Градации структуры, сопровождаемые сменой функции
Возвратимся теперь к другим возражениям м-ра Майварта. В целях защиты насекомые часто сходны с различными предметами, как, например, с зелеными или опавшими листьями, сухими сучками, кусочками лишайников, цветками, иглами, экскрементами птиц и живыми насекомыми; к последнему я еще возвращусь позднее. Это сходство часто бывает поразительно близким и, не ограничиваясь окраской, распространяется на форму и даже на манеру держать себя. Гусеницы, свешивающиеся неподвижно, подобно сухой веточке, с кустарника, на котором кормятся, служат превосходным примером подобного сходства. Случаи подражания таким предметам, как экскременты птиц, редки и исключительны. По этому поводу м-р Майварт замечает следующее: «Так как, согласно теории м-ра Дарвина, существует постоянная тенденция к неопределенной вариации и так как мельчайшие зарождающиеся вариации происходят во всех направлениях, то они должны нейтрализовать друг друга и с самого начала образовывать такие нестойкие модификации, что трудно и даже невозможно понять, как такие неопределенные колебания бесконечно малых зачатков могут когда бы то ни было привести к заметному сходству с листом, тростником или каким-нибудь другим предметом, настолько, чтобы Естественный Отбор использовал и увековечил их».
Но во всех предыдущих случаях насекомые уже в своем исходном состоянии, без сомнения, обладали некоторым грубым и случайным сходством с каким-либо предметом, обычно встречающимся в их местообитании. Это совсем нельзя считать невероятным, принимая во внимание почти бесконечное количество окружающих предметов и разницу в форме и окраске множества существующих насекомых. Так как грубое сходство необходимо уже с самого начала, нам становится понятным, почему крупные и высокоорганизованные животные (насколько я знаю, за исключением одной рыбы) сходны в целях защиты не с отдельными предметами, а лишь с фоном обычно окружающей их местности, и только по окраске. Допустим, какое-то насекомое первоначально оказалось случайно похожим на сухой сучок или опавший лист и что это насекомое слабо варьирует в разных направлениях; тогда все те вариации, которые делают его более похожим на такой предмет и тем самым благоприятствуют его защите, будут сохраняться, а другие вариации будут находиться в пренебрежении и в конце концов утратятся; если же вариации делают насекомое вообще менее похожим на предмет, которому оно подражает, то они будут элиминированы. Возражение м-ра Майварта имело бы, однако, силу, если бы нам удалось объяснить вышеприведенное сходство, независимо от естественного отбора, посредством простой колеблющейся изменчивости; но действительное положение вещей таково, что это возражение не имеет значения.
Я не могу также придавать значения трудности, которую м-р Майварт усматривает в «последних степенях совершенства мимикрии», как, например, в приводимом м-ром Уоллесом случае с палочковидным насекомым (Сегохуlus laceratus), похожим на «покрытую ползучим мхом, или Jungermannia, палочку». Сходство это так велико, что туземец даяк утверждал, что листоподобные выросты представляют собою действительно мох. Насекомые служат добычей для птиц и других врагов, чье зрение, вероятно, острее нашего, и любая степень сходства, помогающая насекомому не быть замеченным или открытым, способствует его сохранению; а чем сходство полнее, тем лучше для насекомого. Изучая своеобразие различий между видами в той группе, к которой принадлежит вышеупомянутый Ceroxylus, мы не находим ничего невероятного в том, что у этого насекомого подвергались модификации неправильной формы наружные выросты, которые приобретали более или менее зеленую окраску; вообще во всякой группе признаки, различные у разных видов, наиболее склонны варьировать, тогда как родовые признаки, т. е. общие всем видам, наиболее постоянны.
Гренландский кит – одно из самых замечательных животных на свете, и так называемый китовый ус представляет одну из его наиболее выдающихся особенностей. Китовый ус состоит из ряда пластинок, которые, находясь приблизительно в числе 300 на каждой стороне верхней челюсти, расположены очень тесно друг к другу поперек продольной оси ротовой полости. С внутренней стороны главного ряда находится несколько добавочных. Концы и внутренние края всех пластинок расщеплены в крепкие щетины, которые покрывают все огромное нёбо и служат для просасывания или процеживания воды и для задержания мелкой добычи, которой питается это огромное животное. Средняя и самая длинная пластинка гренландского кита достигает 10, 12 и даже 15 футов в длину, но у различных видов Cetacea наблюдаются градации в длине пластинок; по Скорсби (Scoresby), у одного вида средняя пластинка достигает четырех футов, у другого – трех, у третьего – 18 дюймов, у Balaenoptera rostrata – только около девяти дюймов в длину. Качество китового уса также отличается у разных видов.
Относительно китового уса м-р Майварт замечает, что, «однажды достигнув такой величины и такого развития, при котором он вообще стал полезен, он мог сохраняться и увеличиваться в нужных размерах уже путем одного только естественного отбора. Но как могло быть получено самое начало такого полезного развития?». В ответ на это можно спросить, почему бы отдаленные предки китов, обладающих китовым усом, не могли иметь рта, построенного более или менее сходно с клювом утки, снабженным пластинками? Утки, подобно китам, кормятся, процеживая грязь и воду, и это семейство иногда называли Criblatores, т. е. цедильщики. Я надеюсь, что не буду ложно понят в том смысле, будто я говорю, что предки китов имели в действительности рот с пластинками, подобно клюву утки. Я хочу только сказать, что это нельзя считать невероятным и что огромные пластины китового уса гренландского кита могли развиться из таких пластинок путем градуальных мелких шагов, из которых каждый был полезен для их обладателя.
Клюв широконосной утки (Spatula clypeata) по своему строению замечательнее и сложнее, чем рот кита. Верхняя челюсть снабжена с каждой стороны (у исследованного мною экземпляра) рядом или гребнем из 188 тонких эластичных пластинок, срезанных наискось, так что они заострены и расположены поперек продольной оси рта. Они отходят от нёба и гибкой перепонкой прикреплены к бокам челюсти. Находящиеся посредине самые длинные достигают около одной трети дюйма в длину, выдаваясь на 0.14 дюйма из-под края. При их основании находится короткий дополнительный ряд из косо поперечных пластинок. В этом отношении они сходны с пластинками китового уса во рту кита, но к концу клюва они очень сильно отличаются от него, так как направлены внутрь, а не прямо свешиваются вниз. Вся голова широконоски, хотя неизмеримо менее объемистая, составляет около одной восемнадцатой длины головы средней величины Balaenoptera rostrata, у которого китовый ус имеет только девять дюймов длины; следовательно, если бы мы представили себе голову широконоски одной длины с головой Balaenoptera, то ее пластинки имели бы шесть дюймов в длину, т. е. составляли бы две трети длины китового уса у данного вида кита. Нижняя челюсть широконоски снабжена пластинками той же длины, что и верхняя, но более тонкими, и в этом отношении резко отличается от нижней челюсти кита, которая лишена китового уса. С другой стороны, края этих нижних пластинок расщеплены на тонкие щетиновидные выросты, чем они замечательно походят на пластинки китового уса. У рода Prion, принадлежащего к особому семейству буревестников, только верхняя челюсть снабжена пластинками, которые хорошо развиты и спускаются ниже края, отчего клюв этой птицы походит в указанном отношении на рот кита.
От высокоразвитого клюва широконоски мы можем без большого перерыва перейти [как показывают сообщение и образцы, присланные мне м-ром Салвином (Salvin)], насколько это касается приспособления для процеживания, через клюв Merganetta armata и в некоторых отношениях через клюв Aix sponsa к клюву обыкновенной утки. У последнего вида пластинки гораздо грубее, чем у широконоски, и крепко приросли к бокам челюсти; с каждой стороны их всего около 50, и они не спускаются ниже ее края. Квадратные верхушки этих пластинок переходят на краю в прозрачную твердую ткань, годную для размельчения пищи. Края нижней челюсти покрыты многочисленными тонкими поперечными ребрами, которые выдаются очень мало. Хотя клюв при таком устройстве стоит в качестве сита гораздо ниже, чем клюв широконоски, однако эта птица, как всем известно, постоянно пользуется им с этой целью. У других видов, как мне сообщил м-р Салвин, пластинки развиты даже значительно меньше, нежели у обыкновенной утки, но я не знаю, пользуются ли они своим клювом для процеживания воды.
Обратимся к другой группе того же семейства. У египетского гуся (Chenalopex) клюв весьма похож на клюв обыкновенной утки, но его пластинки не так многочисленны, не так резко отделены друг от друга и не так вдаются внутрь; однако этот гусь, как мне сообщил м-р Бартлетт (Bartlett), «пользуется своим клювом как утка, пропуская воду через углы рта». Однако главной пищей ему служит трава, которую он щиплет, подобно обыкновенному гусю. У последней птицы пластинки верхней челюсти гораздо короче, чем у обыкновенной утки, почти сливаются, их около 27 с каждой стороны, и они оканчиваются зубоподобными бугорками. Нёбо также покрыто округлыми бугорками. Края нижней челюсти покрыты зазубринами, выступающими сильнее, более грубыми и более острыми, нежели у утки. Обыкновенный гусь не процеживает воды, но пользуется своим клювом исключительно для того, чтобы щипать или срезывать траву, к чему этот клюв приспособлен так хорошо, что гусь может ощипывать им траву тщательнее почти любого другого животного. У других видов гусей, как мне сообщил м-р Бартлетт, пластинки менее развиты, нежели у обыкновенного гуся.
Таким образом, мы видим, что одни члены семейства утиных с клювом, устроенным подобно клюву обыкновенного гуся, адаптированным исключительно для ощипывания травы, или даже имеющие клювы с менее развитыми пластинками, могут путем небольших изменений преобразоваться в такой вид, как египетский гусь, последний – в вид, подобный обыкновенной утке, а этот, наконец, в вид с клювом широконоски, годным почти исключительно для процеживания воды; последнее подтверждается тем, что эта птица едва ли может употреблять свой клюв, кроме его загнутого кончика, для схватывания и размельчения твердой пищи. К этому я могу прибавить, что путем небольших изменений клюв гуся может превратиться в клюв с выдающимися, загнутыми назад зубцами, подобный клюву Merganser (представитель того же самого семейства), служащему для совершенно особой цели – схватывать живую рыбу.
Вернемся к китам. Hyperoodon bidens лишен настоящих, пригодных для работы зубов, но его нёбо затвердело и снабжено, по Ласепеду (Lacopede), небольшими неравными жесткими роговыми выступами. Поэтому нет ничего невероятного в предположении, что какая-нибудь из ранних форм Cetacea имела нёбо, покрытое подобными же ороговевшими выступами, несколько более правильно расположенными, которые, подобно буграм на клюве гуся, помогали схватывать и размельчать пищу. Но если так, то едва ли можно отрицать, что указанные выступы посредством вариаций и естественного отбора могли превратиться в пластинки, столь же развитые, как у египетского гуся, которые могли быть одновременно пригодны как для схватывания предметов, так и для процеживания воды; затем – в пластинки, похожие на пластинки домашней утки, и так далее, пока они стали столь же совершенными, как у широконоски, в форме аппарата, годного исключительно для процеживания. Начиная с этой стадии, на которой пластинки имеют две трети длины пластинок китового уса Balaenoptera rostrata, постепенные градации, наблюдаемые у ныне существующих Сetacea, приводят нас к огромным пластинам китового уса гренландского кита. У нас нет ни малейшего основания сомневаться в том, что каждый шаг в этом последовательном ряде мог быть полезным некоторым древним Cetacea при постепенной смене функций, медленно перестраивающихся в процессе их поступательного развития; эти шаги развития полезны настолько же, насколько полезны градации клювов у разных существующих ныне членов семейства утиных. Мы должны помнить, что каждый вид утиных подвержен жестокой борьбе за существование и что строение каждой части тела птицы должно быть хорошо адаптировано к условиям ее жизни.
Pleuronectidae, или камбалы, замечательны асимметричностью своего тела. Они лежат на одной стороне: большинство видов на левой, некоторые на правой; иногда встречаются взрослые особи с обратным против нормы положением тела. Нижняя сторона, на которой рыба лежит, на первый взгляд походит на брюшную сторону обыкновенной рыбы: она белого цвета, во многих отношениях развита менее верхней, с боковыми плавниками нередко уменьшенного размера. Но самую значительную особенность составляют глаза, так как оба находятся на верхней стороне головы. Однако в раннем возрасте они расположены друг против друга, и тогда все тело симметрично, с одинаково окрашенными обеими сторонами, вскоре глаз, соответствующий нижней стороне, начинает медленно перемещаться вокруг головы на ее верхнюю сторону, но не проходит прямо через череп, как думали раньше. Очевидно, пока нижний глаз совершает этот свой круговой переход, рыба, лежа в своем обычном положении на одном боку, не может им пользоваться. Находясь на нижней стороне, этот глаз, кроме того, подвергался бы трению о песчаную почву. Pleuronectidae по своей плоской форме и асимметричному строению превосходно адаптированы к их образу жизни; это доказывается тем, что многие виды, например морской язык, речная камбала и др., очень обычны. Наиболее важные преимущества, проистекающие из этого, по-видимому, состоят в защите от врагов и в благоприятных условиях питания на дне. Однако, по замечанию Шиёдте (Schiodte), разные члены этого семейства образуют «длинный ряд форм, представляющих градуальный переход от Hippoglossus pinguis, который не меняет сколько-нибудь заметно свой вид, в каком он оставляет яйцо, до морского языка, совершенно перевернутого на одну сторону».
М-р Майварт останавливается на этом случае и замечает, что внезапное самопроизвольное превращение в расположении глаз едва ли понятно, в чем я совершенно согласен с ним. К этому он прибавляет: «Если такое перемещение было градуальным, то далеко не ясно, в чем проявлялась польза для особи от такого перемещения глаза на очень малое расстояние с одной стороны головы на другую. Даже представляется, что такое неуловимое перемещение скорее должно быть вредным». Но ответ на свои возражения он мог бы найти в прекрасных наблюдениях Малма (Malm), опубликованных в 1867 г. Pleuronectidae в очень раннем возрасте еще симметричные, с глазами, расположенными на противоположных сторонах головы, не могут оставаться долго в вертикальном положении вследствие исключительной высоты их тела, очень малого размера их боковых плавников и отсутствия плавательного пузыря. Поэтому, быстро утомляясь, они опускаются на дно на один бок. Находясь в таком положении, они часто, как наблюдал Малм, поворачивают нижний глаз кверху, чтобы смотреть над собой, и делают это так энергично, что крепко прижимают глаз к верхнему краю глазницы. Легко можно видеть, что при этом передняя часть головы, находящаяся между двумя глазами, временно сокращается в ширину. В одном случае Малм видел, как молодая рыбка подымала и прижимала нижний глаз, причем угловое смещение достигало 70°.
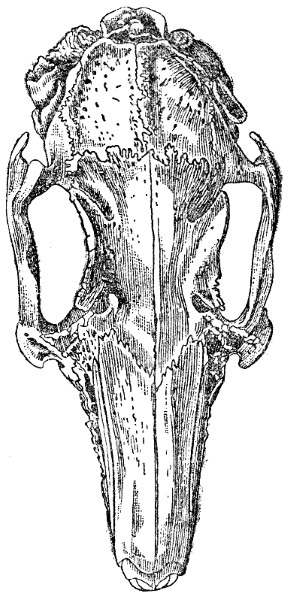
Череп полувислоухого кролика в естественную величину. Видно различие в направлении слухового прохода правой и левой стороны и причиняемое этим общее нарушение симметрии черепа. Свешивалось вперед левое ухо (на правой стороне рисунка).
Мы должны помнить, что череп на своей ранней стадии является хрящевым и гибким, отчего легко поддается воздействию мышц. Известно также, что у высших животных, даже после периода их ранней молодости, череп претерпевает изменения в своей форме, если кожа или мышцы постоянно сокращены под влиянием болезни или какой-либо другой случайности. У длинноухих кроликов, если одно ухо свешивается вперед и вниз, его тяжесть оттягивает вперед все кости черепа той же стороны, что я показал на рисунке. Малм утверждает, что только что вылупившиеся мальки окуней, лососей и разных других симметричных рыб имеют привычку время от времени отдыхать, лежа на дне на одном боку, и он наблюдал, что они часто поворачивают при этом свой нижний глаз кверху, вместе с тем череп становится несколько искривленным. Однако эти рыбы скоро становятся способными держаться постоянно в вертикальном положении, и их временная привычка не вызывает никаких последствий. Pleuronectidae же, с другой стороны, с возрастом все более лежат на боку вследствие возрастающего сплющивания их тела, и этим определяется постоянное действие на форму головы и положение глаза. Судя по аналогии, наклонность к искривлению, без сомнения, усиливается по закону наследственности. Шиёдте думает, в противоположность некоторым другим натуралистам, что Pleuronectidae не бывают совершенно симметричны даже в зародышевом состоянии, и, если это верно, мы можем понять, почему в раннем возрасте рыбки одних видов обычно опускаются и лежат на левой стороне, а других – на правой. Малм в подтверждение вышеприведенного мнения добавляет, что взрослая Trachypterus arcticus, не принадлежащая к семейству Pleuronectidae, лежит на дне на левом боку, а плавает в косом положении: у этой рыбы обе стороны головы, как говорят, несколько несимметричны. Наш выдающийся авторитет по рыбам, д-р Гюнтер, заключает свое резюме статьи Малма замечанием, что «автор дает очень простое объяснение ненормального состояния».
Из этого можно видеть, что первые стадии перемещения глаза с одной стороны головы на другую, которое м-р Майварт считает вредным, могут быть приписаны привычке, несомненно полезной для особи и для вида: пытаться смотреть вверх обоими глазами, лежа на дне на одном боку. Мы можем и другое приписать унаследованным последствиям употребления: рот у разных плоскотелых рыб обращен к нижней стороне и челюсти крепче и сильнее на этой лишенной глаз стороне головы, чем на другой, с целью более легкого, как предполагает д-р Траукер (Traquair), добывания пищи на дне. С другой стороны, неупотребление объясняет недоразвитие всей нижней половины тела, с боковыми плавниками включительно, хотя Яррель (Yarrell) думает, «что уменьшение этих боковых плавников полезно для рыбы, так как для их действия на нижней стороне гораздо менее места, чем на верхней». Быть может, уменьшение числа зубов до 4–7 в верхних половинах челюстей камбалы сравнительно с 25–30 в нижних также может быть объяснено неупотреблением. Из того факта, что брюшная сторона большинства рыб и многих других животных лишена окраски, с полным основанием можно предположить, что отсутствие окраски на одной стороне у камбал, будет ли нижней правая или левая сторона, вызывается отсутствием света. Но нельзя думать, что своеобразная пятнистая окраска верхней стороны морского языка, столь напоминающая песчаное дно моря, а также недавно доказанная Пуше (Pouchet) способность некоторых видов менять свою окраску в зависимости от окружающего фона или присутствие костяных бугорков на верхней стороне камбалы обязаны влиянию света. Здесь, вероятно, действовал естественный отбор, так же как и в приспособлении общей формы тела этих рыб и многих других особенностей к их образу жизни. Мы должны помнить, на чем я настаивал ранее, что унаследованные результаты усиленного употребления органа, а может быть, и их неупотребления, усиливаются под влиянием естественного отбора. Этим путем сохраняются все спонтанные вариации, идущие в надлежащем направлении, а также те особи, которые в наибольшей степени унаследовали результаты усиленного и полезного употребления какого-нибудь органа. Но невозможно в каждом отдельном случае решить, что следует приписать результатам употребления и что – влиянию естественного отбора.
Я могу привести и другой пример такого органа, который, по-видимому. обязан своим происхождением исключительно употреблению или привычке. Конец хвоста некоторых американских обезьян превращен в удивительно совершенный орган хватания и служит в качестве пятой руки. Наблюдатель, до мелочей сходящийся во взглядах с м-ром Майвартом, говорит об этом органе следующее: «Невозможно думать, что в течение ряда веков первая едва уловимая наклонность к хватанию могла служить к сохранению жизни особей, обладающих ею, или благоприятствовала их возможности иметь и вырастить потомков». Но в этом предположении нет никакой надобности. По всей вероятности, в этом случае достаточно привычки, которая уже сама по себе заставляет предполагать, что с ней связана большая или меньшая польза. Брем (Brelim) видел, что детеныши африканских мартышек (Cercopithecus) цеплялись руками за нижнюю часть тела матери, охватив в то же самое время ее хвост своими маленькими хвостиками. Проф. Хензло (Henslow) держал в неволе несколько полевых мышей (Mus messorius), у которых нет настоящего цепкого хвоста; но он часто видел, что они обвивали своими хвостами веточки поставленного в их клетке куста и таким образом помогали себе лазить. Аналогичное наблюдение было сообщено мне д-ром Гюнтером, который видел, как мышь подвешивалась таким образом. Если бы полевая мышь была более древесной по своему образу жизни, то, быть может, ее хвост сделался бы цепким по строению, что мы наблюдаем у некоторых членов того же самого отряда. Трудно сказать, почему Cercopithecus, при его привычках в раннем возрасте, этого не приобрел. Однако возможно, что длинный хвост этой мартышки больше служит ей в качестве органа равновесия при ее удивительных прыжках, нежели в качестве органа хватания.
Млечные железы свойственны всему классу млекопитающих и необходимы для их существования, поэтому они должны были развиться в очень отдаленный период, и мы не знаем ничего положительного о способе их развития. М-р Майварт задает вопрос: «Можно ли допустить, что детеныш какого-нибудь животного когда-нибудь спасся от гибели тем, что случайно высосал каплю едва питательной жидкости из случайно гипертрофированных кожных желез своей матери? И если так случилось однажды, то каковы шансы, что подобное изменение повторится?» Но этот пример изложен здесь неправильно. Большинство эволюционистов допускает, что млекопитающие произошли от сумчатых; а если так, то млечные железы сначала развились в мешке сумчатого. У одной рыбы (Hippocampus) вылупление из яиц и выращивание молоди в течение некоторого времени происходит в подобном же мешке, и американский натуралист м-р Локвуд (Lockwood) на основе собственных наблюдений над развитием мальков думает, что они питаются выделением кожных желез мешка. Что же касается отдаленных предков млекопитающих (еще до того, как они заслужили это название), то разве невозможно по крайней мере, чтобы их детеныши выкармливались подобным же образом. И в этом случае некоторые особи могли выделить жидкость до некоторой степени более питательную, так что она обладала свойствами молока; они могли в течение долгого промежутка времени воспитать большее число хорошо выкормленных потомков, нежели особи, выделявшие менее питательную жидкость; таким путем кожные железы, гомологичные млечным, могли усовершенствоваться, т. е. сделаться более производительными. Железы, находящиеся на некотором участке мешка, могли развиться более других, и это находится в полном согласии с широко распространенным принципом специализации; они могли затем образовать и грудь, сначала без соска, как у Ornithorhyncus, стоящего у основания ряда млекопитающих. Я не берусь решать, по какой причине железы на одном месте стали более специализированными, чем на других местах: при посредстве ли компенсации роста, результатов употребления или естественного отбора.
Развитие млечных желез не имело бы значения и не подпало бы под воздействие естественного отбора, если бы новорожденные не могли в то же самое время пользоваться их выделением. Как новорожденное млекопитающее научилось инстинктивно сосать грудь, понять не труднее, чем объяснить, как невылупившиеся цыплята научились разбивать яичную скорлупу, стуча по ней своим специально для этого адаптированным клювом, или как они научились клевать корм спустя несколько часов после того, как покинули скорлупу. В подобных случаях самое вероятное решение вопроса состоит, по-видимому, в том, что та или другая привычка была приобретена сначала на практике в более позднем возрасте, а потом передана потомку в более раннем возрасте. Но новорожденный кенгуру, как говорят, не сосет, а только присасывается к соску матери, которая обладает способностью впрыскивать молоко в рот своего беспомощного, полусформированного детеныша. Относительно этого м-р Майварт замечает: «Если бы не существовало особого защитного приспособления, новорожденный детеныш наверное погиб бы от проникновения молока в дыхательное горло. Но такое приспособление существует. Гортань удлинена настолько, что она доходит до заднего конца носового прохода, что дает воздуху возможность свободно проникать в легкие, в то время как молоко беспрепятственно стекает по бокам этой удлиненной гортани и таким образом благополучно проходит в пищевод позади нее». Затем м-р Майварт спрашивает, каким образом естественный отбор уничтожил у взрослого кенгуру (и у большинства других млекопитающих, если допустить, что они произошли от сумчатых) «эту по крайней мере совершенно невинную и безвредную особенность строения»? В ответ на это можно высказать предположение, что голос, который, конечно, имеет большое значение для многих животных, едва ли мог достичь полной силы, пока гортань вдавалась в носовой проход; а проф. Флауэр (Flower) высказал мне то соображение, что такое строение служило бы большой помехой для животного, питающегося твердой пищей.
Теперь мы обратимся на короткое время к низшим подразделениям животного царства. Echinodermata (морские звезды, морские ежи и пр.) снабжены замечательными органами, называемыми педицелляриями, которые во вполне развитом виде представляют собою щипцы с тремя ветвями, т. е. три зазубренные ветви, прочно соединенные друг с другом и укрепленные на верхушке гибкого ствола, который приводится в движение мышцами. Эти щипцы могут крепко схватывать разные предметы, и Александр Агассиц (Alexander Agassiz) видел, как Echinus, морской еж, быстро переправлял частицы экскрементов от щипцов к щипцам вниз в определенном направлении по своему телу, чтобы его поверхность не была загрязнена. Но, вне всякого сомнения, помимо удаления разных частиц грязи эти образования несут и другие функции; одной из них является, по-видимому, защита.
Относительно этих органов м-р Майварт, как и во многих других предыдущих случаях, спрашивает: «Какого рода польза могла быть от первого рудиментарного зачатка этих органов и каким образом эти ничтожные зачатки могли сохранить жизнь какому-нибудь одному морскому ежу?», и он добавляет: «Даже внезапное развитие хватательного движения не принесло бы пользы без вполне подвижного стебля, точно так же, как последний не имел бы значения без хватательных щипцов; однако никакие мелкие и притом неопределенные вариации не могли бы одновременно привести к образованию такого рода сложных координаций в строении; отрицать это – значит по меньшей мере защищать совершеннейший парадокс». Однако сколь парадоксально ни казалось бы это м-ру Майварту, щипцы с тремя ветвями, неподвижно прикрепленные к своему основанию, но способные к хватательному движению, несомненно существуют у некоторых морских звезд; и это вполне понятно, если они хотя бы частично служат орудием защиты. М-р Агассиц, любезности которого я обязан многими сведениями по данному вопросу, сообщил мне, что у некоторых морских звезд одна из трех ветвей щипцов редуцирована до уровня подставки для двух других, а существуют и такие роды, у которых третья ветвь вовсе утрачена. У Echinoneus, по описанию г-на Перриера (Perier), панцирь образует двух родов педицеллярии: одни, похожие на педицеллярии Echinus, другие – на педицеллярии Spatangus; такие случаи всегда интересны, так как они представляют средство для объяснения кажущихся внезапными переходов путем утраты одного из двух состояний органа.
Что касается промежуточных шагов, которыми шло развитие этих любопытных органов, то м-р Агассиц, на основании своих собственных исследований и исследований Мюллера, пришел к заключению, что как у морских звезд, так и у морских ежей педицеллярии несомненно должны рассматриваться как модифицированные иглы. В этом можно убедиться как из хода их развития у отдельной особи, так и из длинной совершенной серии градаций у разных видов и родов от простых бугорков к обычной игле и от нее к вполне развитой трехветвистой педицеллярии. Градация распространяется даже на способ, которым обычные иглы и педицеллярии с поддерживающими их известковыми пластинками причленяются к панцирю. У некоторых родов морских звезд мы находим «именно такие сочетания черт строения, которые показывают, что педицеллярии представляют собой просто модифицированные ветвичные иглы». Так, мы встречаем неподвижные иглы с тремя равноудаленными зазубренными подвижными ветвями, разделенными равными промежутками и сочлененными у своего основания, а выше, на той же самой игле, сидят три другие подвижные веточки. Если последние отходят от верхушки иглы, то они действительно образуют примитивную трехветвистую педицеллярию, которую можно видеть на одной и той же игле одновременно с тремя нижними веточками. В этом случае идентичность ветвей педицеллярии и подвижных ветвей иглы не подлежит сомнению. Обычно полагают, что простые иглы служат для защиты; а если так, то нет основания сомневаться, что и те иглы, которые снабжены зазубренными и подвижными ветвями, служат для той же цели; но они сделаются еще более эффективными, как только, соединившись, начнут действовать как хватательный и защелкивающий аппарат. Таким образом, все градации, от простой неподвижной иглы до неподвижной педицеллярии, полезны животному.
У некоторых родов морских звезд эти органы, вместо того чтобы быть прикрепленными к неподвижному основанию, находятся на верхушке короткого, но глубоко мускулистого основания; в этом случае они, вероятно, выполняют, помимо защиты, еще какую-нибудь дополнительную функцию. У морских ежей можно проследить все шаги того, как неподвижно сидящая игла приобретает сочленение с панцирем и делается таким образом подвижной. Я жалею, что не могу привести здесь более полного обзора интересных наблюдений м-ра Агассица над развитием педицеллярии. Он говорит также, что всевозможные градации могут быть равным образом найдены между педицелляриями морских звезд и крючками офиур, другой группы иглокожих, а также между педицелляриями морских ежей и якорями голотурий, также принадлежащих к тому же большому классу.

