12. Триплеты
Хотя мы с Сидни прекрасно понимали, что генетический код – проблема для биохимиков, мы все еще надеялись, что методы генетики смогут внести вклад в ее решение, тем более что генетические методы, при удачном выборе материала, могут принести плоды быстро, тогда как биохимические методы зачастую не столь оперативны. Сеймур Бензер, применив методы генетики, показал, что наследственный материал почти наверняка располагается линейно. Постановку вопроса вдохновило открытие двойной спирали ДНК, но сам метод был совершенно оригинален.
Для точного картографирования гена необходимо отыскать довольно редких особей. Чем ближе две мутации расположены в одном гене, тем меньше вероятность генетической рекомбинации между ними. Система, избранная Бензером, имела два преимущества. Гены, о которых идет речь, принадлежали бактериофагу Т4 – вирусу, который поражает клетки кишечной палочки. Вирус быстро растет и рекомбинируется с высокой скоростью. Он выбрал ген под названием rII – точнее, это пара соседних генов, – поскольку этот ген обладал важными техническими преимуществами. Подобрав нужные штаммы клетки-хозяина, можно было выделить вирус с геном дикого типа, даже если он смешан с миллионами вирусов мутантного варианта. Таким образом можно было отследить очень редкие рекомбинантные гены, настолько редкие, что, по расчетам Бензера, это позволяло даже разделить соседние пары оснований ДНК. К несчастью, не существовало обратного метода выделения мутанта среди множества вирусов дикого типа, однако при использовании подходящей бактерии-хозяина бляшка – малая колония, образованная зоной роста одной бактерии среди посева кишечных палочек в чашке Петри, – отличалась по внешнему виду, и ее было легко заметить. Единственную мутантную бляшку в чашке Петри среди сотен бляшек дикого типа отличить было сравнительно несложно.
Традиционный метод картографирования состоял в том, чтобы набрать ряд мутантов и затем определить рекомбинантное расстояние между любой парой из них. Были возможны и более точные методы с использованием трех мутантов, но все они требовали подсчетов сотен и даже тысяч бляшек, что было крайне трудоемко.
Бензер, никогда не любивший лишней работы, придумал метод получше. Он обнаружил, что, наряду с точечными мутациями, у некоторых из его мутантов, по-видимому, присутствуют делеции. На его генетической карте они выглядели как линии, поскольку перекрывали не менее двух точечных мутаций. Таким образом ему удалось собрать данные о целой серии делеций. Если две делеции перекрывались, то рекомбинация генов не могла вернуть организм к исходному дикому типу, поскольку перекрывающийся отрезок отсутствовал у обоих родителей и не мог быть восстановлен. Напротив, если две делеции не перекрывались, то при удачном случае рекомбинации дикий тип мог восстановиться.
Это можно пояснить с помощью аналогии. Представьте себе два поврежденных экземпляра книги: в одном не хватает страниц с 100 по 120, в другом – с 200 по 215. Очевидно, что по этим двум экземплярам, в каждом из которых имеется только одна непрерывная делеция, можно восстановить полный текст книги. Однако, если во второй книге утрачены не страницы 200–215, а страницы 110–125, восстановить страницы 110–120 не будет никакой возможности, поскольку они отсутствуют в обоих экземплярах.
Чтобы сделать аналогию точнее, немного расширим ее. Представьте себе, что книга содержит весьма подробные инструкции по изготовлению сложного инструмента. Предположим также, что если хотя бы одной страницы не хватает, то инструмент либо не получится, либо получится негодным к употреблению. Наконец, предположим, что у нас миллионы экземпляров каждой дефектной книги. Правило в таком случае следующее: берем по одному экземпляру каждого варианта книги. Затем берем первые n страниц из одной книги и оставшиеся страницы из другой. Проверяем, обеспечит ли новая гибридная книга изготовление работоспособного инструмента. Проделываем это миллион раз, отбирая пересекающуюся страницу (страницу n) всякий раз случайным образом. Если в какой-то момент инструмент получился пригодным, то две делеции не перекрываются. Если пригодный инструмент так и не получился, то делеции, вероятно, перекрываются.
Этот метод может показаться слишком мудреным, но другого у нас не было, ведь мы не могли заглянуть внутрь фага. Таким образом, Бензеру оставалось лишь скрестить два вируса, одновременно заразив ими культуру кишечной палочки. После их роста и рекомбинации внутри бактериальной клетки вирусы можно было размножить в чашке Петри на специальном штамме бактерии-хозяина. Если делеции не перекрывались, в посеве обнаружились бы рекомбинантные бляшки. Если они перекрывались, то ничего бы не изменилось. Заниматься трудоемкими подсчетами нужды не было. Все, что требовалось, – простой ответ «да / нет».
Бензер утверждал, что если ген имеет двумерную структуру, то рано или поздно обнаружится закономерность из четырех делеций. Делеция А будет перекрывать В и С, как и делеция D; но делеции B и С не будут перекрываться между собой, равно как и А и D (см. рис. на с. 211). Очевидно, что это невозможно, если структура гена одномерна, т. е. линейна. Бензер идентифицировал сотни делеций и все скрестил друг с другом попарно. Картины, отраженной на схеме, так и не получилось. Следовательно, заключил он, ген имеет линейную структуру. Его результаты также позволили ему расположить все делеции по порядку, так чтобы приблизительно определить их место на карте гена.
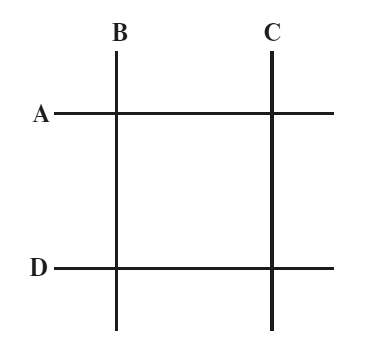
На плоскости это можно изобразить как пересечение линии А одновременно с В и С, в то время как линия D пересекается одновременно с В и С, не пересекаясь с А, а линия В не пересекается с С. Это было бы невозможно, если бы линии А, В, С и D были отрезками (одномерной) прямой. Поскольку Бензер так и не обнаружил подобного варианта перекрытий среди множества изученных им делеций, он сделал верный вывод, что ген, изучаемый им, имел линейную природу. Это укладывалось в гипотезу, что ген состоит из ДНК.
По ряду причин наша команда выбрала такой же метод исследований. Интересовал нас главным образом другой тип мутаций, вызываемый другими химическими мутагенами, а также обратные мутации, вызываемые этими веществами. Мутации, очевидно, делились в грубом приближении на два класса. Большинство химических мутагенов давало мутации первого класса. Однако мутации, возникающие под воздействием акридинов, попадали в другой. Мутации каждого класса было легче всего ревертировать с помощью того же мутагена, который их вызвал. Эрнст Фриз предположил, что первый класс составляют транзиции (замены пурина на пурин или пиримидина на пиримидин, см. Приложение А), тогда как второй – трансверсии, как их назвали (замена пурина на пиримидин и наоборот). Мы выдвинули другую идею. Одни мутации были ослабленными – то есть ген сохранял некоторую активность, пусть и не полноценную, – тогда как другие были неослабленными («непросачивающимися»), то есть ген, в сущности, отключался. Мы заметили, что мутации, вызванные профлавином (типичным акридином), почти всегда оказывались неослабленными. Это привело нас к предположению, что профлавиновые мутации – мельчайшие делеции или вставки в последовательности оснований, тогда как другой класс мутаций состоит из различных нуклеотидных замен. Однако у нас не было дальнейших доказательств этой идеи.
Тем временем я выступил с совершенно другой идеей. Размышляя над тем, как же молекула РНК может служить матрицей, я задумался, не закручивается ли она обратно сама на себя, образуя подобие двуспиральной структуры. Идея состояла в том, что одни основания могут образовывать пары, тогда как другие не могут соединиться по правилам образования пар и дадут выступающие петли. «Код» в таком случае будет зависеть либо от спаренных оснований, либо от петель, либо от какой-то более сложной комбинации двух возможностей. Это соображение было довольно туманным, но позволило сделать одно важное предсказание. Последствия мутации на одном конце матрицы теоретически могли быть скомпенсированы другим основанием, расположенным с другого конца, способным образовать пару с ним. Следовательно, у некоторых мутаций должны быть дистанционные супрессоры (как их теперь называют) в пределах того же гена.
Эта идея мне нравилась, но остальные были о ней невысокого мнения. До той поры я не имел опыта самостоятельной работы с генетикой фагов, довольствуясь наблюдениями за результатами опытов моих коллег. Поскольку никто не горел желанием проверить мою мысль, я взялся протестировать ее самолично. Выучиться генетическим исследованиям фагов было нетрудно, особенно при помощи специалистов. И все же я допустил несколько элементарных ошибок, к счастью, быстро исправленных. Благодаря опытам я также осознал, насколько поверхностны мои знания, пусть мне и случалось принимать участие в многочисленных дискуссиях о самом устройстве этой системы. Ничто не может заменить практических занятий экспериментами, если хочешь понять всю подноготную того, как это работает. Кроме того, это помогает закрепить в памяти детали, тем более что чтение раздела «Методы эксперимента» в научных публикациях, как правило, редкостная тягомотина.
Конечно же, для опытов я выбрал гены rII, сосредоточившись на втором из них, так называемом B-цистроне. (Цистрон – пижонское название, придуманное Бензером для гена, на основании так называемого цис-транс-теста.) Я выбирал из нашей культуры мутантный вариант, пытался отыскать реверантный – более близкий к дикому типу, – а затем проверял, вызывается ли реверсия второй мутацией, расположенной где-то в том же гене. Если мне не удавалось найти его, я продолжал опыты с другим мутантом.
Поначалу я не мог найти никаких супрессоров. Вероятно, замена, возвращающая мутантный ген обратно к дикому типу, располагалась там же, где первичная замена, или очень близко к ней, слишком близко, чтобы ее можно было отловить. Однажды ко мне зашел попить кофе Лесли Орджел. Он заглянул мне через плечо, и я объяснил ему, чем занимаюсь, посетовав, что результатов до сих пор нет. Он вернулся к остальным, а я поспешно проверил оставшиеся чашки Петри. К моей радости, оказалось, что кандидат на роль супрессора у меня есть.
Вскоре я располагал тремя мутантами с супрессорами, удачно расположенными вдоль карты. Я выделил супрессоры и стал их картографировать. Моя теория оказалась тут же опровергнута. Супрессоры не располагались в предсказанных точках, на некотором удалении от мутаций – напротив, каждый супрессор находился очень близко к мутации. Действие супрессоров должно было объясняться какими-то иными причинами.
Хотя я об этом и не знал, другие тоже отмечали, что мутации в rII может сопутствовать супрессор в том же гене. Вероятно, самый примечательный случай произошел в Калифорнийском технологическом институте. Дик Фейнман, физик-теоретик, настолько заинтересовался этими генетическими проблемами, что решил провести кое-какие эксперименты самостоятельно. Он наткнулся на образец внутреннего супрессора. Не зная, что это может значить, он посоветовался со своим руководителем, Максом Дельбрюком. Макс предположил, что исходный мутант продуцировал измененную аминокислоту и что вторая мутация изменила другую аминокислоту где-то в другом месте молекулы белка, и это каким-то образом компенсировало первое изменение. Такую возможность допустить было легко, но что такое явление может быть распространенным, ожидать не приходилось.
Я, безусловно, знал о такой возможности, но она меня не устраивала, отчасти потому, что я тщательно изучил все то немногое, что на тот момент было известно о структуре белков. Я решил разобраться, сколько различных супрессоров может быть у одной данной мутации. Для дальнейшего исследования мне требовалось выбрать одного мутанта из трех, и из практических соображений я выбрал того, у которого супрессор располагался дальше всех от исходной мутации, надеясь, что это даст мне больше степеней свободы. Я заметил также, что две из трех мутаций были вызваны профлавином. Хотя это было едва ли значимо статистически, мне это показалось любопытным.
К тому времени я приобрел некоторый опыт, так что эксперименты пошли довольно быстро. Преимущество генетики вирусов – в скорости экспериментов, при условии, что все отлажено. Можно за короткий срок произвести сотню скрещиваний, поскольку процедуры несложные, а само скрещивание занимает каких-то минут двадцать – на то, чтобы фаг поразил бактерию, размножился внутри ее клетки (обмениваясь генетическим материалом в ходе этого процесса) и прорвал ее, отчего клетка погибает. Результаты скрещиваний следует затем высеять в чашку Петри с бактериальной пленкой. Потом чашки подвергаются инкубации, чтобы выросла культура бактерий. Там, куда попал отдельный фаг и заразил клетку, вырастет колония фагов, по мере роста убивая бактерии вокруг себя и образуя отчетливую дырочку (бляшку) в культуре бактерий, растущих на поверхности питательной среды. Этот процесс занимает несколько часов, так что есть время немного отдохнуть. Затем чашки Петри вынимают из инкубатора, где их выдерживали при 37 ºС, и обследуют на наличие бляшек – есть ли они и какого типа. Наиболее любопытные бляшки затем «выделяют» – то есть берут из них немного фагов с помощью бумажки или зубочистки, снова размножают и повторяют процесс заново, чтобы убедиться, что популяция штамма чистая. Если постараться, можно завершить серию скрещиваний за один день и подготовиться к новой серии на завтра.
Эксперименты становились всё интереснее и интереснее, и я убедился, что при тщательном планировании можно провести две последовательных серии скрещиваний за день. Для этого нужно было вставать рано поутру, возвращаться домой на обед, снова экспериментировать в послеобеденное время, опять домой – ужинать, и опять в лабораторию заканчивать опыт после ужина. К счастью, мы с Одилией жили в нескольких минутах ходьбы от лаборатории, в историческом центре Кембриджа, и работа была мне не в тягость. По правде говоря, если верить Одилии, я никогда не бывал веселее, чем в тот период, когда экспериментировал с утра до ночи, – но этим я обязан, вероятно, отчасти тому, что вот уже несколько недель опыты продвигались великолепно.
Вскоре я обнаружил, что у моей первоначальной мутации имеется не один, а несколько различных супрессоров, и все расположены достаточно близко к самой мутации. Я решил дать им специальное название. Зачастую я продолжал работу на выходных, а в понедельник брал отгул, чтобы наша лабораторная кухня (где мыли всю посуду, в том числе стерилизовали чашки Петри перед работой) успела все вымыть. Вышло так, что новое название понадобилось во время выходных, когда рядом никого не было. Мутации обычно обозначались буквами, за которыми шли цифры. Например, P31 означает тридцать первую мутацию в серии P, вероятно, вызванную профлавином. На беду, я не смог точно вспомнить, до какой буквы мы уже дошли, поэтому решил назвать своего мутанта FC0 – будучи уверен, что мои инициалы для обозначения мутаций никто покуда не использовал. Новые супрессоры получили затем наименования FC1, FC2 и так далее. Из-за того, что я назвал их своими инициалами, меня попрекали тщеславием, но истинное объяснение – в том, что у меня дырявая память.
Все новые супрессоры с виду были надежными, неослабленными мутациями. А если, подумалось мне, у них тоже есть супрессоры? Так и оказалось. Я даже пошел дальше и обнаружил супрессоров супрессоров супрессоров.
Как же это работало? К счастью, у нас под рукой уже были подходящие гипотезы. Допустим, генетическое послание читается (при синтезе белка) по три нуклеотида за один прием, начиная с определенного места. Для ясности возьмем простейший «текст», в котором просто повторяется триплет ТАГ – раз за разом:
… ТАГ, ТАГ, ТАГ, ТАГ, ТАГ, ТАГ …
Многоточия означают, что кодирующий «текст» стоит и перед последовательностью, и после нее. Запятые для наглядности отмечают рамку считывания. Я предположил, что эта рамка задается специальным «стартовым» сигналом, расположенным где-то левее указанного отрезка.
Предположим, что наша исходная мутация (получившая название FC0) прибавляет одно основание (нуклеотид) к нуклеотидной последовательности. Следовательно, с этого места рамка считывания сдвинется на один шаг, и получится бессмысленный белок – белок, чья аминокислотная последовательность, порожденная мутацией, окажется совершенно неправильной, и продукт гена не сможет функционировать. Тогда наша простейшая последовательность превращается в:
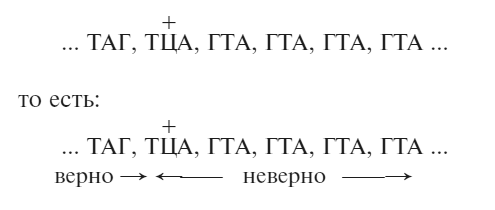
(Лишнее основание для наглядности обозначено как Ц, но оно может быть каким угодно из четырех.)
В таком случае супрессор, подобный FC1, будет делецией одного основания где-то поблизости. В промежутке между FC0 и FC1 последовательность останется бессмысленной, она по-прежнему будет считываться со сдвигом, но далее считывание будет происходить нормально.
Наш пример приобретет следующий вид:
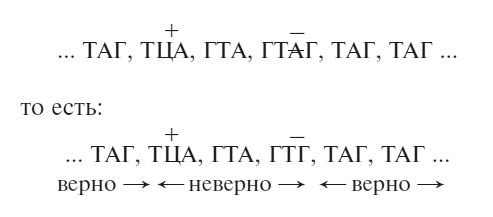
Если измененный отрезок аминокислотной последовательности не очень важен (а в данном случае были и другие свидетельства в пользу этого), то белок будет работать достаточно хорошо и носитель двух мутаций (FC0 + FC1) будет ближе к дикому типу, чем носитель неослабленной мутации.
Потому я обозначил первый ряд супрессоров знаком минус. Следующий ряд – супрессоров первого ряда супрессоров – мы обозначили плюсом, а их супрессоров – снова минусом.
К опытам я приступил в начале мая, теперь же лето было в разгаре. Я уже спланировал заранее поездку с семьей в летний отпуск – едва ли не первый настоящий отпуск в нашей совместной жизни, поскольку теперь мое финансовое положение немного улучшилось. За совсем небольшую плату мы сняли большую виллу на древней горе в Танжере – североафриканском городе у самого Гибралтара. Там мы вели роскошную жизнь: у нас был один постоянный слуга-араб и еще один приходящий ежедневно. Одилия вместе с нашей домработницей-немкой, Элеанорой, учились покупать продукты на арабском базаре, торгуясь, притворяясь, что уйдут, и тому подобное. Наши две дочки упражнялись в плавании на море, я же обычно проводил дни на террасе, в пятнистой тени пальм.
По пути в Танжер я заехал на одну конференцию. Даже в те времена ученые с неохотой посещали конференции, если только они не проводились в каком-то интересном месте. Эта проводилась на перевале Коль де Во, на полдороге вверх по склону Монблана. Я сделал доклад о своих предварительных результатах, которые впоследствии были опубликованы в виде чрезвычайно краткого релиза в материалах конференции.
Проведя месяц в Танжере, я отправился на Биохимический конгресс 1961 г. в Москве. Моя семья осталась на вилле еще на неделю или около того. Москва заметно изменилась со времени моего первого приезда в 1945 г., во время войны. Теперь было лето, а не середина зимы, город выглядел более ярким и цветущим, чем в унылое военное лихолетье. Я поселился в общежитии университета, где проводился конгресс, и познакомился с некоторыми из русских организаторов. Среди них выделялся Игорь Тамм, русский физик. Влияние Лысенко – человека, который на какое-то время уничтожил генетику в СССР, – ощутимо клонилось к закату. По моим впечатлениям, закат этот был во многом заслугой физиков наподобие Тамма, обладавших значительным политическим влиянием и способных распознать бред в науке с первого взгляда. Многие из нас получили приглашения сделать доклады в рамках биологической секции Российского института атомной энергии – событие, невозможное еще несколько лет назад. Доклады мы читали на английском, но их великолепно переводил (блоками, в ходе выступления) Бреслер, русский ученый, с которым мы уже были знакомы – он приезжал в Кембридж. Бреслер не только понимал, о чем мы говорим, но в иных случаях, насколько я мог судить на слух, пояснял ссылки, которые давали докладчики, – воистину замечательная работа.
Московский конгресс стал особенно интересным благодаря результатам, изложенным в докладе Маршалла Ниренберга, тогда мало кому известного. О его опытах я знал понаслышке, но без подробностей. Мэтт Мезельсон, с которым я столкнулся в коридоре, затащил меня на доклад Маршалла в семинарской аудитории где-то в закоулках. Он произвел на меня такое впечатление, что я пригласил Маршалла принять участие в более масштабной секции, где я председательствовал. Он открыл, что можно добавить искусственную матрицу в пробирку, где идет синтез белков, и заставить ее управлять синтезом. А именно, он добавил ряд У – матричную РНК, состоящую только из цепочки урацилов, – и система синтезировала полифенилаланин. Это означало, что триплет УУУ (если код действительно состоит из триплетов) служит кодоном для фенилаланина, и так и оказалось. Позднее я утверждал, что публика была ошеломлена (по-моему, я употребил слово «наэлектризована») этими вестями. Однако Сеймур Бензер опроверг это с помощью фотографии, на которой все, судя по выражению лиц, помирают со скуки! И все же это было эпохальное открытие, после которого прогресс было уже не остановить.
Кроме того, во время своего недельного пребывания в Москве я ходил в гости. Я любил приходить в одну старомодную квартиру с тяжелой мебелью и кроватью за огромным книжным шкафом. И в другую, более современную, оформленную в куда более светлых тонах. Ее хозяин коллекционировал современное русское искусство. Меня развеселило, когда я увидел, что Алекс Рич показывает хозяину странный новый американский танец (потом я распознал в нем твист). Поскольку талия у Алекса не особенно выделяется, твист в его исполнении смотрелся не то чтобы бойко.
Я вернулся в Кембридж. Следовало продолжать эксперименты дальше, чтобы обосновать наше обозначение мутаций гена rII плюсом или минусом, подтвердив, что в нем есть какой-то смысл. Теория предсказывала, что всякая комбинация вида (+ +) или (– ) будет мутантной. Мы с коллегами сконструировали множество подобных пар, и все они, как и предсказывалось, оказались неослабленными мутантами. В упрощенном виде теория также предсказывала, что всякая комбинация вида (+ —) будет диким типом или чем-то близким к нему. Конечно, мы знали, что для некоторых случаев это верно, поскольку именно так мы изначально сумели выделить супрессор, но многие другие комбинации плюса с минусом оставались непротестированными. Их мы назвали «дяди и тети», поскольку для их создания нередко требовалось совместить мутацию одного поколения с мутацией из предыдущего поколения, но не прямого предка первой. Я попросил Сидни проследить за некоторыми из этих опытов в мое отсутствие, но у него были собственные планы, так что по возвращении мне пришлось проделать это самому.
Тут возникло небольшое осложнение. Некоторые из комбинаций (+ —), которые должны были быть дикого типа, оказались мутантными. Мы избавились от этого затруднения, предположив, что в некоторых случаях мелкий локальный сдвиг рамки считывания между плюсом и минусом давал «бессмысленную» мутацию. Теперь известно, что бессмысленные отрезки возникают из-за триплета, обрывающего синтез полипептидной цепи, в результате чего получается нефункциональный фрагмент белка. Я также понял, что это зависит от точного расположения рамки считывания. Для кода из неперекрывающихся триплетов существует одна верная рамка считывания, но две неверных, так что на локальном уровне комбинация (+ —), то есть плюса, за которым идет минус, будет отличаться от комбинации (– +).
Если вернуться к нашему простому примеру, комбинация (+ —) может выглядеть так:
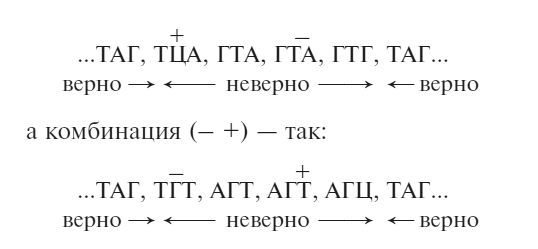
В первой между двумя изменениями стоит последовательность ГТА, во второй – АГТ. Мы продемонстрировали, что наши неудачные варианты (+ —) или (– +) подчиняются тому же правилу, и это вселило в нас уверенность, что мы на правильном пути.
Перед тем Сидни пришла в голову еще одна мысль. Он допустил, что мутантный вариант (+ +) может реверсировать назад к дикому типу. Он попытался провести такой опыт, но обратная мутация, вероятно, располагалась слишком близко к исходной, поскольку он не смог идентифицировать ее. Другой, несколько более трудоемкий подход состоял в том, чтобы сконструировать тройного мутанта вида (+ + +) или (– —). Наша теория предсказывала, что получится дикий тип, так как три последовательных сдвига по фазе должны были восстановить нужную рамку считывания – разумеется, при условии, что код состоит из триплетов.
Для нашей простейшей последовательности пример мог бы выглядеть так:
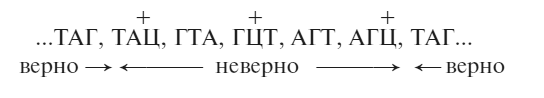
Прямой, но трудоемкий способ создать такого тройного мутанта – выбрать три мутации, все со знаком + и не слишком удаленные друг от друга, затем собрать две пары с одинаковой средней по счету мутацией (см. рис. ниже). Трудоемок он потому, что не существует метода отобрать подобную комбинацию мутаций. Необходимо проводить скрещивание и упорно проверять потомство с мутантным фенотипом, перебирая по отдельности, пока не найдется искомое сочетание (+ +). Последний этап не представляет затруднений. Нужно просто скрестить два двойных. Поскольку каждый содержит среднюю мутацию из трех, истинный дикий тип получиться никак не может. Если же скрещивание дает бляшки, явно похожие на дикий тип, то они почти наверняка и относятся к искомой комбинации (+ + +). В любом случае, это легко проверить, разобрав по частям предполагаемый триплет.
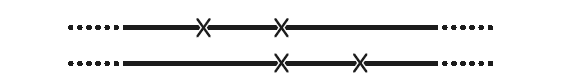
Каждая линия изображает один из двух родительских штаммов. Крестики обозначают мутации. Невозможно рекомбинировать два родительских штамма так, чтобы получить штамм вовсе без мутаций. Средняя мутация будет сохраняться всегда. Более того, у некоторых потомков все три делеции могут оказаться в одном штамме.
Разумеется, триплет будет походить на дикий тип лишь при условии, что код действительно состоит из триплетов. Если основания считываются по 4 или 5 за раз, – мы не могли исключить такую возможность, – вариант (+ + +) будет мутантным, и нам придется сконструировать сочетания (+ + + +) и даже (+ + + + +). Не все сотрудники лаборатории были уверены, что эксперимент удастся. Я же почти не сомневался в успехе. Как и Сидни, который на тот момент уехал в Париж. Он составил список из трех возможных вариантов комбинации (+ + +) для испытания, но, на счастье, после его отъезда я догадался, что два из них, скорее всего, не годятся, поскольку они дадут стоп-кодон, так что мы сконструировали третий, который, по-видимому, не создавал подобных осложнений.
К тому времени я привлек себе на помощь Лесли Барнетта. Были проведены последние этапы скрещиваний, батарея чашек Петри выставлена в инкубатор. После обеда мы зашли их проверить. Нам хватило одного взгляда на ту судьбоносную чашку. Бляшки были! Тройной мутант демонстрировал признаки (фенотип) дикого типа. Мы тщательно проверили номера на чашках Петри, чтобы удостовериться, что нам попалась нужная. Все было в порядке. Я взглянул на Лесли. «Ты хоть понимаешь, – спросил я, – что мы с тобой единственные в мире, кто знает, что код состоит из триплетов?»
Ведь результат и вправду был потрясающий. Перед нами было три различных мутации, каждая из которых отключала определенную функцию гена. Из них мы могли сконструировать три возможных двойных мутации. Каждая из них по отдельности также делала ген нерабочим. Но если мы совмещали все три в пределах одного и того же гена (а мы с помощью отдельных экспериментов продемонстрировали, что все они находятся в одной вирусной частице, не так, что часть попала в один вирус, а остальные – в другой), то ген снова начинал работать. Это было легко объяснимо, если мутации в самом деле представляли из себя вставки или делеции и если код вправду состоял из триплетов. Короче говоря, мы представили первые убедительные доказательства в пользу того, что код триплетный.
Ну, я слегка преувеличиваю. Данные укладывались также в картину кода с шестью основаниями на кодон, но этот вариант, как показали дополнительные эксперименты, был маловероятен, и его не приходилось принимать всерьез.
У нас все еще оставалось много работы по уточнению результатов. Мы сконструировали не одного, а шесть различных тройных мутантов – пять типа (+ + +) и одного (– —) – и продемонстрировали, что все они ведут себя как дикий тип. Хлопот у меня стало еще больше, хотя теперь мне оказывал немалую помощь Лесли. Не то чтобы отдыха совсем не было. Однажды вечером, после ужина, я работал в лаборатории, когда вдруг объявилась моя фешенебельная знакомая. Она стояла у меня за спиной, пока я продолжал возиться с пробирками и чашками Петри. «Приходи на вечеринку», – сказала она, запустив пальцы мне в волосы. «Я занят до невозможности, – ответил я, – а где она?» «Вообще-то, – сказала она, – мы думали устроить ее у тебя дома». В конце концов компромисс был достигнут. Они с Одилией организуют небольшой прием гостей, а я приду к ним, когда закончу работу.
Оглядываясь назад, я удивляюсь, как мало мы работали – ведь я летом уезжал на шесть недель, бывал на Монблане, в Танжере и Москве, – и как при этом быстро и напряженно мы трудились. К решающему эксперименту я приступил в начале мая, а статья вышла в последнем выпуске Nature за тот же год.
Мы не остановились на достигнутом. В особенности Сидни, который в дальнейшем провел много остроумных экспериментов с нашей системой. В итоге мы решили, что лучше опубликовать полноценный отчет, так что мы с Лесли Барнеттом приложили усилия, чтобы увязать все концы. Это привело к одному важному результату. К тому времени было уже известно, что триплеты УАА и УАГ являются стоп-кодонами. Я был убежден, что УГА – третий стоп-кодон. Сидни изобрел хитроумный способ проверить это генетически, но наши опыты всегда давали отрицательный результат. Подводя письменные итоги, мы обнаружили, что провели не все возможные эксперименты данного типа. Мы решили не оставлять пробел в наших таблицах, а попросить Лесли в обычном порядке провести те опыты, которые были упущены. К нашему удивлению, теперь результат был положительный! Тогда мы повторили все прежние опыты, и на этот раз они тоже удались! Выяснилось, что во время их проведения мы использовали набор контролей, чтобы убедиться, что все протекает как надо. К несчастью, в каждом опыте тот или иной контроль оказывался упущенным. Когда все контроли были задействованы должным образом, полученный результат дал весомые основания предположить, что УГА – тоже стоп-кодон.
Мы планировали с почетом похоронить наши результаты в августовском сборнике «Философских трудов Королевского общества». Поскольку теперь мы получили кое-что более интересное, мы изъяли описания экспериментов из статьи, предназначенной для «Философских трудов», и сделали из них отдельную статью, которая вскоре вышла в Nature. Я несколько удивился, увидев свое имя в подготовленной рукописи, поскольку в обычае нашей лаборатории было подписывать своим именем лишь те статьи, куда ты внес существенный вклад. Просто дружеский совет на соавторство не тянул. «Почему ты вписал меня?» – спросил я у Сидни. Он ухмыльнулся. «Из вредности», – ответил он, и я не стал спорить.
Один из наиболее трудоемких экспериментов Лесли состоял в том, чтобы собрать шесть «плюсов» в одном гене и доказать, что получившийся мутант похож на дикий тип. Трудно описать, насколько подобный эксперимент утомителен и сложен. Искомые шесть «плюсов» требуется соединять поэтапно, на каждом этапе проверяя, получилась ли нужная структура гена. Когда получена и испытана конечная комбинация, ее нужно еще разобрать на части, тоже поэтапно, чтобы убедиться, что она соответствует теории. Даже обзорное описание того, что проделал Лесли, заняло несколько широкоформатных страниц «Философских трудов».
Когда мы занимались окончательной вычиткой рукописи, я сказал Сидни, что, по моему мнению, мы с ним окажемся единственными в мире, кто читал эту статью внимательно. Смеха ради мы решили вставить фальшивую ссылку. Прибавив в одном месте: «Леонардо да Винчи (в частной беседе)», – мы подали статью в Королевское общество. Один из рецензентов (оставшийся нам неизвестным) пропустил ссылку без обсуждения, но нам позвонил Билл Хейз, второй рецензент. Он спросил: «А что это за молодой итальянец работает у вас в лаборатории?» С неохотой мы удалили ссылку.
Генетическое доказательство того, что код состоит из триплетов, было эффектным достижением, но очень скоро этот факт был установлен прямыми биохимическими методами. Еще важнее в долгосрочной перспективе стало доказательство того, что акридиновые мутации представляли собой мелкие делеции и выпадения участков ДНК. Даже об этом уже в какой-то мере догадывались, потому что Леонард Лерман получил весьма многообещающие физико-химические данные в пользу того, что молекулы акридинов втискиваются между основаниями ДНК, а это запросто может привести к вставкам или делециям участков ДНК при копировании. Вместе с тем теория нуждалась в подтверждении прямыми биохимическими методами. Билл Дрейер и Георг Штрайзингер, оба биохимики, планировали добиться этого, хотя ответ они получили не скоро – в то время заниматься биохимией было нелегко с технической точки зрения. Ежемесячно мы с Сидни обсуждали, не стоит ли нам самим взяться за это, но приступить не решались, тем более что Георг был «своим» – то есть успел поработать в нашей лаборатории. В конце концов Георг получил нужные данные – не на материале неизвестных продуктов двух генов, а на вирусном лизоциме. Вышло именно так, как мы и ожидали. В промежутке между мутациями аминокислотная цепочка явно изменилась, и более того, результаты укладывались в картину знаний о генетическом коде, которая начинала вырисовываться.
Вскоре после этого я посетил симпозиум, организованный биологом Конрадом Уоддингтоном (для друзей – Уод) на вилле Сербельони на озере Комо. Там я впервые познакомился с математиком Рене Торном. Чуть ли не первые слова, которые я от него услышал, – что наши исследования акридиновых мутаций наверняка ошибочны. Поскольку до меня уже дошли вести, что наши идеи были доказаны биохимически, я несколько удивился и спросил его, почему он так считает. Он объяснил, что, создавая, например, тройную мутацию, мы неизбежно получим пуассоновское распределение одинарных, двойных, четверных и т. д., так что наши доводы неосновательны. Поскольку мы провели кропотливую сборку наших многочисленных мутантов (и каждый подвергался тщательной проверке), мне с первого взгляда стало ясно, что его возражение не имеет силы, будучи основанным на недоразумении. То ли он недостаточно внимательно прочел нашу статью, то ли прочел, но не понял. Впрочем, по моему опыту, математикам большей частью свойственна леность ума, и в особенности они не любят читать экспериментальные работы.
Мне Рене Торн показался хорошим математиком, но несколько чванным – его раздражала необходимость растолковывать свои идеи языком, понятным для не-математиков. К счастью, на симпозиуме присутствовал другой тополог, Кристофер Зиман, отлично умевший пересказать идеи Торна.
Еще мне показалось, что Торн по существу слабо понимает, как устроен процесс научного исследования. Того, чего он не понимал, он не одобрял и презрительно называл «англосаксонским». По моим впечатлениям, он был наделен недюжинной биологической интуицией, но, к несчастью, со знаком «минус». Подозреваю, любая высказанная им идея в области биологии наверняка оказалась бы ошибочной.

