Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 6 Золотой век
Дальше: Глава 8 Когда раскрылся железный занавес
Глава 7
Ковровые камни
В первую очередь следы жизнедеятельности древних бактерий, даже целых сообществ, сохранились в виде строматолитов (от греч. στρωμα – подстилка, ковер и λίθος – камень) – тонкослоистых, обычно известковых, куполовидных построек (рис. 7.1). Миллиметровая, реже сантиметровая, слоистость строматолитов проявляется в изменении соотношения микрокристаллических сростков разной размерности, концентрации глинистых частиц или органического вещества и содержания некоторых элементов, но всегда любой из этих показателей многократно, закономерно и ритмично меняется.
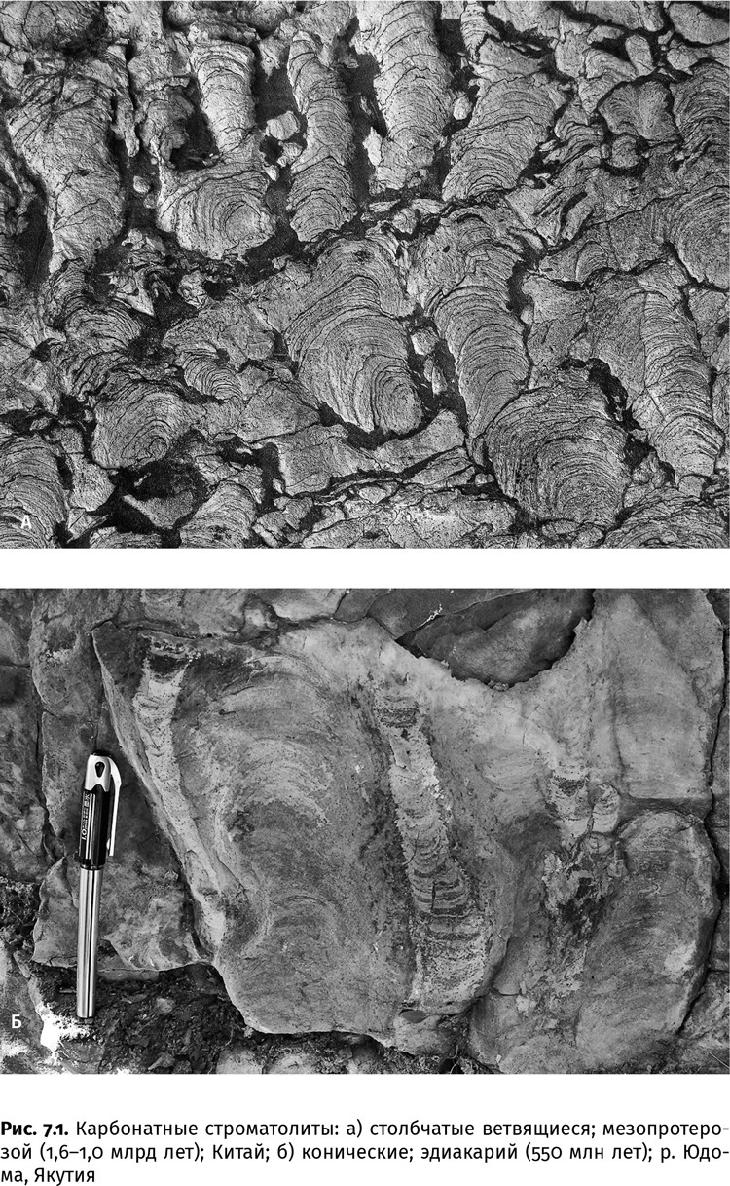
Нередко в качестве примера строматолитов приводят современные известковые столбики, растущие в гиперсоленой мелководной бухте Хамелин залива Шарк на западе Австралии. В развитии последних, правда, существенную роль играют одноклеточные эукариоты – диатомовые и зеленые водоросли. Поэтому и структура у них другая – не тонкослоистая, состоящая из миллиметровых разностей карбоната кальция, а рыхлая, грубая. К докембрийским строматолитам ближе те, что образуются сугубо бактериальными сообществами в лагунах Те-Куронг в Южной Австралии или Вермелья в Бразилии. Впрочем, и они не являются прямым аналогом, поскольку выживают в весьма особенных условиях: при резких изменениях солевого и температурного режима. Да и форма у них довольно простенькая: низкие, почти плоские купола. Не сравнить с тем огромным разнообразием строматолитов, что существовали в протерозойском эоне – кустистые и столбчатые, правильные конические (до 3 м высотой) и подушечные, словно гигантские площади (несколько квадратных километров), вымощенные булыжником. Внешне похожие на них бактериальные постройки сейчас существуют на глубинах в десятки метров в щелочных озерах на востоке Турции (Ван), юго-западе Канады (Павилион) и в Индонезии (кратер вулкана Сатонда). Но отчетливых слойков эти микробные сообщества не образуют, поэтому называются микробиалитами. Вряд ли стоит распространять и эту модель, к тому же реализованную при очень низком содержании кальция, на сложные древние строматолиты и, как уже говорилось, предполагать существование в архейском и протерозойском эонах щелочного океана, тем более болотно-озерных ландшафтов, хотя такие допущения делаются. Примеры палеопротерозойской группы Петей (1,88 млрд лет) на северо-западе Канадского щита или мезопротерозойской буровой свиты (1 млрд лет) на северо-западе Сибирской платформы (река Нижняя Тунгуска) показывают, что бактериальные строматолиты были распространены в очень разных морских обстановках: «булыжные мостовые» из коробчатых построек формировались на бурном, но хорошо освещенном мелководье; столбчатые и кустистые образовывали рифы несколько глубже; а огромные, многометровой высоты конусы развивались даже на внешней сублиторали, на пределе фотической (освещенной) зоны и ниже базиса штормовой эрозии. Глубоководные формы, стремясь к свету, нарастали на самом верхнем кончике, образуя столбики и конусы.
Строматолиты являются древнейшими видимыми невооруженным глазом свидетельствами жизни: небольшие конусы, в несколько сантиметров высотой, обнаружены в архейских породах Каапвааля и Пилбары (3,5–3,4 млрд лет). Конечно, похожие структуры могли отлагаться и неорганическим путем: ламиниты – на литорали, радиальные ботриоиды морского цемента – на сублиторали. И опыты показывают, что нечто очень похожее на строматолит получается без участия живых существ. Но есть у «живых» строматолитов ряд признаков, которые вряд ли появились за счет лишь физических и химических процессов. Биогенный строматолит чутко реагирует на смену обстановки: меняет форму в зависимости от того, откладываются его слойки в затишье или в шторм; ветвится и даже прорастает сквозь тонкий слой осадка, накрывший всю постройку после урагана. Бактерии проберутся среди песчинок и начнут свою работу заново. Неживая природа на такое не способна. Кроме того, в биогенных строматолитах есть слойки с микросгустковой текстурой – вероятно, микрит, осевший на бактериальную слизь и приклеившийся к ней. В ламинитах и морских цементах подобных неправильностей не обнаружено. А главное, в строматолитовых керогенах сохранились заметные следы органического фракционирования изотопов углерода (до –30‰), азот и сера.
Выделять слизь, точнее внеклеточное полимерное вещество (в основном полисахариды), микробам приходилось, чтобы хоть немного обезопасить себя от ультрафиолетового излучения. Молекулы такого вещества [карбоксильная (–COOН), гидроксильная (–OH), фосфатная (–РО) и аминовая (–NH2) группы], а также клеточная оболочка, теряя в нейтральной или слегка щелочной морской среде протоны, становились отрицательно заряженными. Поэтому поверхность бактериальной колонии легко улавливала различные катионы, в том числе Са2+, если среда, конечно, была им достаточно насыщена. Кроме того, цианобактериальные колонии в процессе кислородного фотосинтеза выделяли гидроксильные ионы, которые реагировали с растворенным в воде бикарбонатом (HCO3–), образуя анионы карбоната (CO32 –):
HCO3– + OH– ↔ CO32- + H2O.
Анионы CO32– в свою очередь притягивали катионы Са2+, и из раствора выпадали мельчайшие (2–8 мкм в диаметре) микросгустки аморфного кальцита или арагонита.
В основном строматолиты строили нитчатые цианобактерии (менее 0,5 мкм в диаметре и несколько десятков мкм длиной), образующие своего рода «микролуга» – однослойные биопленки и многослойные биоматы. Биоматы состояли из чередующихся вертикально стоящих и горизонтально уложенных бактериальных чехлов, а также слойков чистого карбоната. Слойки с хорошо различимыми бактериальными чехлами формировались в весенне-летнее время при хорошей освещенности и при поступлении больших объемов биогенных элементов; благодаря обильным внеклеточным органическим полимерам они улавливали тонкие минеральные частицы или связывали катионы металлов. Карбонатные отлагались в осенне-зимнее время, когда цианобактериальные макромолекулы деградировали, использовались в пищу другими микробами, а выделявшиеся при распаде органических веществ анионы (особенно карбонат и бикарбонат) реагировали с ионами металлов. Именно тогда из раствора выпадали микросгустки аморфных карбонатных минералов, которые позже кристаллизовались и срастались в минеральные слойки.
При поступлении в окружающую среду ионов железа могли получиться «железные» (сидеритовые) строматолиты, марганца – родохрозитовые. (Все это руды названных металлов.) Сама слизь улавливала различные по составу частицы. При бескислородном фотосинтезе, который осуществляют серные пурпурные и зеленые бактерии, в осадок могли выпадать сульфаты: гипс (CaSO4 × 2H2O), барит (ВaSO4) или целестин (SrSO4). (Кстати, гипс – важное строительное вяжущее вещество, а два других минерала – руды бария и стронция.)
Если строматолиты пропитались растворенным кремнеземом, поступавшим из близлежащего термального источника (как в современной Исландии), то в них нередко, словно в стеклянных препаратах, сохранялись и сами бактерии, создавшие эти странные конструкции. Не исключено, что кремневые строматолитовые корки могли формироваться бактериями «умышленно», особенно в архейских условиях: кремнезем прекрасно экранирует ультрафиолетовые лучи ближнего спектра (150-микронный слой этого минерала обеспечивает полную защиту; без него 70 % клеток гибнет в течение 24 часов).
Преимущественно коническая и столбчатая форма строматолитов предопределялась несколькими факторами. В первую очередь фототаксисом – стремлением к свету бактериальных сообществ, но не только. Поскольку цианобактериальная часть сообщества выделяла кислород, пузырьки газа скапливались на кончике строматолита, куда устремлялись оксифильные бактерии, а за ними другие – потребители органики. (Такие ископаемые пузырьки начинают встречаться в строматолитах возрастом 2,7 млрд лет – на исходе архейского эона.) А правильность сложных построек (выдержанное расстояние между отдельными конусами) объясняется тем, что бактериальное сообщество дробилось на отдельные группы, борющиеся друг с другом за ресурсы.
Строматолитовая форма существования широко распространилась (рис. 4.1л), поскольку жить под защитой минеральных слойков оказалось очень выгодно: биопленки и биоматы быстро высыхали и повреждались ультрафиолетовым излучением, а со временем стали выедаться одноклеточными эукариотами. Под минеральными слойками сохранялась влага, для ультрафиолета они были непроницаемы, а для того, чтобы их разрушить, нужны были железные зубы, как у современных моллюсков – хитонов. Кроме того, в инертные минералы можно было упрятать ионы тяжелых металлов и мышьяка, растворенные в бескислородном океане в повышенных концентрациях и потому представлявшие опасность для жизни.
В архейском океане в возведении карбонатных построек могли участвовать протео- и ацидобактерии (известные в современных микробиалитах), но, поскольку они не имели таких прочных оболочек, как у цианобактерий, шансов сохраниться у них не оставалось. Эти члены сообщества прокариот могли обеспечить и другие пути образования карбонатов, например через последовательность реакций, начинавшуюся с разложения диамида угольной кислоты [(NH2)2CO]:
(NH2)2CO + 3H2O → 2NH4– + HCO3– + OH–;2HCO3–+ Ca2+ → CaCO3 + H2O + CO2.
«Пузырьки кислорода», впервые застрявшие в строматолитах около 2,7 млрд лет назад, вероятно, и являются древнейшими свидетельствами существования цианобактерий – основной группы фототрофов (как в виде отдельных организмов, так и в форме пластид многих водорослей и высших растений). К этому же рубежу приурочены заметные изменения в изотопной подписи углерода (рис. 4.1е), повышенное содержание органического углерода в морских отложениях (такие объемы вряд ли могли образоваться за счет одного бескислородного фотосинтеза) и находки микрофоссилий, похожих на цианобактерии.
Морфология протерозойских бактериальных чехлов, скажем из сухотунгусской свиты Сибирской платформы, указывает на их принадлежность цианобактериям, внешне похожим на современные калотрикс (Calothrix) и формидиум (Phormidium): именно эти мелководные фототрофы нуждаются в толстых оболочках, предохраняющих от ультрафиолетового излучения. На присутствие цианобактерий указывают и состав органического вещества, иногда сохраняющегося в чехлах, и соотношение стабильных изотопов углерода. Способны были поучаствовать в строматолитостроении и коккоидные бактерии, образующие обильные колонии пленочной и грибовидной формы (напоминающие Entophysalis). В некоторых строматолитах насчитывается до десятка видов разных цианобактерий.
Цианобактерии жили не только под защитой строматолитовых корок: они освоились даже на периодически пересыхающем мелководье и играли там весьма заметную роль. Заметную благодаря золотистому пигменту (лат. pygmentum – краска) сцитонемину, предохранявшему клетки как цианобактерий, так и водорослей от ультрафиолетового излучения. Конечно, в клетках должны были находиться и пигменты, связанные с фотосинтезом, подобные хлорофиллам, поскольку изначально и они служили для рассеивания излучения. (Механизм рассеивания затем и использовался в фотосинтезе.) От гибели бактерии также защищались с помощью акинет – толстостенных удлиненных спор с обильными запасами питательных веществ: акинеты могли выдержать и временную засуху, и чрезмерное осолонение, и сильное опреснение.
Прослеживание молекулярной истории этих прокариот также показывает, что оксифототрофная ветвь отделилась от нефотосинтезирующей и анаэробной линии цианобактерий 2,6–2,5 млрд лет назад. Последние могли существовать и раньше и даже отладить фотосистему II – один из важнейших компонентов всей системы фотосинтеза, необходимый для расщепления молекул воды. В бескислородном архейском океане эта фотосистема была задействована для забора электрона у Н2S, чтобы окислить S2– до S0. С этим источником электронов, так же как с Fe2+, разные прокариоты не могли расстаться еще долго.
Назад: Глава 6 Золотой век
Дальше: Глава 8 Когда раскрылся железный занавес

