Исследование № 16
Как превратить простой лист в сложный
Отбор работает с наследственной изменчивостью, с генетическим разнообразием, которое возникает и поддерживается благодаря мутациям (в самом широком смысле этого слова), а также перекомбинированию генетических вариантов (рекомбинации). То, что основой для дарвиновской эволюции служит генетическая изменчивость, – утверждение вполне тривиальное. Но, как и многие другие тривиальные утверждения общего характера, оно остро нуждается в наглядных примерах и детализации. Мы рассмотрим несколько исследований, в которых разбирается, какие мутации оказываются “интересны” отбору, какие из них ведут к формообразованию, к становлению новых фенотипов, в том числе – более сложных, чем предковые. Расшифровка генетических основ эволюции сложных признаков – задача технически крайне трудная, сегодня это передний край науки. В первом примере генетики из Великобритании, Германии и США работали с модельным растением резуховидкой Таля (Arabidopsis thaliana). Исследование показало, с помощью каких мутаций можно превратить обычные простые листья резуховидки в сложные. Оказалось, что для этого достаточно добавить в геном один-единственный ген RCO (Reduced COmplexity), позаимствованный у другого растения – сердечника шершавого (Cardamine hirsuta). Ген RCO экспрессируется в основаниях мелких выступов по краям зачатка листа и тормозит деление клеток в промежутках между выступами, что в итоге делает лист лопастным или перистым. Ген RCO появился у предков резуховидки и сердечника в результате дупликации древнего гена LMI1, регулирующего развитие цветов и листьев у многих цветковых растений. Приобретение гена RCO было сопряжено с усложнением формы листа, а его последующая утрата некоторыми видами, такими как A. thaliana, – с вторичным упрощением.


Расшифровать во всех деталях генетические основы эволюции сложного морфологического признака – задача исключительно трудная даже для современного уровня развития науки. Каждый новый успех на этом пути – важное достижение, достойное освещения на страницах самых престижных научных журналов.
В 2014 году международная команда генетиков сумела идентифицировать ген, сыгравший важную роль в эволюции формы листа у растений семейства капустных, или, как их называли раньше, крестоцветных (Vlad et al., 2014). Главным объектом исследования был сердечник шершавый (Cardamine hirsuta). Лист у сердечника сложный, перистый, состоящий из нескольких боковых вторичных листочков и одного концевого (рис. 16.1, слева). Растения обрабатывали мутагенным веществом – этилметансульфонатом. У одного из полученных мутантов произошло радикальное упрощение формы листьев: вторичные листочки редуцировались до небольших выступов по краю основного листа и пары лопастей у основания (рис. 16.1, справа). При этом число первичных листьев и их расположение остались прежними.
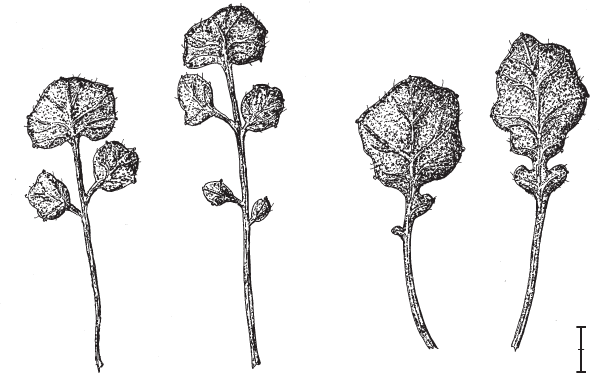
Итак, есть ген RCO, от которого зависит форма листьев, и он появился в результате дупликации другого гена, тоже работающего с формой листьев и цветов. У ранних представителей группы дупликация еще не произошла, поэтому листья у них простые. Но и у резуховидки Таля (Arabidopsis thaliana) листья тоже простые, а ген RCO отсутствует, хотя она не относится к базальным представителям группы. При этом у близкого вида, A. lyrata, ген RCO есть, а листья более сложные, лопастные. Вся совокупность данных говорит о том, что вид A. thaliana с простыми листьями и без гена RCO произошел от предков, у которых ген RCO был, а листья имели более сложное строение.
Вроде бы все ясно, но если вдуматься, то сущность новоприобретения остается туманной. Что это за ген RCO? Каким образом он делает лист более сложным? Чем RCO отличается от своего “исходника”, гена LMI1, и почему у него возникла новая функция? Это как раз те вопросы, которые следует задать, чтобы понять ход эволюционных преобразований. Общие принципы более или менее понятны уже давно, а сегодня нам нужна конкретика. Так что на следующем этапе исследования Даниела Влад и ее коллеги занялись выяснением этой конкретики: что именно делает новый ген RCO? Изначально ученые отталкивались от прочтенной нуклеотидной последовательности RCO: этот ген, будучи дубликатом LMI1, со временем накопил отличия как в белок-кодирующей, так и в регуляторной области. Проще всего было предположить, что эти отличия не имеют значения (то есть являются нейтральными мутациями), а дело просто в увеличении производства белка, кодируемого геном LMI1 и его дубликатом. Исследователи проверили эту гипотезу, создав трансгенные растения с дополнительными копиями гена LMI1. Это ничего не дало: ни у сердечника с мутацией rco, ни у резуховидки Таля форма листьев не усложнилась. Значит, RCO – не просто еще одна копия LMI1. Он обладает своей спецификой, которая может быть заключена как в строении кодируемого белка (то есть в белок-кодирующей части гена), так и в том, где и когда ген включается (что определяется его регуляторной областью). Следовательно, нужно изучить отдельно работу регуляторных и белок-кодирующих участков гена RCO.
Сначала изучили работу регуляторных участков LMI1 и RCO в зачатках листьев сердечника и резуховидки. Чтобы удобнее было регистрировать экспрессию этих генов, к их регуляторным участкам присоединили бактериальный ген, включение которого окрашивает клетки в голубой цвет. Оказалось, что области экспрессии, определяемые регуляторными участками двух генов, сильно различаются. LMI1 экспрессируется на концах будущих вторичных листочков, которые закладываются как небольшие выступы на краю развивающегося листа, а RCO – в основаниях этих выступов (илл. II, см. цветную вклейку). При этом LMI1 экспрессируется также в зачатках цветов и прилистников, тогда как работа RCO приурочена исключительно к основаниям вторичных листочков сложного листа. Области экспрессии LMI1 похожи у разных крестоцветных, включая и упомянутый базальный вид – крылотычинник с простыми листьями.
Таким образом, отличия в регуляторном участке гена RCO, накопленные после дупликации, изменили область его экспрессии по сравнению с исходной (предковой), характерной для LMI1. Это указывает на то, что RCO приобрел новую функцию (произошла неофункционализация).
Затем ученые проверили, влияют ли на новую функцию гена RCO особенности его белок-кодирующей части. Они проделали поистине ювелирную работу, присоединив кодирующую часть LMI1 к регуляторной части RCO и вставив эту конструкцию в геном сердечника с мутацией rco (как мы помним, у сердечника с этой мутацией сложные листья превращаются в простые). В результате простые листья растений-мутантов снова стали сложными. Следовательно, для формирования сложных листьев важны только особенности регуляторной части RCO, а белок-кодирующая часть срабатывает и в старом варианте. Стало быть, белки, кодируемые генами RCO и LMI1, сами по себе остались взаимозаменимыми. Все дело в том, что появились новые области экспрессии (в основаниях выступов листа) вдобавок к старым (на внешнем краю листовой пластинки).
И вот пришло время решающего эксперимента. Если предположения исследователей верны и листья у A. thaliana упростились в связи с потерей гена RCO, то возвращение утраченного гена должно привести к тому, что листья любимого цветочка генетиков усложнятся. Эксперимент был проведен – и дал положительный результат (рис. 16.2).
Правда, листья у трансгенных A. thaliana не стали такими же сложными, как у сердечника C. hirsuta. Вторичные листочки не сформировались, вместо них получились лопасти. По-видимому, чтобы лопасти стали полноценными вторичными листочками с собственными черешками, помимо RCO нужно что-то еще – какие-то дополнительные генетические особенности, имеющиеся у сердечника и отсутствующие у резуховидки. Интересно, что точно такие же лопастные листья, как у трансгенных A. thaliana, характерны для родственного вида A. lyrata, в геноме которого, как мы уже упоминали, сохранился ген RCO.
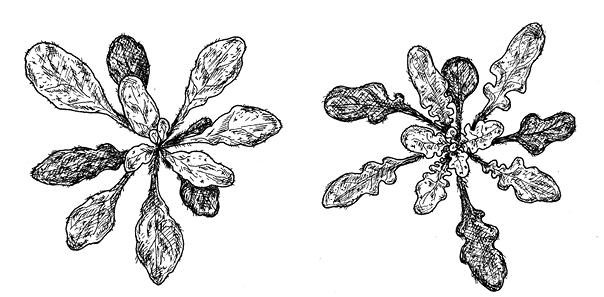
рис. 16.2. Ген RCO, вставленный в геном Arabidopsis thaliana, превращает простые листья в лопастные. Слева – обычное растение A. thaliana, справа – трансгенное. Длина масштабной линейки – 1 см.
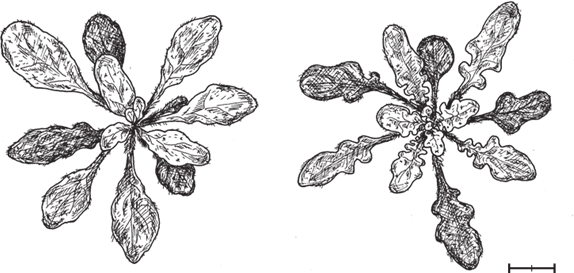
Дальнейшие опыты показали, что различные варианты гена LMI1, взятые у разных представителей крестоцветных (и даже у некоторых других двудольных), вызывают формирование лопастных листьев у A. thaliana, если белок-кодирующие участки этих генов объединить с регуляторным участком RCO. Это еще один довод в пользу того, что ключевое эволюционное новшество при формировании сложных листьев у крестоцветных состояло в изменении регуляторного участка RCO, тогда как свойства белков, кодируемых этими генами, весьма консервативны.
Наконец, ученые попытались выяснить, каким образом ген RCO со своим специфическим регуляторным участком превращает простой лист с волнистым или зазубренным краем в лопастной или перистый. Для этого потребовалось внимательнее приглядеться к тому, что именно делает RCO, как он влияет на развитие листа. Исследователи сравнили рост листовых зачатков у обычного сердечника с перистыми листьями и у мутантов rco с простыми листьями. В обоих случаях на краю листовой пластинки формируются небольшие округлые выступы. Дальнейший рост этих выступов протекает одинаково у обычных C. hirsuta и у мутантов rco. Однако в поведении клеток, расположенных в промежутках между выступами, наблюдаются контрастные различия. В норме у C. hirsuta эти клетки не делятся. Поэтому между растущими выступами формируются глубокие вырезы, а сами выступы в итоге становятся вторичными листочками. У мутантов rco клетки в промежутках между выступами активно делятся, поэтому край листового зачатка растет равномерно. В результате вместо перистого листа получается сплошная листовая пластинка с волнистым краем. По-видимому, это значит, что ген RCO, экспрессирующийся в основаниях выступов, не столько ускоряет рост самих выступов, сколько подавляет деление клеток в промежутках между зонами своей экспрессии.
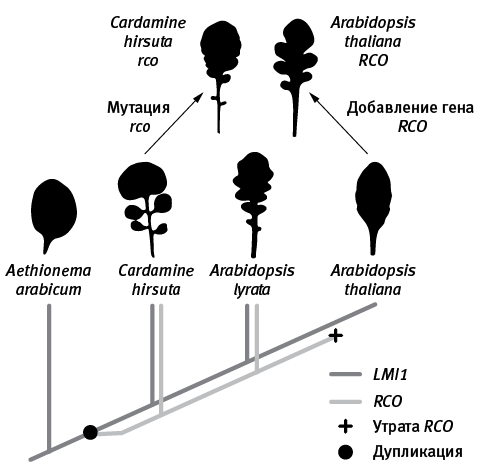
рис. 16.3. Эволюция формы листа у крестоцветных. Внизу – эволюционное дерево генов LMI1 (темно-серые линии) и RCO (светло-серые). У базальных крестоцветных (Aethionema arabicum) не было гена RCO, а листья были простыми. У общего предка Cardamine и Arabidopsis произошла дупликация гена LMI1 (черный кружок). Одна из двух копий удвоившегося гена сохранила старую функцию, а другая (RCO) приобрела новую благодаря изменениям регуляторного участка, что способствовало усложнению формы листьев. Впоследствии утрата RCO (черный крестик) привела к вторичному упрощению листьев у A. thaliana. Поломка гена RCO у C. hirsuta (мутация rco) ведет к упрощению листьев. Возвращение утраченного гена RCO в геном A. thaliana приводит к усложнению формы листа. По рисунку из Vlad et al., 2014.
Таким образом, детали головоломки постепенно сложились в целостную картину (она схематично изображена на рис. 16.3). У первых крестоцветных, унаследовавших ген LMI1 от своих предков и еще не имевших RCO, листья были простыми, как у крылотычинника (Aethionema). Затем произошла дупликация LMI1 с последующей неофункционализацией одной из копий. Новая функция возникла благодаря изменению регуляторной части гена, что повлияло на область его экспрессии. Ген начал экспрессироваться в основаниях выступов развивающейся листовой пластинки. Это затормозило деление клеток в промежутках между выступами и привело к усложнению формы листа. В дальнейшем предки A. thaliana утратили ген RCO, что вызвало вторичное упрощение листа (как у C. hirsuta с мутацией rco). Однако в геноме A. thaliana сохранились все прочие необходимые компоненты программы развития сложного листа, не хватает лишь гена RCO (или любого похожего гена, управляемого регуляторной областью RCO). Если его искусственно вставить в геном A. thaliana, растение приобретает сложные лопастные листья, в точности как у родственного вида A. lyrata.
Скорее всего, тот же самый эффект мог быть достигнут и без генной дупликации – путем формирования в регуляторной области LMI1 новых энхансеров (так называют регуляторные участки гена, к которым прикрепляются специальные белки – регуляторы транскрипции, или транскрипционные факторы, – что приводит к усилению или, наоборот, ослаблению экспрессии гена). Именно таким способом развивались, например, узоры на крыльях дрозофил (о чем мы рассказывали в книгах “Рождение сложности” и “Эволюция. Классические идеи в свете новых открытий”). В обоих случаях ключевую роль сыграли изменения регуляторных, а не кодирующих участков генов. По-видимому, это самый типичный путь формирования новых морфологических признаков. Гены, контролирующие развитие организма, как правило, многофункциональны. Причина в том, что в ходе эволюции к ним очень легко “привешиваются” новые функции, ведь фактически эти гены служат профессиональными регуляторами активности других генов – и им все равно, какие именно гены регулировать. Многофункциональность регуляторов развития накладывает жесткие ограничения на эволюцию их белок-кодирующих частей, потому что изменения в них приведут к множественным последствиям, большинство из которых наверняка будут вредными. Обойти эту проблему позволяют изменения регуляторных участков таких генов. При этом формируются новые области экспрессии гена-регулятора, что позволяет создать новую функцию, не вредя старым.


