Вероятные следствия действия естественного отбора путем дивергенции признака и вымирания потомков одного общего предка
На основании только что кратко изложенных соображений мы можем допустить, что модифицированные потомки какого-нибудь вида будут иметь тем более успеха, чем многообразнее будет их строение, что позволит им захватить места, занятые другими существами. Теперь посмотрим, как действует этот принцип полезности, выведенный из дивергенции признака и связанный с естественным отбором и вымиранием.
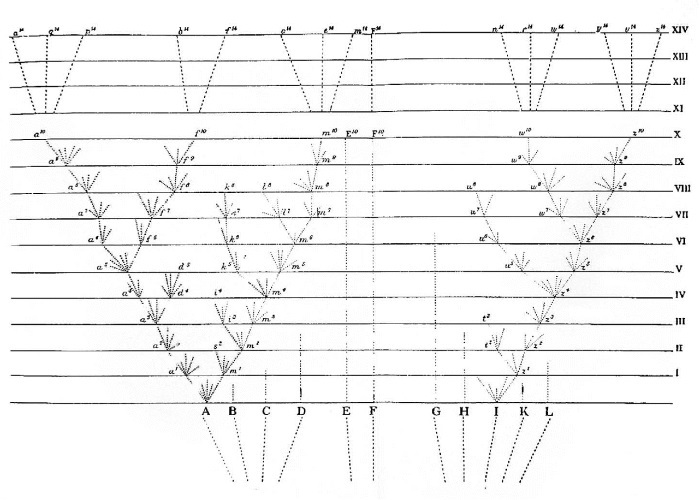
Прилагаемая диаграмма поможет нам уяснить себе этот довольно сложный вопрос. Пусть А до L будут виды обширного рода, обитающие в своей родной стране; предполагается, что эти виды сходны друг с другом не в одинаковой степени, как это бывает обычно в природе и как представлено на диаграмме буквами, расположенными на неравных расстояниях друг от друга. Я сказал: обширного рода, потому что, как мы видели во II главе, в среднем варьирует большее число видов в сравнительно больших родах, чем в меньших, и варьирующий вид значительно большего рода образует большее количество разновидностей. Мы видели также, что виды наиболее обычные и наиболее широко расселенные более вариабельны, чем виды редкие и с ограниченным распространением. Пусть (А) будет обычный, широко расселенный и варьирующий вид, принадлежащий к обширному роду в своей стране. Ветвящиеся и дивергирующиеся от (А) пунктирные линии различной длины представляют его варьирующих потомков. Вариации крайне слабые, но весьма многообразные; предполагается, что они возникают не все одновременно, но нередко через долгие промежутки времени и сохраняются неодинаково долго. Только те вариации, которые так или иначе полезны, сохраняются или подвергаются естественному отбору. Здесь обнаружит свое важное значение принцип полезности, выведенный из дивергенции признака, так как естественным отбором будут сохраняться и кумулироваться вариации наиболее различающиеся или наиболее дивергентные, представленные крайними пунктирными линиями. Когда пунктирная линия достигает одной из горизонтальных линий, где она обозначена строчной буквой с цифрой, предполагается, что кумулированный размер изменения достаточен для сформирования довольно хорошо выраженной разновидности, которая заслуживает упоминания в систематических сочинениях.
Промежутки между горизонтальными линиями на диаграмме могут соответствовать тысяче или еще большему числу поколений. Предполагается, что через тысячу поколений вид (А) произвел две прекрасно выраженные разновидности, а именно а1 и m1. Эти две разновидности будут в общем все еще подвержены действию тех же условий, которые сделали их родителей вариабельными, а наклонность к вариабельности сама по себе наследственна, следовательно, они также будут склонны варьировать и обычно почти в том же направлении, как и их родители. Сверх того, эти две разновидности, будучи слабо модифицированными формами, унаследуют те преимущества, которые сделали их родоначальную форму (A) более многочисленной, чем большинство других обитателей той же страны; они будут обладать и более общими преимуществами всего рода, к которому принадлежит произведший их вид; благодаря таким преимуществам этот род сделался обширным в его родной стране. А все эти обстоятельства благоприятствуют образованию новых разновидностей.
Если же две разновидности будут вариабельными, то снова наиболее дивергировавшие из них вариации будут обычно сохраняться в течение следующей тысячи поколений. Предполагается, что по истечении этого периода разновидность a1 на диаграмме образовала разновидность a2, которая в силу принципа дивергенции отличается от (A) более, чем разновидность а1. Разновидность m1, как предполагается, произвела две разновидности, m2 и s2, отличающиеся одна от другой и еще более от общего родоначальника (A). Этот процесс может продолжаться подобными ступенями неопределенно долгое время; одни разновидности через каждую тысячу поколений образуют только одну все более и более уклоняющуюся разновидность, другие произведут их две или три, и, наконец, третьи ничего не произведут. Таким образом, в общем будут происходить увеличение числа разновидностей или модифицированных потомков общего предка (A) и дивергенция признаков у них. На диаграмме процесс доведен до десятитысячного поколения, а в сокращенной и упрощенной форме до четырнадцатитысячного поколения.
Но я должен сделать здесь оговорку, что я не предполагаю, чтобы процесс этот когда-нибудь шел с такой правильностью, как показано на диаграмме, хотя и в ней допущены некоторые неправильности; не предполагаю я также, чтобы процесс этот был непрерывен; гораздо более вероятно, что каждая форма в течение долгих периодов остается неизменной и затем вновь подвергается модификации. Я не думаю также, чтобы неизменно сохранялись наиболее дивергентные разновидности: средняя форма может нередко уцелеть на долгое время, не произведя, а может быть, и произведя несколько модифицированных потомков, потому что естественный отбор всегда действует в соответствии с природой мест незанятых или отчасти занятых другими существами, а это зависит от бесконечно сложных соотношений. Но, как общее правило, чем разнообразнее будет строение потомков какого-нибудь вида, тем больше мест они способны захватить и тем более численно возрастет модифицированное потомство. На нашей диаграмме линии преемственности прерываются на определенных расстояниях, обозначенных строчными буквами с цифрой; эти буквы соответствуют последовательным формам, сделавшимся достаточно различающимися, чтобы быть отмеченными как разновидности. Но эти перерывы воображаемые, их можно было бы поместить где угодно, через промежутки, достаточно длинные для кумулирования значительного размера дивергентного изменения.
Так как все модифицированные потомки обычного и широко расселенного вида, принадлежащего к большому роду, будут склонны сохранять те преимущества, которые обеспечили жизненный успех их предку, то они будут увеличиваться в числе и дивергировать в своих признаках; это показано на диаграмме несколькими дивергировавшимися ветвями, исходящими от (А). Модифицированные потомки позднейших и более усовершенствованных родословных ветвей, вероятно, захватят места более ранних и менее усовершенствованных ветвей, а следовательно, и уничтожат их: это выражено на диаграмме тем, что некоторые из нижних ветвей не достигают верхних горизонтальных линий. В некоторых случаях, без сомнения, процесс модификации ограничится одной родословной линией, и число модифицированных потомков не возрастет, хотя размер дивергентной модификации, возможно, возрастет. Такой случай можно изобразить на диаграмме, уничтожив все линии, которые дивергируются от (А), за исключением линии, тянущейся от а1 до а10. Именно таким путем английская скаковая лошадь и английский пойнтер, по-видимому, медленно дивергировали в признаках от своих родоначальных форм, не образовав ни в том, ни в другом случае каких-либо ветвей или рас.
Через десять тысяч поколений вид (A), как мы предполагаем, образовал три формы а10, f10 и m10, которые вследствие дивергенции признаков в ряде последовательных поколений обнаруживают глубокие, хотя, быть может, и неравные различия как между собой, так и с их общим предком. Если мы предположим, что размер изменения в промежутке между двумя горизонтальными линиями на нашей диаграмме крайне мал, эти три формы могут представлять собой только три хорошо выраженные разновидности; но стоит допустить, что эти ступени в процессе модификации будут более многочисленны или большими по размерам, чтобы эти три формы превратились в сомнительные или даже во вполне определенные виды. Таким образом, диаграмма иллюстрирует ступени перерастания малых различий, свойственных разновидностям, в крупные различия, свойственные видам. Продолжаясь в течение еще большего числа поколений (что показано на диаграмме в сокращенной, упрощенной форме), этот процесс даст восемь видов, обозначенных буквами от а14 до m14, которые все происходят от (А). Таким путем, я полагаю, умножается число видов и образуются роды.
В сравнительно большом роде, по всей вероятности, будут изменяться не один, а несколько видов. На диаграмме я допускаю, что и другой вид (I) после десяти тысяч поколений произвел аналогичными ступенями либо две хорошо выраженные разновидности (w10 и z10), либо два вида соответственно предположению, какой размер изменения представлен расстоянием между горизонтальными линиями. Далее предполагается, что по истечении 14 тысяч поколений образовалось шесть новых видов, обозначенных буквами n14 до z14. В каждом роде те виды, которые уже наиболее между собой разнятся, будут обыкновенно склонны производить наибольшее количество модифицированных потомков, так как у этих последних будет наибольшая возможность завладеть новыми и наиболее различными местами в экономии природы; на основании этого я выбрал на диаграмме крайний вид (A) и почти крайний (I) представителями существенно различающихся форм, давших начало новым разновидностям и видам. Остальные девять видов (обозначенные прописными буквами) нашего первоначального рода могут продолжать в течение долгих, но неравных периодов давать почти неизменное потомство; это обозначено на диаграмме восходящими пунктирными линиями неравной длины.
Но в течение этого процесса модификации, представленного на диаграмме, будет играть важную роль и другой из установленных нами принципов – принцип вымирания. Так как во всякой предельно заселенной стране естественный отбор действует, только предоставляя отобранным формам некоторое преимущество над остальными в борьбе за жизнь, то у улучшенных потомков каждого вида будет проявляться тенденция на каждой стадии процесса заместить и истребить своих предшественников и исходного основателя рода. Не следует забывать, что конкуренция будет всего упорнее между формами, наиболее близкими по строению, конституции и образу жизни. Отсюда склонность к исчезновению будут иметь все промежуточные формы; а именно между ранними и более поздними состояниями, или, иначе, между менее совершенными и более совершенными состояниями одного и того же вида, а равно и сам родоначальный вид. То же, вероятно, обнаружится и во многих боковых родословных линиях, которые будут побеждены позднейшими и более усовершенствованными линиями. Если, однако, модифицированные потомки в отличие от исходного вида попадут в совершенно иную страну или быстро адаптируются к совершенно новой стации, где потомок и основатель рода не будут конкурировать друг с другом, то оба могут сохраниться.
Если предположить далее, что наша диаграмма представляет значительный размер модификации, значит, вид (A) и более всего ранние разновидности вымерли и заменены восемью новыми видами (а14 до m14), а вид (I) заменен шестью новыми видами (n14 до z14).
Но мы можем пойти еще далее. Первоначальные виды нашего рода, как мы уже условились, представляют весьма различные степени сходства друг с другом; оно так и бывает на деле в природе; вид (А) более близок к В, С и D, чем к другим видам, а вид (I) более близок к G, H, К, L, чем к другим. Эти два вида (А) и (I) по условию относятся к очень обыкновенным и широко расселенным видам, так что первоначально они должны были иметь какое-то преимущество над большинством других видов того же рода. В четырнадцатитысячном поколении этих двух видов 14 модифицированных потомков, вероятно, унаследовали часть этих преимуществ; они также модифицировались и улучшались в различных направлениях на каждой стадии рассматриваемого процесса, так что успели сделаться адаптированными ко многим местам в экономии природы своей страны. Весьма вероятно поэтому, что они заняли места не только своих родоначальников (А) и (I), но также и некоторых из первоначальных видов, наиболее близких к этим родоначальпым формам, и в конце концов истребили как тех, так и других. Таким образом, только небольшое число первоначальных видов доведет свое потомство до четырнадцатитысячного поколения. Предположим, что лишь один (F) из двух видов (Е и F), наименее близких к остальным девяти первоначальным видам, довел свое потомство до этой последней стадии рассматриваемого процесса.
Число новых видов, происшедших от первоначальных 11, будет на нашей диаграмме равно 15. Благодаря склонности естественного отбора к дивергенции, предельный размер различий в признаках между видами а14 и z14 будет значительно превышать различие между самыми крайними из первоначальных 11 видов. Сверх того, взаимная связь между новыми видами будет совсем иного рода. Из восьми потомков (А) три, обозначенные буквами a14, q14, p14, будут в очень близком родстве между собой, так как представляют ветви, недавно дивергировавшие от а10, b14 и f14; дивергировавшие в сравнительно ранний период от a5 будут в некоторой степени отличны от трех первых видов; и, наконец, о14, е14 и m14 будут в близком родстве между собой, но так как они дивергировали с остальными в самом начале процесса модификации, то будут глубоко отличаться от остальных пяти видов и образуют подрод или самостоятельный род.
Шесть потомков (I) образуют два подрода или рода. Но так как первоначальный вид (I) сильно отличался от (А), находясь почти на противоположном от него конце первоначального рода, то и шесть потомков (I) будут, в силу уже одной наследственности, резко отличаться от восьми потомков (A); а сверх того, эти две группы, как мы предполагали, продолжали дивергировать в различных направлениях. Промежуточные виды (и это особенно важное соображение), связавшие первоначальные виды (А) и (I), все, за исключением (F), вымерли, не оставив потомков. Отсюда шесть новых видов, происшедших от (I), и восемь, происшедших от (A), придется отнести к двум очень различным родам или даже к отдельным подсемействам.
Таким образом, по моему мнению, из двух или более видов одного рода образовались два или больше родов в силу общности происхождения, сопровождаемого модификацией. А эти два или более родоначальных вида, можно предположить, произошли от одного вида, принадлежащего к более древнему роду. На диаграмме это указано пунктирными линиями (под прописными буквами), образующими пучки ветвей, сходящихся в нисходящем направлении к одной точке; эта точка представляет собою тот вид, который был предполагаемым родоначальником некоторых из наших новых подродов и родов.
Следует на минуту остановить внимание на своеобразии нового вида F14, который, как мы предположили, не подвергся значительной дивергенции признаков, но сохранил форму (F) совсем без изменения или изменился лишь в слабой степени. В таком случае его родственная связь с остальными новыми видами будет очень любопытной и какой-то окольной. Происходя от формы, стоящей между родоначальными видами (А) и (I), о которых предполагается, что они вымерли и нам неизвестны, он будет до известной степени промежуточным по своим признакам между обеими группами, происшедшими от этих двух видов. Но так как эти две группы продолжали дивергировать в признаках от своих родоначальных форм, то новый вид (F14) будет являться непосредственно промежуточным не между существующими видами, а, скорее, между типами обеих групп; каждый натуралист, без сомнения, припомнит такие случаи.
На нашей диаграмме, как мы предположили, каждая горизонтальная линия соответствует тысяче поколений, но она может соответствовать миллиону и более поколений; может она также представлять разрез последовательных слоев земной коры, включающих остатки ископаемых. В главе о геологии мы вернемся к этому вопросу и, я полагаю, убедимся. что эта диаграмма бросает свет на родство вымерших форм, которые, хотя обычно относятся к тем же отрядам, семействам и родам, что и ныне живущие, тем не менее по своим признакам занимают нередко промежуточное положение между современными группами; и мы легко можем понять этот факт, так как вымершие виды жили в различно отдаленные эпохи, когда ветви нашей родословной еще не успели так дивергировать, как теперь.
Я не вижу основания для того, чтобы ограничивать процесс модификации, здесь поясненный, образованием одних только родов. Если мы предположим, что на нашей диаграмме каждая последующая группа расходящихся пунктирных линий представляет очень значительную величину изменения, то формы, обозначенные буквами а14 до p14; а равно и b14, и f14, и о14 до m14, образуют три четко различающихся рода. Мы получим также два очень различных рода, происходящих от (I) и еще больше отличающихся от потомков (А). Эти две группы родов образуют уже два различных семейства или отряда, смотря по тому, какова будет допущенная нами на диаграмме величина дивергентной модификации. А эти два новых семейства или отряда произошли от двух видов первоначального рода, которые в свою очередь являются, как мы предположили, потомками еще более древней, неизвестной нам формы.
Мы видели, что в каждой стране виды, принадлежащие к сравнительно большим родам, чаще образуют разновидности, или зарождающиеся виды. Этого и следовало ожидать: так как естественный отбор действует в силу преимущества, которое имеет какая-нибудь форма над другими в борьбе за существование, то он и будет главным образом действовать на те, которые уже обладают каким-либо преимуществом, а обширность какой-нибудь группы доказывает, что ее виды унаследовали от общего предка какое-то общее всем им преимущество. Таким образом, борьба за образование новых модифицированных потомков будет происходить главным образом между большими группами, стремящимися увеличить свою численность. Одна большая группа будет медленно одолевать другую большую группу, сокращая ее численность и тем снижая вероятность ее дальнейшего изменения и совершенствования. В пределах одной и той же большой группы позднее образовавшиеся и более совершенные подгруппы, из числа ответвляющихся и захватывающих новые места в экономии природы, будут постоянно склонны замещать и истреблять более старые и менее совершенные подгруппы. Малые и расчлененные группы и подгруппы, наконец, совершенно исчезнут. Заглядывая в будущее, мы можем предсказать, что группы органических существ, теперь обширные и доминирующие и в то же время наименее расчлененные, т. е. наименее пострадавшие от вымирания, будут еще долго разрастаться. Но за какими группами в конечном счете останется превосходство, никто не может предсказать, потому что, как мы знаем, многие группы, ранее наиболее экстенсивно развитые, теперь уже вымерли. Заглядывая в еще более отдаленное будущее, мы можем предсказать, что благодаря продолжительному и постоянному разрастанию больших групп множество более мелких групп будет окончательно уничтожено и не оставит модифицированных потомков, вследствие чего из видов, живущих в какой-нибудь определенный период, только очень немногие сохранят потомство в отдаленном будущем. Мне придется вернуться к этому вопросу в главе о классификации, но я все же добавлю, что, согласно с этим воззрением, до настоящего времени сохранилось потомство очень незначительного числа древнейших видов, всё потомство одного вида образует класс; мы можем отсюда понять, почему число классов так ограничено в каждом большом отделе животного и растительного царства. Хотя очень немногие из древнейших видов оставили по себе модифицированных потомков, тем не менее в отдаленные геологические периоды земля могла быть почти так же густо населена, как и теперь, видами многочисленных родов, семейств, отрядов и классов.

