Исследование № 1
Есть ли предел приспособленности?
Знаменитый эволюционный эксперимент на бактериях, начатый Ричардом Ленски в 1988 году, продолжает приносить интересные и порой неожиданные результаты. С начала эксперимента сменилось уже более 70 000 поколений подопытных бактерий Escherichia coli (у людей на это ушло бы около двух миллионов лет). Казалось бы, все возможные полезные мутации за это время должны были у бактерий закрепиться, но нет, микробы в колбах у Ленски продолжают накапливать полезные мутации. Их приспособленность к неизменным условиям эксперимента неуклонно повышается. И хотя она растет уже не так быстро, как в начале эксперимента, рост не собирается выходить на плато, как ожидали многие эксперты. В шести из двенадцати популяций закрепились мутации, резко повысившие темп мутагенеза, что лишь ускорило рост приспособленности, несмотря на то что от мутаций, как известно, в среднем намного больше вреда, чем пользы.


Уникальный эксперимент, начатый в 1988 году Ричардом Ленски из Мичиганского университета (США), позволяет с небывалой степенью детальности следить за эволюцией бактерий в реальном времени. Эксперимент проводится параллельно с дюжиной популяций кишечной палочки (Escherichia coli). Все популяции изначально были одинаковыми – их получили от одного и того же предкового штамма. Бактерий выращивают на “минимальной” питательной среде, в которой размножение бактерий сдерживается недостатком пищи (глюкозы). Каждый день из колбы с микробами берут 0,1 мл содержимого и помещают в новую колбу с 9,9 мл свежей питательной среды. Периодически часть популяции замораживают при –80°C и сохраняют для последующего изучения. Бактерии хорошо переносят заморозку, так что в распоряжении исследователей имеется живая “ископаемая летопись” эксперимента. Это мудро, потому что аналитические методики, в частности методы секвенирования (“прочтения”) геномов, сейчас стремительно развиваются и столь же стремительно дешевеют. Живая “ископаемая летопись” позволяет не только следить за эволюционными событиями, но и проигрывать те или иные события повторно, чтобы отделить случайности от закономерностей. Регулярно проводится оценка приспособленности популяций к тем условиям, в которых их содержат. Для этого сравнивают скорости размножения подопытных микробов и контрольного (предкового) штамма, который тоже, конечно же, бережно хранится в замороженном виде.
К 2013 году в колбах сменилось более 59 000 поколений микробов (каждые 75 дней сменяется примерно 500 поколений). Длительность эксперимента и размер популяций были достаточными для того, чтобы каждая из возможных точечных мутаций (нуклеотидных замен) в ходе случайного мутирования произошла более одного раза (размер генома подопытного штамма кишечной палочки – 4,6 × 106 пар нуклеотидов). Пока это единственная в мире экспериментальная система, позволяющая в деталях проследить эволюционные изменения в большой популяции на таком длительном интервале времени.
Система предельно упрощена по сравнению с природными сообществами микроорганизмов. Во-первых, бактерии размножаются в монокультуре, что позволяет абстрагироваться от межвидовых взаимодействий. По крайней мере, Ричард Ленски и его коллеги надеялись, что позволяет (см., однако, Исследование № 3). Во-вторых, питательная среда бедная, в ней мало пищи, поэтому плотность микробного населения в колбах остается низкой. Это минимизирует влияние бактерий друг на друга посредством выделения тех или иных веществ. В-третьих, популяции бесполые: они лишены средств для горизонтальной передачи генов, так что те передаются только вертикально – от родителей потомкам.
С одной стороны, все эти упрощения делают эксперимент несколько оторванным от реальности. С другой же – позволяют получать понятные, однозначно интерпретируемые результаты. Цель эксперимента – изучить самые фундаментальные эволюционные процессы (мутагенез, отбор, генетический дрейф, адаптацию к среде) в чистом, так сказать, виде. “Сложности” можно будет добавлять потом, по мере необходимости, когда станут понятны основы.
В 2013 году Ленски и его коллеги сообщили об очередном важном результате (Wiser et al., 2013). Ученые сосредоточились на росте приспособленности в подопытных популяциях. Приспособленность, напомним, оценивается как скорость размножения бактерий в стандартных условиях эксперимента по сравнению с предковым штаммом, который сохраняется в замороженном состоянии, так что его в любой момент можно разморозить и использовать в опытах.
В трех из двенадцати популяций в ходе эксперимента произошли настолько радикальные эволюционные изменения, что сравнивать их приспособленность с другими линиями стало трудно. В одной популяции развилась способность использовать в пищу цитрат, присутствующий в среде как вспомогательное вещество. Обычные кишечные палочки питаться им не могут. Это привело к резкому росту плотности популяции. Две другие разучились образовывать колонии на агаре. Из-за этого ученые не смогли применить к ним стандартную методику оценки скорости роста. Поэтому поздние этапы эволюции этих трех популяций были исключены из анализа (а ранние учитывались). Между прочим, во всех трех популяциях закрепились так называемые аллели-мутаторы, то есть аллели, несущие мутации, которые повышают темп мутагенеза (см. ниже).
Ранее было показано, что на протяжении первых 20 000 поколений приспособленность росла с замедлением. Что будет дальше? Следует ли ожидать выхода на плато, то есть достижения постоянного уровня приспособленности, или же плато не получится и приспособленность будет продолжать понемногу расти? Этот вопрос – достижим ли вообще предел приспособленности – очень важен для биологов-теоретиков, ведь речь, по сути, идет о границах творческих возможностей эволюции.
Для начала ученые сравнили приспособленность бактерий из поколений № 40 000 и 50 000. Оказалось, что в среднем подопытные популяции за период смены 10 000 поколений повысили свою приспособленность на целых 3 %. Таким образом, даже после 40 000 поколений адаптация не прекратилась: микробы все еще продолжают накапливать полезные мутации и повышать свою приспособленность.
Затем исследователи проследили траекторию роста приспособленности, заставляя замороженных на разных этапах эксперимента микробов соревноваться с предками (рис. 1.1).
Статистический анализ полученных данных показал, что изменения приспособленности соответствуют степенной модели с неограниченным (хотя и замедляющимся) ростом. Это значит, что приспособленность вовсе не собирается выходить на плато. Иными словами, бактерии, по всей видимости, не намерены останавливаться на достигнутом. Несмотря на долгую жизнь в одних и тех же условиях, у бактерий все еще время от времени закрепляются новые мутации, в данных условиях полезные (напомним, что в эволюционной биологии слово “полезный” обычно используется в смысле “повышающий приспособленность”).

рис. 1.1. Рост приспособленности бактерий за 50 000 поколений. Точками (первые 20 000 поколений) и кружочками (следующие 30 000 поколений) показаны усредненные значения по всем подопытным популяциям. По вертикальной оси – относительная приспособленность (скорость роста по сравнению с предковым штаммом). Пунктиром показаны предсказания моделей: гиперболической (приспособленность асимптотически стремится к предельному уровню; нижняя кривая) и степенной (неограниченный, хотя и замедляющийся рост приспособленности). Параметры моделей основаны на данных по первым 20 000 поколений. Видно, что гиперболическая модель хуже предсказывает динамику приспособленности за последующие 30 000 поколений, чем степенная. По рисунку из Wiser et al., 2013.
Удивительная неисчерпаемость эволюционных возможностей, впервые столь наглядно продемонстрированная в эксперименте, имеет огромное значение для понимания эволюции. Ведь можно было предположить (многие так и думали), что в постоянных условиях, тем более в монокультуре, возможности для адаптивной эволюции быстро исчерпаются и система придет в состояние стазиса. В таком случае объяснить продолжающуюся эволюцию жизни на нашей планете можно было бы только непостоянством среды (абиотической и биотической).
По-видимому, запаса потенциально полезных мутаций хватит не только на 50 000 поколений бактерий, но и на 50 000 поколений наблюдающих за ними исследователей. Как ни странно, идущий безостановочно эволюционный процесс не приведет к какому-то невероятному, запредельному росту приспособленности. Ученые рассчитали, что если приспособленность будет и впредь увеличиваться по той же траектории (описываемой степенной моделью), то пятидесятитысячный по счету преемник Ленски увидит бактерий, удваивающих свою численность каждые 23 минуты. Это высокая скорость размножения для бактерий, но не какая-то неслыханная (численность бактерий предкового штамма удваивается за 55 минут).
Откуда же берутся новые полезные мутации? Ведь за время эксперимента каждая возможная нуклеотидная замена уже наверняка успела произойти хотя бы у одной бактерии! Основных причин, по-видимому, две. Первая связана с эпистазом – своего рода взаимодействием между мутациями. Например, мутация Б становится полезной только после того, как бактерия уже обзавелась мутацией А. Здесь нужно помнить еще и о том, что многие полезные мутации имеют негативные побочные эффекты. Мутация А может улучшить работу одной молекулярной системы, немного повредив другой. Допустим, первый эффект сильнее второго, поэтому в суммарном зачете мутация А оказывается “полезной” и поддерживается отбором. Но теперь, когда в популяции закрепилась мутация А, полезной окажется любая компенсаторная мутация Б, сглаживающая негативный побочный эффект мутации А.
Вторая причина неисчерпаемости запаса полезных мутаций состоит в том, что мутации с очень слабым полезным эффектом требуют очень долгого времени для закрепления в популяции. Такая мутация, появившись, с большой вероятностью будет вскоре потеряна из-за генетического дрейфа – случайных колебаний частот аллелей. Чтобы отбор начал хоть немного “помогать” такой мутации распространяться, число ее носителей должно стать довольно большим, а до тех пор она будет находиться целиком под властью безжалостного, неразборчивого дрейфа (см. Исследование № 4). Расчеты показывают, что подавляющее большинство вновь возникающих полезных мутаций теряются. Чтобы данная слабополезная мутация в конце концов все-таки закрепилась, она должна многократно появиться у разных бактерий независимо. Ленски и его коллеги рассчитали, что мутация, повышающая приспособленность бактерий на одну миллионную, должна возникнуть примерно 250 000 раз, прежде чем она наконец распространится и зафиксируется в популяции. Если принять во внимание частоту мутирования подопытных бактерий (в среднем одна мутация на 1010 нуклеотидов за поколение) и размер их популяций (эффективная численность каждой из них, с учетом ежедневных бутылочных горлышек, составляет примерно 33 000 000 клеток), получится, что такой мутации понадобится 100 000 000 поколений, чтобы “спастись” от дрейфа, и еще миллионы поколений, чтобы зафиксироваться (достичь стопроцентной частоты). Таким образом, накопление слабополезных мутаций в эксперименте Ленски вряд ли закончится в обозримом (и даже необозримом) будущем.
Причины замедления роста приспособленности связаны с тем, что каждая закрепившаяся полезная мутация делает последующие генетические усовершенствования в среднем менее полезными. Авторы называют этот эффект “эпистазом убывающей доходности” (по аналогии с экономическим законом убывающей доходности). Иными словами, сначала происходят наиболее радикальные адаптивные изменения, а затем идет все более тонкая настройка и оптимизация фенотипа.
Еще один фактор, способствующий замедлению роста приспособленности и тесно связанный с предыдущим, – клональная интерференция, то есть конкуренция между клонами бактерий с разными полезными мутациями. Напомним, что бактерии, участвующие в эксперименте, – бесполые, они не могут обмениваться генами. Поэтому, если у одной бактерии возникнет одна полезная мутация, а у другой – другая, эти мутации не смогут объединиться в одном геноме (как это произошло бы у нормальных микробов, способных к горизонтальному переносу генов, и уж тем более у организмов, размножающихся половым путем; см. Исследование № 7). Вместо этого потомки первой бактерии начнут не на жизнь, а на смерть (не на свою, конечно, а своей линии) конкурировать с потомками второй. Победит, естественно, тот клон, чья мутация окажется полезнее. Вторая, менее полезная мутация будет вытеснена и потеряна. Придется теперь ждать, пока она случайно появится снова у бактерии, уже имеющей первую мутацию. А закрепиться она сможет, только если у нее не окажется более удачливых конкурентов. Клональная интерференция во многом объясняет, почему на первых этапах эксперимента фиксировались в основном мутации с сильным полезным эффектом (в среднем первая закрепившаяся в каждой популяции мутация повышала приспособленность на 10 % – это очень много), а затем уже постепенно начинали фиксироваться все менее и менее полезные мутации.
Разумеется, у бактерий, способных к горизонтальному переносу генов, адаптация шла бы гораздо быстрее (см. Исследование № 7). Но и анализировать результаты было бы труднее, потому что к мутационной изменчивости добавилась бы комбинативная, а на эволюцию стали бы влиять такие факторы, как наследственные различия по склонности к приему и передаче генов, избирательность при выборе партнеров и т. д.
Еще один важный результат дало сравнение роста приспособленности у популяций, сохранивших исходный (низкий) темп мутагенеза, и тех, где закрепились аллели-мутаторы, несущие мутации, которые резко (в среднем на два порядка) повысили темп мутирования. На сегодняшний день “гипермутаторами” стали шесть из двенадцати популяций.
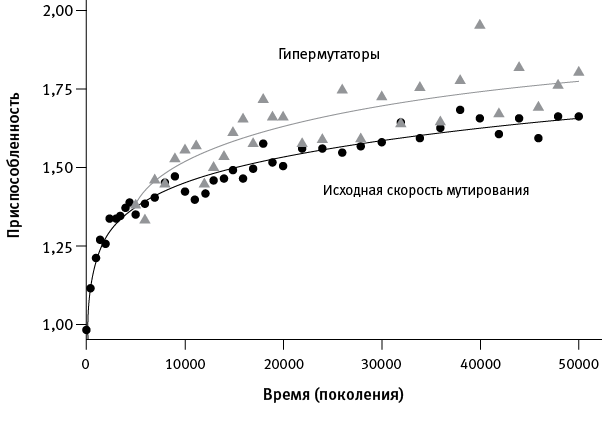
рис. 1.2. Повышение темпа мутагенеза ускоряет адаптацию. Черные точки – усредненные данные по шести популяциям, в которых темп мутагенеза остался на исходном низком уровне. Серые треугольники – усредненные данные по трем популяциям, в которых на ранних этапах эксперимента (за первые 20 000 поколений) закрепились аллели-мутаторы. Видно, что приспособленность у вторых росла быстрее, чем у первых. Еще три популяции, у которых мутаторы закрепились поздно, в данном случае не рассматривались. По рисунку из Wiser et al., 2013.
Оказалось, что у “гипермутаторов” адаптация протекала ускоренными темпами (рис. 1.2). Получается, повышенный темп мутагенеза пошел бактериям на пользу. Это противоречит распространенной идее о том, что в стабильных условиях организмам выгодно снизить темп мутирования до нуля. Ведь большинство вновь возникающих мутаций вредны, поэтому мутация, повышающая темп мутагенеза, в краткосрочной перспективе приносит больше вреда, чем пользы, и отбор, по идее, должен действовать против нее.
Почему же аллели-мутаторы все-таки распространяются? Дело в том, что в бесполой популяции они часто спасаются от отбраковки благодаря эффекту, который называют генетическим автостопом. Если какой-то гипермутабельной бактерии повезет и у нее появится редкая и очень полезная мутация, отбор начнет поддерживать последнюю. При этом вместе с полезной мутацией будет автоматически распространяться и сидящий в том же геноме аллель-мутатор. Он будет, подобно пассажиру-безбилетнику, ехать прицепом к хромосоме, распространение которой в генофонде популяции “оплачено” полезной мутацией. А все потому, что в бесполой популяции, где нет перетасовки генов и хромосомных участков между отдельными клетками, отбираться могут только целые геномы, но не отдельные гены.
В популяции, способной к генетическому обмену, ситуация будет иной. Связка мутатора с полезной мутацией не будет неразрывной, что позволит отбору работать с ними индивидуально. В итоге отбор сможет закрепить полезную мутацию и отбраковать породивший ее аллель-мутатор (см. Исследование № 7). Но это пока лишь теория: эволюционный эксперимент на организмах, способных к генетическому обмену, сопоставимый по масштабу с экспериментом Ленски, еще не поставлен.
Итак, мы увидели, что предела приспособленности, по всей видимости, нет. Об этом нам говорят экспериментальные данные. Этот вывод кажется контринтуитивным, потому что “высшую точку” приспособленности можно легко себе представить как нечто реальное и достижимое. Казалось бы, для заданных постоянных условий должно существовать некое оптимальное, наиболее приспособленное состояние – такое, что у организма, его достигшего, никакая мутация уже не сможет повысить приспособленность. Даже если для реальной эволюционирующей популяции этот оптимум недостижим, он продолжает оставаться для биологов удобной абстракцией, упрощением, помогающим решать определенные задачи. Эволюционисты привыкли представлять себе этот оптимум в виде горной вершины на воображаемом ландшафте. Но при этом важно понимать, какие пути ведут к высшей точке, или, образно выражаясь, какой рельеф имеют склоны горы. Упрощенное понимание естественного отбора рисует нам гладкие склоны и прямой путь к вершине. Но это, очевидно, не соответствует биологическим реалиям. Благодаря стремительному развитию науки, “путь к вершине приспособленности” постепенно перестает быть абстракцией и начинает поддаваться картированию. Об этом – следующая глава.

