае отношений действительности.
Существеннейшая трудность при попытках объяснить механизм перечисленных фактов гипнотического состояния заключается в том, что при раздражениях, прикладывающихся несомненно к клеткам больших полушарий, мы часто не знаем, что в происшедшей нервной деятельности надо отнести насчет больших полушарий и что насчет низших инстанций, низших отделов головного и даже спинного мозга. По мере филогенетического развития центральной нервной системы нервные комбинационные системы в виде определенных и все более усложняющихся так называемых рефлекторных центров переносились все ближе к головному концу, представляя все больший анализ и синтез возбуждающих агентов в связи с нарастанием сложности организма и умножением отношений организма с внешней средой все в больших ее районах. Таким образом, постепенно, рядом с более или менее стереотипной нервной деятельностью, с готовыми комплексами физиологических функций, вызываемыми элементарными и немногочисленными раздражениями, развивалась высшая нервная деятельность, считающаяся все с большим числом условий, большим числом уже комплексных раздражений, притом еще колеблющихся. Тогда возникает для исследования очень сложный вопрос о связи и виде связи этих разных этажей. Говоря по отношению к нашему первому вопросу о разъединении секреторной и двигательной реакций нашего условного пищевого рефлекса, надо решить, что при этом рефлексе следует отнести насчет коры и что насчет ближайшей подкорки, или, выражаясь иначе, более обыденно, что в этом процессе произвольное и что рефлекторное. Подходя еще ближе к делу, нужно знать, одинаково ли в условном пищевом рефлексе секреторный и двигательный компоненты его зависят от коры или есть разница по отношению к этим компонентам; не зависит ли двигательный преимущественно от коры, а секреторный от подкорки?
Обратимся к известным фактам.
На основании фактов человеческого гипноза необходимо признать, что в коре вместе с грандиозным представительством внешнего мира через афферентные волокна (а это есть необходимое условие высшего регулирования функций) имеется также и широкое представительство внутреннего мира организма, т. е. состояний, работы массы органов и тканей, массы внутренних органических процессов. В этом отношении особенно убедительны фактические подробности так называемой мнимой самовнушенной беременности. Масса процессов, относящихся до деятельности таких пассивных тканей, как жировая, возникают, усиливаются под влиянием полушарий. Но ясно, что по степени эти два рода представительства очень, чрезвычайно разнятся между собой. В то время как представительство скелетно-мышечного аппарата в высшей степени тонко и подробно, может быть, равно в этих отношениях представительству внешних энергий, как звуковая и световая, представительство других внутренних процессов отстает чрезвычайно резко. Может быть, это зависит и от малой жизненной практики этого последнего представительства. Во всяком случае это – постоянный физиологический факт. На этом основании, очевидно, различаются произвольные и непроизвольные функции организма, причем к первым причисляется только деятельность скелетной мускулатуры. Произвольность эта значит, что работа скелетной мускулатуры на первом плане определяется ее корковым представительством, двигательной областью коры (двигательным анализатором но нашей терминологии), непосредственно связанной со всеми внешними анализаторами, т. е. постоянно в ее направлениях определяется аналитической и синтетической работой этих анализаторов.
Исходя из этих данных, механизм образования нашего условного пищевого рефлекса надо представлять себе следующим образом. С одной стороны, это есть соединение корковых пунктов приложения условных раздражителей с рефлекторным пищевым центром ближайшей подкорки со всеми его частными функциями, с другой – более близкая связь тех же пунктов с соответствующими, т. е. с участвующими в акте еды отделами двигательного анализатора. Тогда разъединение во время гипнотизации в пищевом акте секреторного компонента от двигательного можно было бы понимать так. В силу данной гипнотизации состояние коры таково: двигательный анализатор заторможен, все остальные – свободны. С последних разрешается рефлекс на пищевой центр подкорки со всеми его функциями, а торможение двигательного анализатора, так сказать но его прямому проводу, выключает из этого рефлекса двигательный компонент, обусловливая недеятельность в последней инстанции движения, в клетках передних рогов, и таким образом от пищевого акта остается только видимая секреторная реакция.
Теперь обратный случай: на искусственный условный раздражитель слюна не течет, а двигательная реакция есть, собака подаваемую еду ест сразу. Теперь объяснение просто. Это было бы небольшое задержание всей коры, так что одного искусственного раздражения недостает для преодоления существующего торможения, и только при подаче еды, когда с искусственным условным раздражителем суммируются натуральные (вид и запах еды, а эти последние даже сами по себе сильнее искусственных), наступает полный рефлекс с обоими компонентами.
Но есть еще случай, встречавшийся в других опытах из наших лабораторий и притом наступавший вне гипнотического состояния, но который уместно анализировать в связи с теперешними объяснениями. Собака ест пищу, а слюна не течет 10 и 20 секунд. Это несомненно связано с развитием нарочного торможения в коре при помощи искусственных условных раздражителей на определенные сроки времени. Как этот случай понять? Каков его механизм? Надо представить, что с пунктов искусственных условных раздражителей развивается сильное торможение на весь подкорковый пищевой центр с его обоими главными компонентами – секреторным и двигательным, а также и на соответствующий отдел коркового двигательного анализатора. При подаче еды с пунктов более сильных натуральных условных раздражителей, которые к тому же не участвовали в развитии торможения, происходит быстрое раздражение пищевого отдела двигательного анализатора, как более подвижного сравнительно с подкорковым центром, когда в этом последнем торможение рассеивается только при более длительном действии безусловного раздражителя. Может быть, это можно было бы хотя бы отчасти аналогировать с нарочным, волевым введением пищи в рот, жеванием и глотанием ее, когда не чувствуется ни малейшего аппетита.
Но, конечно, можно принимать (и для этого у нас есть веские основания), что условная связь со слюноотделением происходит также в коре, через корковое представительство слюнных желез, и тогда все случаи разъединения секреторной и двигательной реакций сведутся на разнообразные локализации торможения при наступлении и распространении гипнотического состояния.
Следующее гипнотическое явление, физиологическим механизмом которого мы должны заняться, есть негативизм. Конечно, он есть явление торможения, потому что представляет фазовое явление, которое постепенно кончается сном. Также нет сомнения, что это – корковое локализованное торможение, потому что одновременно существующей слюнной реакцией демонстрируется условная, т.с. корковая, деятельность. Тогда естественно заключить, что это – двигательное торможение, относящееся к двигательной области коры, к двигательному анализатору. Но как понимать форму этого торможения? Почему сперва существует отрицательная фаза двигательного акта, а затем положительная? Что это за смена? Нам кажется, что это без натяжки может быть сведено на более общие, уже ранее нам известные факты. Когда начинается гипнотическое состояние, состояние торможения, корковые клетки как бы приходят в более слабое, менее работоспособное состояние, для них понижается предел допустимой, возможной возбудимости. Это так называемая парадоксальная фаза. Тогда обыкновенно сильный раздражитель делается сверхсильным и может вызывать не возбуждение, а торможение, усиливать торможение. Кроме того, мы должны представлять себе, что исходящее от двигательного анализатора движение, как и вообще, состоит из двух противоположных иннерваций: положительной и отрицательной, движения к предмету и от предмета, подобно отношениям сгибателей и разгибателей на конечностях. Тогда негативизм можно понимать следующим образом. Условный раздражитель из области коры, мало или совсем не заторможенный, посылает раздражение к соответствующему положительному иннервационному пункту двигательной области, а он в силу определенного состояния гипнотизации находится в парадоксальном состоянии. Поэтому это раздражение ведет не к возбуждению этого пункта, а приводит его в еще более глубокое торможение. Тогда это экстренное и локализованное торможение вызывает по правилу взаимной индукции возбуждение отрицательного пункта, как тесно ассоциированного с положительным пунктом. Отсюда первая отрицательная фаза негативизма. При удалении раздражителя, с одной стороны, тот же экстренно заторможенный положительный пункт сейчас же сам переходит на основании внутренней взаимной индукции в возбужденное состояние, а кроме того, индукционно возбужденный отрицательный пункт тоже сейчас же переходит в экстренно тормозное состояние и со своей стороны положительно индуцирует положительный пункт. Таким образом, положительный пункт после первого его экстренного торможения возбуждается как бы вдвойне. Соответственно этому обыкновенно, если гипнотизация не идет дальше, после одного или нескольких раз подачи и удаления еды положительная фаза берет перевес, собака начинает брать еду. Перед нами очень лабильное состояние клеточной деятельности, как качество переходной фазы. Что это так, доказывается дальнейшим ходом явлений. Далее, если гипнотическое состояние усиливается, остается только одна отрицательная фаза, обратное индуцирование делается невозможным, а дальше отсутствует и вообще какое-либо возбуждение двигательного иннервационного прибора.
Здесь же в этом приблизительно периоде условной пищевой двигательной реакции при гипнотизации обнаруживается одно из условий дробного распределения торможения по коре при гипнозе. На одной из наших собак, как показано в фактической части этой статьи, выступило очень интересное и своеобразное (уже упомянутое в ранней статье одного из нас) явление. Это – известная последовательность торможения в близких районах двигательной области. Эта последовательность может быть понята только так, что торможением раньше захватывается то, что перед надвигающейся гипнотизацией сильнее работало. Так как при повторяющемся акте еды больше всего работали жевательные мышцы и язык, затем шейные мышцы и потом туловищные, то и торможение обнаруживается в той же последовательности.
Интересный факт положительного раздражающего действия во время гипнотизации всякого малейшего изменения вида пищи и способа подачи еды имеет свое основание тоже в уже известной нам общей черте корковой деятельности. Давно в нашей лаборатории (д-ром Ю.В. Фольбортом) было установлено, что существует условное торможение второго порядка, как есть условное раздражение второго порядка. Явление состоит в следующем. Если с выработанным тормозным процессом (например, при дифференцировке) совпадает во времени несколько раз индифферентный раздражитель, то он скоро делается тормозным агентом. Тогда легко понять и то, что все, что надает на большие полушария во время гипнотизации (а она есть известная степень торможения), тоже делается тормозным. Вот почему и одного введения собаки в экспериментальную комнату иногда уже достаточно для ее загипнотизирования. А всякие новые раздражения, хотя бы и очень незначительные, естественно, этим тормозящим действием не обладают, а потому возбуждают положительную деятельность коры.
Приведенный в фактической части статьи терапевтический рефлекс есть только один из подкорковых рефлексов, которые выступают при гипнотизации после короткого подкармливания. Акт еды со всеми его раздражающими компонентами, как сильный раздражитель более или менее загипнотизированной коры, влечет за собой углубление торможения коры. Тогда с коры происходит положительная индукция на подкорковые центры, для которых в данный момент существуют субминимальные наличные раздражители или следы прошлых сильных раздражений. Животное начинает чихать, чесаться и т. д., чего при вполне бодром состоянии не делает. Сюда же относится и экспериментальный случай на собаке, аналогичный военному неврозу; случай этот и его анализ изложены в этом же томе «Трудов».
Что касается действия дифференцировок, т. е. условных тормозных раздражителей, то относительно их влияния на разлитое торможение мы давно знаем, что оно двойное, прямо противоположное. Если дело идет о слабой дымке разлитого торможения коры, о слабом напряжении гипнотического состояния, то хорошо выработанный тормозной раздражитель, концентрируя разлитое торможение в большей или меньшей степени, или совсем устраняет гипнотическое состояние, или переводит его в более легкие степени. Обратно, в случае сильного тормозного тонуса коры он углубляет торможение, как бы суммируясь с наличным торможением. Значит, результат определяется силовыми отношениями.
Наконец, последний опыт фактической части статьи: чрезвычайно сильный раздражитель, в противоположность умеренно сильным и слабым раздражителям, часто не углублял торможения, а производил положительное действие. Его надо отнести к прямому действию чрезвычайно сильного раздражителя на подкорку, а уже сильное возбуждение подкорки сообщалось и коре, рассеивая или ослабляя в ней тормозной процесс. Подтверждением правильности такого понимания дела может служить наш частый экспериментальный прием. Когда однообразная экспериментальная обстановка начинает гипнотизировать некоторых наших животных мы противодействуем этому, между прочим, тем, что повышаем пищевую возбудимость собак, уменьшая в известной мере их ежедневную порцию еды. А повышение пищевой возбудимости в этом случае всего естественнее полошить в подкорковом пищевом центре.

