Книга: Энергия и цивилизация
Назад: 2. Энергия в доисторические времена
Дальше: Истоки сельского хозяйства
Общества охотников-собирателей
Самые достоверные собрания надежных доказательств показывают, что средняя плотность населения современных обществ охотников-собирателей – отражающая различие среды обитания, навыки и техники добывания пищи – варьируется в пределах трех порядков (Murdock 1967; Kelly 1983; Lee and Daily 1999; Marlowe 2005). Минимум составляет меньше 1 человека на 100 км2, максимум – несколько сотен человек на 100 км2, со средним значением в 25 человек на 100 км2 для 340 изученных культур. Слишком низкая плотность, чтобы поддерживать более сложные общества с возросшей функциональной специализацией и социальной стратификацией. Средние значения плотности охотников-собирателей были ниже, чем средняя плотность травоядных со сравнимой массой, способных переваривать изобильную целлюлозную фитомассу. Аллометрические уравнения предсказывают значение около 5 млекопитающих весом около 50 кг на квадратный километр, при этом плотность шимпанзе составляет 1,3–2,4 животных на километр, а плотность охотников-собирателей XX века была ниже одного человека на километр в теплом климате и 0,4 в Новом Свете (Marlowe 2005; Smil 2013а). Плотность популяции была значительно выше для групп, комбинировавших собирание имевшихся в изобилии растений с охотой (хорошо изученные образцы включают группы в постледниковой Европе и, в относительно недавнее время, на Мексиканском нагорье) и для прибрежных общин, зависящих в основном от морских видов (с хорошо документированными археологическими находками в балтийском регионе и недавними антропологическими исследованиями на северо-западе Тихого океана).
Собирание моллюсков, рыбная ловля, прибрежная охота на морских млекопитающих поддерживали наиболее высокую плотность охотников-собирателей, и подобная практика вела к появлению полупостоянных и даже постоянных поселений. Прибрежные деревни северо-запада Тихого океана с их большими домами и организованной совместной охотой на морских млекопитающих выглядели исключительными по уровню оседлости. Подобные значительные вариации плотности были не просто функцией от потоков биосферной энергии: значения не уменьшались по направлению к полюсам, и не увеличивались по направлению к экватору (в пропорции к более высокой фотосинтетической продуктивности), и не соотносились с общей массой животных как объектов охоты. Вариации определялись переменными экосистемы, сравнительной зависимостью от растительной и животной пищи, использованием сезонных запасов. Как и большинство приматов помимо человека, все охотники-собиратели были всеядными, но убийство более крупных животных оказалось главным энергетическим вызовом, поскольку его целью становился куда меньший по объему резервуар съедобных веществ, чем содержалось в растениях для собирательства: естественное последствие уменьшения энергии, переносимой между трофическими уровнями.
Травоядные потребляют только 1–2% от полезной первичной продуктивности в листопадных умеренных лесах и до 50–60 % на некоторых тропических лугах, за среднюю величину можно взять 5-10 % (Smil 2013а). В целом меньше чем 30 % от потребленной фитомассы переваривается; большая ее часть тратится зря, только 1–2% от нее в случае птиц и млекопитающих становится зоомассой. В результате травоядные, на которых чаще всего охотятся, заключают в себе менее процента энергии, изначально запасенной в фитомассе экосистемы, где они обитают. Это объясняет, почему охотники предпочитают убивать животных, которые сочетают сравнительно большую массу тела с высокой продуктивностью и плотностью популяции: дикая свинья (90 кг), олень и антилопа (25-500 кг) были общей целью охотников.
Там, где подобные животные были сравнительно широко распространены, например в тропических или умеренных лесах и на лугах, охота представлялась более благодарным делом, но в противоположность общему мнению по поводу изобилия видов животных, тропические леса находятся в самом низу списка экосистем, благоприятных для охотников. Большая часть животных, обитающих в такой среде – древесные, питающиеся листвой и фруктами (обезьяны, птицы), они активны и слабо достижимы в кронах деревьев (а многие еще и ночные), и охота на них обещает низкую энергоотдачу. Обнаружено (Sillitoe 2002), что и собирательство, и охота в тропических дождевых лесах нагорий Папуа Новой Гвинеи очень затратны и живущие там люди расходуют в четыре раза больше энергии на охоту, чем получают с пищей. Очевидно, что при такой низкой энергоотдаче охота не может стать первоочередным способом добывания пищи (отрицательную энергоотдачу можно объяснить только необходимостью в животном белке), и поэтому для обеспечения достаточного количества пищи требовались те или иные формы кочевого земледелия.
Ученые считали (Bailey and co-workers 1989), что однозначных этнографических сообщений об охотниках-собирателях, живущих в тропических лесах без какой-либо опоры на одомашненных животных и на растения, не существовало. Позже это мнение изменилось (Bailey and Headland 1991), когда поступили археологические доказательства из Малайзии, что изобилие диких свиней и саговых пальм допускает некоторые исключения. Собирательство часто было удивительно неблагодарным как в изобильных видами тропиках, так и в умеренных лесах. Эти экосистемы содержат большую часть фитомассы планеты, но она заключена чаще всего в тканях высоких древесных стволов, целлюлозу и лигнин из которых человек не в состоянии переварить (Smil 2013а). Богатые энергией фрукты и семена составляют малую долю от общей массы растений и часто недоступны в высоких кронах; семена обычно защищены твердой скорлупой и требуют энергозатратного процесса очистки перед употреблением. Собирательство в тропических лесах также требует долгого поиска: большое разнообразие видов означает, что деревья или лозы одного вида расположены друг от друга на большом расстоянии (рис. 2.2). Сбор бразильского ореха может быть прекрасным примером подобной ситуации (примечание 2.2).
По контрасту с часто неудачной охотой в тропических и умеренных лесах травянистые равнины или лесостепь предлагают прекрасные возможности для собирательства и охоты. В этих экосистемах содержится куда меньше энергии на единицу площади, чем в густом лесу, но большая ее доля присутствует в форме легко собираемых и высокопитательных семян и фруктов или сконцентрирована в скоплениях крахмалистых листьев и клубней. Высокая плотность энергии (до 25 МДж/кг) делает орехи в особенности полезными, и некоторые из них, такие как желуди или лесной орех, очень легко собирать. И, в отличие от лесных, многие животные, пасущиеся на травянистых равнинах, вырастают до значительного размера, часто передвигаются большими стадами и поэтому обещают значительную отдачу на энергию, инвестированную в охоту.

Рисунок 2.2. Тропический дождевой лес может похвастаться изобилием населяющих его видов, но сравнительно беден растениями, которые могут поддерживать большие популяции охотников-собирателей. На фотографии – кроны джунглей в Ла-Фортуна, Коста-Рика (Corbis)
Примечание 2.2. Сбор бразильского орехаБразильский орех благодаря высокому содержанию липидов (66 %) характеризуется энергией в 27 МДж/кг (сравните с примерно 15 МДж/кг для злаковых культур), в нем около 14 % белка, он также является источником натрия, магния, кальция, фосфора и даже селена (Nutrition Value 2015). Сбор этих орехов одновременно сложен и опасен. Bertolletia excels вырастает до 50 метров, и отдельные деревья стоят далеко друг от друга. От 8 до 24 орехов собраны в тяжелые (до 2 кг) капсулы, покрытые эндокарпом подобно кокосу. Собиратели орехов должны правильно выбирать время сбора: слишком рано, и «стручки» останутся недоступными на вершинах деревьев, так что придется идти еще раз; слишком поздно, и агути (Dasyprocta punctata), крупные грызуны и единственные животные, способные добраться до мякоти орехов, съедят их прямо на месте или спрячут в собственных кладовых (Haugaasen et al. 2010).
Гоминины могли добывать мясо в степях и лесостепях даже безо всякого оружия, действуя как падальщики, или как несравненные загонщики, или как коварные строители ловушек. Учитывая, что физические возможности первых людей не впечатляли, а эффективное оружие отсутствовало, наиболее вероятно, что наши предки и в самом деле были скорее падальщиками, чем охотниками (Blumenschine and Cavallo 1992; Pobiner 2015). Крупные хищники – львы, леопарды, саблезубые кошки – нередко оставляли после себя частично объеденные тела травоядных. Это мясо, или по меньшей мере питательный костный мозг, могло доставаться нашим предкам, а не их конкурентам в виде грифов, гиен и других падальщиков.
Но есть точка зрения (Dominguez-Rodrigo 2002), согласно которой сбор падали не мог дать достаточное количество мяса, и что только охота на травянистых пространствах могла обеспечить нужный объем белка. В любом случае, прямохождение человека и его «умение» потеть сильнее любого другого млекопитающего давало ранним людям возможность преследовать и загонять до полного изнеможения даже самых быстрых травоядных (примечание 2.3).
Некоторые исследователи (Carrier 1984) считают, что выдающаяся скорость рассеивания тепла человеком обеспечила ему важное эволюционное преимущество, которое отлично послужило нашим предкам, когда они заняли новую нишу – дневного, высокотемпературного хищника. Свойство обильно потеть, а значит, активно работать при высоких температурах осталось и у человеческих популяций, которые мигрировали в зоны холодного климата: не обнаружено значительной разницы в плотности мерокринных желез у населения различных температурных зон (Taylor 2006). Обитатели средних и высоких широт могут потеть точно так же, как аборигены тропических областей, после краткого периода акклиматизации.
Примечание 2.3. Бег и рассеивание тепла у людейВсе четвероногие имеют оптимальные скорости для разных видов передвижения, например шаг, рысь и галоп у лошадей. Энергетические затраты бега человека относительно высоки по сравнению с затратами на бег млекопитающих сравнимой массы, но в отличие от них, у человека объем затрат прямо не связан со скоростью бега, если она между 2 и 6 м/с (Carrier 1984; Bramble and Lieberman 2004). Прямохождение и эффективное рассеивание тепла объясняют этот подвиг. Вентиляция четвероногих ограничена одним вдохом на двигательный цикл. Грудные кости и мускулы должны поглотить ударную нагрузку от передних конечностей, в то время как спинно-брюшной каркас ритмично сжимает и расширяет грудную клетку, а вот частота человеческого дыхания может варьироваться произвольно. Люди могут бежать с какой угодно скоростью, а у четвероногих набор оптимальных скоростей ограничен структурно.Экстраординарные способности человека в области терморегуляции опираются на очень высокую скорость потоотделения. Лошади теряют воду с часовой интенсивностью в 100 г/м2 шкуры, верблюды – 250 г/м2, но человек может терять больше 500 г/м2, а пиковые значения достигают 2 кг/м2 (Torii 1995; Taylor and Machado-Moreira 2013). Скорость потения определяет потерю тепла в 550–625 Вт, что достаточно для регуляции температуры даже во время исключительно тяжелого труда. Люди также могут пить меньше, чем они теряют влаги, и восполнять частичное обезвоживание многими часами позже. Именно бег превратил людей в дневных высокотемпературных хищников, которые могли загонять животных до истощения (Heinrich 2001; Lienberg 2006). Документированные случаи загонной охоты включают индейцев тарахумара в Северной Мексике, способных загнать оленя; пайутов и навахо, жертвами которых были вилорогие антилопы. Басарва из Калахари могли добывать дукеров, ориксов, а во время сухого сезона даже зебр, ну а некоторые аборигены Автралии проделывали то же самое с кенгуру. Бегущие босиком люди снижают потерю энергии примерно на 4 % (и реже страдают от повреждений лодыжек и хронических болезней ног) по сравнению с бегунами в легкоатлетических кроссовках (Warburton 2001).
Но как только были изобретены и вошли в обиход необходимые инструменты, охота с их помощью получила преимущество перед загоном добычи, и после исследования 51 коллекции находок среднего каменного века и 98 из позднего каменного века (Faith 2007) подтвердилось, что первые африканские охотники умело добывали крупных травоядных, в том числе быков. Энергетические императивы охоты на крупных животных также внесли значительный вклад в социализацию человека. Исследователи (Trinkaus 1987, 131–132) сделали вывод, что «большая часть отличительных характеристик человека, таких как прямохождение, ловкость рук, продвинутые технологии и заметное увеличение головного мозга, могут быть рассмотрены как требования, выдвинутые гибкой и развитой системой добывания пищи».
Роль охоты в эволюции человеческих сообществ выглядит самоочевидной. Индивидуальный успех в охоте на крупных животных с помощью примитивного оружия крайне маловероятен, и желающие выжить охотничьи группы должны были поддерживать минимальный уровень кооперации, чтобы выслеживать раненых животных, разделывать их, транспортировать мясо и затем распределять добычу. Общественная охота приносила в то время наибольшую выгоду и представляла собой хорошо спланированный загон животных (использовались загородки из камня и дерева, естественные густые заросли и скалы, и даже рампы), чтобы они оказались в заранее приготовленной ловушке или – возможно, наиболее простое и хитроумное решение – упали с обрыва (Frison 1987). Многие крупные травоядные – мамонты, бизоны, олени, антилопы, горные козлы – могли быть убиты таким образом, обеспечивая людям запас замороженного или обработанного (закопченного или превращенного в пеммикан) мяса.
Обрыв Хед-Смэшт-Ин-Баффало-Джамп около Форт Маклеод (Альберта, Канада), часть Всемирного наследия ЮНЕСКО, может служить ярким примером такой охотничьей стратегии, поскольку использовался около 5700 лет. «Чтобы начать охоту… молодой человек… мог заманить стадо в нужном направлении, имитируя голос потерявшегося теленка. Как только бизоны подходили к заграждениям для охоты (длинные линии пирамидок из камней, которые складывали, чтобы помочь охотникам направить бизонов к обрыву), охотники появлялись позади стада и пугали животных криками и взмахами рук», и в конечном итоге стадо падало с обрыва (UNESCO 2015а). Полезная энергоотдача от белка и жира, полученного из животных, была очень велика. Охотники позднего плейстоцена могли быть очень успешными, у многих ученых-исследователей Четвертичного периода появилась гипотеза, что именно охота большей частью (или полностью) стала причиной сравнительно быстрого исчезновения мегафауны позднего палеолита, животных с весом тела больше 50 кг (Martin 1958, 2005; Fiedel and Haynes 2004), но она до сих пор окончательно не подтверждена (примечание 2.4).
Примечание 2.4. Исчезновение мегафауны позднего плейстоценаНепрерывное истребление медленно размножающихся животных (с единственным отпрыском, который рождается после долгого вынашивания) может привести к их исчезновению. Если мы примем, что люди позднего плейстоцена имели дневную потребность в пище в 10 МДж/на человека, что они ели в основном мясо и что большая его часть (80 %) обеспечивалась как раз мегафауной, тогда популяции в два миллиона человек требовалось почти 2 Мт (вес в свежем виде) мяса в год (Smil 2013а). Если мамонты были единственным видом, на который охотились, то их требовалось истреблять 250–400 тысяч голов в год. Но охота на мегатравоядных касалась и других крупных млекопитающих (слоны, гигантские олени, бизоны, дикие быки), и потребность в 2 Мт мяса в год могла обеспечить уничтожение около двух миллионов разных животных в год. Наиболее вероятным объяснением исчезновения мегафауны позднего плейстоцена может служить комбинация естественных (изменения климата и растительного покрова) и антропогенных (охота и огонь) факторов (Smil 2013а).
Все до-сельскохозяйственные общества характеризовались всеядностью, составляющие их люди не могли позволить себе роскошь игнорировать какие-либо из доступных источников питания. Хотя охотники-собиратели употребляли в пищу множество видов растений и животных, обычно несколько основных определяли рацион. Неизбежно предпочтение оказывалось семенам – помимо того, что их легко собирать и хранить, в семенах сочетается высокое содержание энергии со сравнительно высоким содержанием белка. Семена диких растений содержат столько же энергии, сколько и зерна культивируемых (пшеница – 15 МДж/кг), у орехов плотность энергии до 80 % больше (грецкий орех – 27,4 МДж/кг).
Мясо любых диких животных – прекрасный источник протеина, но чаще всего оно содержит мало жира и поэтому имеет низкую плотность энергии, вполовину меньше по сравнению с зерном в случае маленьких, жилистых млекопитающих. Ничего удивительного, что широко распространено предпочтение охотиться на крупных и сравнительно жирных животных. Единственный небольшой мамонт давал столько же годной в пищу энергии, сколько 50 северных оленей, а бизон равнялся 20 оленям (примечание 2.5). Именно по этой причине наши неолитические предки устраивали засады на огромных мамонтов, вооружившись простым оружием с каменными наконечниками, и по этой же причине индейцы равнин Северной Америки тратили много энергии на преследование бизонов (чтобы сделать потом из жирного мяса способный долго храниться пеммикан).
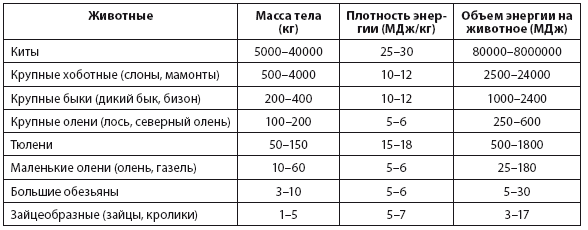
Примечание 2.5. Масса тела, плотность энергии и содержание энергии в пище в зависимости от видов добываемых на охоте животныхПримечание: я предполагаю, что в пищу пригодны две трети массы тела китов и тюленей и половина массы тела других животных. Средняя плотность энергии для китов была рассчитана с учетом того, что 25 % массы их тела составляет ворвань.Источники: основано на данных: Sanders, Parsons and Santley (1979), Sheehan (1985) и Medeiros and co-workers (2011).
Но подход с точки зрения энергии не может дать нам полного понимания, как вели себя охотники-собиратели. Если бы принимались во внимание только энергетические соображения, тогда оптимальная стратегия выглядела бы так: собиратели и охотники пытаются максимизировать объем полученной энергии, одновременно минимизируя затраты времени и сил на ее получение (Bettinger 1991). Оптимальный способ добывания пищи объясняет предпочтение охотиться на крупных жирных мамонтов или собирать менее питательные растения, которые не требуют обработки, потому что орехи с высокой плотностью энергии трудно вскрыть. Вне всякого сомнения, многие охотники-собиратели так и вели себя, максимизируя полезную энергоотдачу, но другие экзистенциальные императивы часто действовали в обратном направлении. Среди наиболее важных – доступность ночного убежища, необходимость защищать территории от конкурентов и потребность в надежных источниках воды, а также витаминах и минералах. Пищевые предпочтения и отношение к работе тоже имели большое значение (примечание 2.6).
Наша неспособность реконструировать баланс энергии в доисторическую эпоху провоцирует появление недопустимых обобщений. Для некоторых групп общие усилия по добыванию пищи были сравнительно низкими, всего несколько часов в день. Подобные факты ведут к тому, что охотников-собирателей описывают как «исходно зажиточные общества», живущие в условиях материального изобилия, лени и праздности (Sahlins 1972). Самый известный случай – племя Добе! Кунг из пустыни Калахари (Ботсвана), процветавшее на диких растениях и мясе. Это племя считали прекрасным примером доисторических охотников-собирателей, живших вроде бы довольной, здоровой и энергичной жизнью (Lee and DeVore 1968). Это заключение, базирующееся на ограниченных и ненадежных данных, должно было – и подверглось сомнению (Bird-David 1992; Kaplan 2000; Bogin 2011).
При упрощающем теоретизировании по поводу процветающих охотников-собирателей не берется во внимание то, насколько тяжела и опасна работа по собиранию пищи и насколько часто стрессовые ситуации и инфекционные болезни посещают примитивные общества. Сезонная нехватка пищи провоцировала потребление негодных в пищу тканей растений и вела к потере веса, а часто к опустошительным эпидемиям. Результатом также была высокая детская смертность (включая детоубийство), а следовательно, низкий уровень воспроизводства. Ничего удивительного, что повторный анализ оценок энергетических затрат и демографических данных, собранных в 60-е, показал, что питание и здоровье Добе! Кунг выглядели в лучшем случае нестабильными, а в худшем говорили об «обществе на грани исчезновения» (Bogin 2011, 349). Как писал другой исследователь (Froment 2001, 259): «Сражающиеся с угрозами и несущие тяжелую ношу болезней, охотники-собиратели не живут – и никогда не жили – в Эдемском саду; они вовсе не богаты, они бедны, их потребности и возможности их удовлетворить ограничены».
Примечание 2.6. Пищевые предпочтения и отношение к трудуПищевые предпочтения убедительным образом демонстрируют различия между схожими во всем остальном группами охотников-собирателей. Племя! Кунг Басарва (Ботсвана) обязано своей популярности в антропологической литературе тем, что зависит от питательных и часто встречающихся орехов монгонго, которые обеспечивают лучший уровень энергоотдачи из задокументированных объектов собирательства. Но /Аисе, другая группа басарва, имеющая доступ к орехам, не употребляет их в пищу, поскольку находит вкус монгонго отвратительным (Hitchcock and Ebert 1984). Схожим образом, обитатели морского берега южной Австралии получают пищу с высокой плотностью энергии с помощью рыболовства, но через пролив от них, на Тасмании в отходах ее прежних обитателей не обнаруживают рыбьей чешуи (Taylor 2007).Прекрасный пример вариативности культурной реальности – упрощенные энергетические модели (Lizot 1977), где сравниваются две обитающие рядом группы индейцев яномами (северная Амазония). Группа, живущая в лесу, потребляла менее половины количества энергии животной пищи и белка, чем их соседи, живущие в среде, хуже обеспеченной дикими свиньями, тапирами, и обладающие такими же охотничьими инструментами и навыками. Объяснение следующее: люди первой группы были просто намного более ленивыми, охотились редко и предпочитали есть меньше. «Одну из недель… мужчины не охотились ни разу, они только собирали их любимые галлюциногены (Anadenanthera peregrina) и проводили целые дни, употребляя наркотики; женщины жаловались, что мяса нет, но мужчины их не слушали» (Lizot 1977,512).Это общий случай основной вариации энергии, добываемой с помощью охоты, при котором вариация не связана ни с доступностью ресурсов (наличие животных), ни с затратами энергии во время охоты (учитывая простое, одинаковое оружие), но является только функцией отношения к труду. Другой пример действий, не имеющих энергетического объяснения, приходит из анализа данных по дележу мяса среди танзанийских хадза (Hawkes, O'Connell and Jones 2001). Лучшее объяснение широко распространенному дележу мяса больших животных – снизить риск, неизбежный при охоте на крупную дичь, но дележ хадза не мотивирован снижающей риск взаимностью, он совершается в первую очередь для повышения статуса охотника как желаемого соседа.
Примерные расчеты для небольшого количества групп охотников-собирателей XX века показывают наиболее высокий возврат полезной энергии при собирании некоторых корней. На одну затраченную единицу энергии в этом случае приходится 30–40 единиц энергии от полученной пищи. И наоборот, охотничьи вылазки, особенно за маленькими древесными или наземными млекопитающими в тропических дождевых лесах, ведут к потере полезной энергии или в крайнем случае имеют нулевую эффективность (примечание 2.7). Типичный выигрыш при собирательстве – в 10–20 раз, тот же самый, что при охоте на больших млекопитающих. В доисторические времена он был, без сомнений, выше, поскольку экосистема содержала больше биомассы, что и определило постепенное увеличение сложности общества.
На самом деле многие общества охотников-собирателей достигали уровня сложности, обычно ассоциируемого с более продвинутыми сельскохозяйственными обществами. Они жили в деревнях с высокой плотностью населения, у них были солидные запасы провианта, социальная стратификация, разработанные ритуалы, зарождалась культивация злаков. Охотники на мамонтов верхнего палеолита на территории Моравского лёссового региона могли похвастаться умело построенными каменными домами, изготавливали множество отличных инструментов и могли обжигать глину (Klima 1954). Социальная сложность групп, населявших в верхнем палеолите юго-запад Франции, определялась сильным атлантическим влиянием, что означало прохладное лето, но исключительно мягкие зимы. Благодаря такой погоде удлинялся сезон вегетации и повышалась продуктивность наиболее южной на континенте тундры, покрытой характерной для степи растительностью и способной поддерживать стада травоядных, более многочисленные, чем где-нибудь еще в ледниковой Европе (Mellars 1985). Сложность этих палеолитических культур лучше всего доказывается их замечательными скульптурами, резьбой и рисунками на стенах пещер (Grayson and Delpech 2002; French and Collins 2015) (рис. 2.3).
Примечание 2.7. Возврат полезной энергии при охоте и собирательствеЯ использую метод, описанный во примечании 1.10, и предполагаю, что люди доисторической эпохи были меньше современных (средний вес взрослого – около 50 кг). Это означает, что для базового метаболизма было нужно около 6 МДж/кг (около 250 кДж/ч) и минимальная потребность в энергии для взрослого составляла 8 МДж, грубо – 330 кДж/час. Для собирания растений требовался в основном свет, чтобы труд был успешным; охота и рыбная ловля предполагали физическое напряжение разной степени. Типичные формы деятельности по добыче пищи требовали четырехкратных затрат по сравнению с базовым метаболизмом для мужчин, и пятикратных – для женщин, или почти 900 кДж/кг. Если отнять базовые сущностные потребности, то вложение полезной энергии будет около 600 кДж/ч. Энергетическая отдача – просто размер пригодной для употребления порции собранных растений или мяса.

Рисунок 2.3. Сделанные углем рисунки на стенах пещеры Шове в Южной Франции. Эти замечательные изображения датируют периодом между 32 900 и 30 000 лет назад (Corbis)
Самые высокие показатели продуктивности в сложном процессе охоты и собирательства связаны с эксплуатацией водных ресурсов (Yesner 1980). Раскопки мезолитических стоянок в южной Скандинавии показали, что когда охотники постледниковой эпохи истребляли стада крупных травоядных, они начинали охотиться на дельфинов и китов, ловить рыбу и собирать морских моллюсков (Price 1991). Они жили в больших, часто постоянных поселениях, рядом с которыми располагались кладбища. Племена тихоокеанского северо-запада, зависящие от рыбной ловли, жили поселками по несколько сотен человек, сооружали качественные деревянные дома. Регулярный нерест лососевых гарантировал надежный и доступный источник пищи, которую было легко запасать (коптить), что позволяло хорошо питаться. Благодаря высокому содержанию жира (около 15 %) мясо лососевых рыб имеет высокую энергетическую плотность (9,1 МДж/кг), и это почти в три раза больше, чем у трески (3,2 МДж/кг). Превосходный образец высокой плотности населения, зависящего от морской охоты – эскимосы северо-западной Аляски. При охоте на мигрирующего гладкого кита они добивались возврата полезной энергии в двухтысячекратном размере (Sheehan 1985; примечание 2.8).
Пищевая стратегия, которая базируется на нескольких сезонных потоках энергии, требует умения хранить пищу. Способы хранения включают зарывание в вечную мерзлоту, сушение и копчение морепродуктов, ягод и мяса; складирование семян и корней, погружение в масло, изготовление колбас, муки из орехов и зерна. Крупномасштабное, долговременное хранение пищи изменило отношение охотников и собирателей к времени, труду, природе, а также помогло стабилизировать высокую плотность населения (Hayden 1981; Testart 1982; Fitzhugh and Habu 2002). Умение планировать и управлять временем стало, возможно, наиболее важным эволюционным преимуществом. Новый способ существования предотвращал частые перемещения и предлагал другой способ выживания, основанный на накоплении излишков. Процесс был саморасширяющимся: попытка манипуляции еще большей долей потоков солнечной энергии вывело общества на путь к еще более высокой сложности.
Примечание 2.8. Китобои АляскиМенее чем за четыре месяца прибрежной охоты на гладкого кита, чьи пути миграции проходят вдоль берега Аляски, люди в умиаках (лодки из плавучего дерева или китовой кости, обтянутые тюленьей шкурой и вмещающие до восьми человек) добывают пропитание для поселений, в которых до контакта с европейцами обитало до 2600 человек (Sheehan 1985; NcCartney 1995). Крупнейшие взрослые киты могут весить до 55 тонн, но даже самые обычные двухлетние животные достигают 12 тонн. Высокая плотность энергии ворвани (около 36 МДж/ кг) и муктука (кожа и ворвань, в которой также содержится витамин С в количестве, сравнимом с тем, что есть в грейпфруте) обеспечивает выигрыш энергии более чем в 2000 раз.Не такую большую, но все равно замечательно высокую отдачу энергии дает эксплуатация ежегодного нереста лососевых прибрежными племенами тихоокеанского северо-запада. Плотность рыбы, идущей вверх по течению, порой столь высока, что рыбаки могут просто вычерпывать ее в лодки или на берег. Столь высокая энергоотдача позволяет существовать крупным постоянным поселениям, характеризующимся социальной сложностью и наличием искусства (большие деревянные тотемы). Окончательные пределы росту населения этих прибрежных поселков устанавливала необходимость охотиться на другие виды морских животных. и эксплуатировать участок земли, чтобы добывать материалы для одежды, строительства и охотничьего снаряжения.
Хотя наше понимание эволюции гоминин значительно улучшилось за последние два поколения, ключевые моменты остаются не проясненными: вопреки всем популярным утверждениям по поводу пользы палеолитической диеты, мы все еще не можем восстановить репрезентативную структуру питания досельскохозяйс-твенной эпохи. Это совсем не удивительно (Henry, Brooks and Piperno 2014). Быстро разлагающиеся остатки растительной пищи очень редко сохраняются на десятки тысяч лет и почти никогда – на миллионы, поэтому исключительно трудно определить долю растений в типичном рационе. Кости сохраняются чаще, но их залежи могут остаться не только от гоминин, но и от хищников, что нужно тщательно отличать, и даже если это сделать, то невозможно понять, насколько репрезентативны они для конкретного рациона питания.
Как отмечают исследователи (Pryor and co-workers 2013), широко распространенный образ обитателей верхнего палеолита Европы как умелых охотников на крупных млекопитающих, населявших большей частью безлесные ландшафты, базируется на плохой сохранности растительных останков на древних стоянках. Работа упомянутых авторов показала, что потенциал таких стоянок в том, что касается макроокаменелостей растений, съеденных людьми, недооценивался и что «способность эксплуатировать растительные источники пищи могла быть важным компонентом в успешной колонизации холодных экосистем Европы» (Pryor et al. 2013, 971). Другие ученые (Henry, Brooks and Piperno 2014) анализировали микроостатки растений – крахмальные зерна и фитолиты, – сохранившиеся в зубном кальции и на каменных орудиях, после чего сделали вывод, что и анатомически современные люди, и их соседи неандертальцы употребляли в пищу разнообразные растения, включая корневища и семена.
Изменения размеров и массы тела, а также строения черепа (грацилизация челюстного аппарата) косвенным образом указывают на основной рацион питания и могут быть вызваны различными сочетаниями видов пищи. Находки каменных инструментов, используемых для убийства и разделки животных, нельзя прямо связать со средним потреблением мяса на человека в широкие периоды времени. Поэтому только прямые показатели изотопного анализа (пропорция 13С/12С и 15N/14N) обеспечивают точные сведения по поводу долгосрочных источников белка, их трофического уровня, а также происхождения (наземного или морского). Этот же способ помогает различить фитомассу, синтезированную двумя принципиально разными способами (С3 и С4), и гетеротрофные организмы, питающиеся такой фитомассой. И все это вместе дает нам сведения о базовом содержании рациона. Но даже подобные исследования нельзя трансформировать в надежные паттерны потребления питательных веществ (углеводов, протеинов, липидов). Все же мы можем сделать вывод, что в граветтский период в Европе мясо было главным источником протеина в пище и морские виды вносили около 20 % от общего количества, а в прибрежных местностях даже больше (Hublin and Richards 2009).
Прежде чем завершить тему энергии в жизни древних людей, я должен заметить, что охота и собирательство продолжали играть важную роль во всех ранних сельскохозяйственных обществах. В Чатал-Гуюке, большом неолитическом сельскохозяйственном поселении в Турции, которое датируется примерно 7200 д. н. э, жители питались в основном семенами и дикими растениями, но при раскопках находят и кости животных от диких быков до лис, барсуков и зайцев (Atalay and Hastorf 2006). В Тель Абу-Хурейра в северной Сирии охота оставалась важным источником пищи через тысячу лет после начала одомашнивания растений (Legge and Rowley-Conwy 1987). Египет до фараонов (ранее 3100 года до н. э.) мог похвастаться выращиванием двузерной пшеницы и ячменя, но население также охотилось на водоплавающих птиц, антилоп, диких свиней и слонов (Hartmann 1923; Janick 2002).
Назад: 2. Энергия в доисторические времена
Дальше: Истоки сельского хозяйства

