Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 25 Неожиданный конец ордовикской истории
Дальше: Глава 27 Конец первозданной пустыни
Глава 26
Во враждебном мире
По мере того как одни механизмы, влиявшие на темпы химического выветривания, себя исчерпывали, растения, эволюционируя, создавали новые. И хотя развитие жизни на суше шло плавно и поступательно, особенно важные растительные инновации сказывались на морских обитателях не всегда положительно и даже, скорее, наоборот. Об этом позднеордовикские события и свидетельствуют.
Переход из воды на сушу был шагом болезненным и нередко смертельным. Достаточно после небольшого шторма пройтись по пляжу, чтобы увидеть сотни водорослей, медуз, морских звезд и ежей, рыб, которым никогда уже не суждено вернуться в спасительную стихию. Освоение суши давалось с большим трудом и длилось не одну сотню миллионов лет. Ведь требовалось «придумать», как избежать потерь драгоценной влаги, а для животных – еще и овладеть существенно иным стилем передвижения. Взлететь – и то оказалось проще.
Неудивительно, что из 30 типов животных на сушу рискнули выползти и выйти представители только восьми. Кроме известных кольчатых червей, моллюсков (улитки и слизни), членистоногих и позвоночных это плоские и круглые черви, тихоходки и онихофоры (последние две группы являются потомками ксенузий). Все наземные растения произошли от одной-единственной группы зеленых водорослей, а первые грибы тоже появились в водной среде. Даже обосновавшись на суше, многие организмы стремились при случае вернуться в воду. Так появились морские травы и мангровые леса, водные насекомые, огромное разнообразие морских рептилий (из которых до нашего времени дожили только крокодилы и черепахи), ушастые и настоящие тюлени, а также киты. А земноводные по-настоящему так никогда и не отважились покинуть прибрежную полосу.
Большинство жителей суши по-прежнему развиваются в водной среде. Ведь и яйцо птицы или рептилии, и утроба высших млекопитающих не что иное, как бассейн, до краев наполненный жидкостью. Эмбрион человека – это сосуд с водой, который со временем пустеет: 77 % жидкости остается в новорожденном и 60 % – у взрослого, перевалившего на вторую половину жизненного пути. Лишившись четвертой части своей жидкости из-за отсутствия питьевой воды, человек умирает, а процессы, необратимо ведущие к смерти, начинаются в теле при потере 10 % влаги. Через почки в год протекает 1000 л жидкости; сердце прогоняет более 100 л ежедневно. В среднем за свою жизнь человек выпивает (и выделяет) 75 000 кг воды, а каждое поколение людей (при средней продолжительности жизни 70 лет) потребляет половину всего годового стока рек мира. О невежах римляне говорили: «Он не умеет ни читать, ни плавать». Ведь и великие человеческие цивилизации развивались только там, где в избытке была вода.
Так стоило ли при таких затратах и проблемах вообще вылезать на сушу?
Не очень хотелось, но пришлось. Причины, вынудившие животных и растения покинуть колыбель жизни, на первый взгляд кажутся загадочными. Действительно, зачем выбираться из жидкой среды, которая и поит, и кормит, и поддерживает в вертикальном положении, и надежно укрывает от отнюдь не полезного и разрушительного для нежных клеток ультрафиолетового излучения, и удачно решает проблему размножения – обеспечивает встречу с себе подобными или с их половыми клетками?
Впрочем, какими бы световоспринимающими пигментами ни обладали водоросли (зелеными, желтыми, пурпурными), они все равно нуждаются в свете Солнца – главном источнике энергии. Вода очень быстро поглощает излучение, и на глубинах свыше 180 м царит полный мрак. Потому все многообразие донных водорослей сосредоточено у кромки берега, а зеленые и бурые, зависимые от лучей средней части светового спектра, вынуждены ютиться в приливно-отливной зоне. Волей-неволей некоторым из них пришлось приспосабливаться к приливно-отливным циклам, пребывая дважды в сутки по нескольку часов на воздухе. В ордовикском периоде, когда толщу воды наполнили мириады планктонных организмов, застивших и без того скудный свет, а по таллому (тело водоросли) поползли зубастые моллюски, вгрызавшиеся в самую мякоть, у водорослей не оказалось иного выхода, кроме выхода на сушу. Преуспеть в этом могли именно самые мелководные – бурые и зеленые. Среди древних представителей этих групп и должны были появиться предки наземных растений.
В современных приливно-отливных сообществах распространены бурые водоросли. Их клетки покрыты прочной кутикулой (тонкая пленка из органических полимеров, предохраняющая таллом от высыхания, микроорганизмов и отчасти от выедателей), и сами они покрупнее и помассивнее прочих соплеменников. Потому нежные зеленые водоросли скрываются в часы осушения под талломами бурых, где сохраняется влага. И все-таки преуспели в завоевании суши зеленые. Возможно, сказался климатический фактор: бурые водоросли сосредоточены в полярных и умеренных широтах, а зеленые – в тропических. А у холодноводных организмов многие процессы замедлены; длительные зимние периоды они вообще пребывают в состоянии анабиоза – ни жизни, ни смерти. Если в течение нескольких десятилетий летние температуры остаются низкими, то все это время продолжается и спячка – в виде цист, спор, куколок и т. д. Одно поколение весьма неспешно замещает предыдущее, и число преобразований, необходимых для превращения одного вида в другой, накапливается долго. В тропиках деятельная жизнь продолжается круглый год, и смена поколений происходит в ускоренном темпе, усиливая изменчивость. Кроме того, в тропиках гораздо разнообразнее и растительноядные животные, и паразиты, от которых необходимо как-то спасаться, изобретая все новые и новые способы и средства защиты. Конечно, на эволюции видов сказываются все эти факторы, вместе взятые.
Так что зеленые водоросли получили над бурыми определенное преимущество, чем и воспользовались. Родство высших растений с зелеными водорослями трудно оспаривать. И те и другие получают и преобразуют солнечную энергию с помощью одного и того же набора зеленых пигментов (одинаковые разновидности хлорофиллов), и те и другие накапливают свои энергетические запасы в виде крахмала. Данные молекулярной биологии тоже показывают, что предками всех наземных растений стали именно зеленые водоросли, но какие именно, не вполне ясно до сих пор. Крупные многоклеточные ульвовые и харовые освоили пресноводные и земноводные условия и очень похожи на сосудистые растения, особенно харовые. Членистые талломы харовых с правильно расположенными листовидными ответвлениями очень напоминают стебли с листьями хвощей (одно время их считали близкими родственниками). Однако первые ископаемые харовые – а они, благодаря обызвествлению некоторых клеточных оболочек, сохраняются очень хорошо – известны лишь из девонских отложений, в то время как наземные растения, причем довольно разнообразные, встречаются уже в силурийских. К тому же у харовых нет правильного чередования полового и бесполого поколений, свойственного ульвовым. У последних сменяют друг друга многоклеточные пластинчатые гаметофит и спорофит, как у ранних сосудистых растений. (На гаметофите образуются гаметы – половые клетки с одинарным набором хромосом, т. е. гаплоидные; разнополые гаметы сливаются в зиготу, из которой развивается спорофит с двойным набором хромосом; спорофит производит бесполые споры, что сопровождается делением клеточных ядер, и новое поколение гаметофитов, прорастающее из таких спор, вновь оказывается гаплоидным.)
Несомненно, зеленые растения оказались на суше еще в ордовикском периоде (около 470 млн лет назад), о чем и свидетельствуют находки фрагментов жестких чехлов и многочисленных спор с плотными оболочками. Такие чехлы понадобились водорослям еще в эдиакарском периоде, чтобы противостоять выеданию. Они же помогли выйти на сушу, где без опоры не обойтись. Одновременно чехол предохранял растение от потери влаги. У позднеэдиакарских-раннекембрийских водорослей (550–530 млн лет) встречаются и другие необычные для водных обитателей органы: скрученные в тугую пружинку перфорированные ленты – кохлеатины (Cochleatina) до 70 мкм шириной и до 300 мкм в диаметре. Стоило такой пружинке высохнуть, и она должна была с треском раскрыться и разметать на несколько метров вокруг себя возможные споры, как это происходит у разных современных наземных растений и грибов. Значит, водоросли, несущие кохлеатины, если и не росли на суше, то возвышались над водой, и подобный способ воздушного расселения позднее мог использоваться настоящими обитателями суши. Споры, обернутые защитной, устойчивой к высыханию оболочкой, помогали внедриться в негостеприимное сухое пространство: они сохраняются в засушливое время, и с приходом влажного сезона растение возрождается к жизни. Не исключено, что растения осваивали сушу в два захода: сначала – относительно приспособленный к безводным условиям спорофит, а позднее – гаметофит с половыми клетками, нуждавшимися в водной среде для перемещения.
В конце силурийского периода (427–419 млн лет назад) на суше появились несомненные растения: риниофиты (Rhyniophyta; от шотландского местечка Райни и греч. φυτόν – растение), зостерофиллофиты (Zosterophyllophyta; от греч. ζωστήρ – пояс и φύλλον – лист), тримерофиты (Trimerophyta; от греч. τρί-μέρος – трехчастный), плауны (они единственные дожили до наших дней; рис. 24.1) и несколько других менее многочисленных групп. Пока у этих низкорослых первопоселенцев не было ни корней, ни настоящих листьев. Был лишь тонкий, правильно ветвящийся побег (ось) с округлыми спорангиями. Собственно, и фотосинтезирующие клетки находились в побеге. Нижняя часть побега (ризомы) тоже многократно ветвилась и стелилась вдоль поверхности почвы или даже в водной среде. От ризомов отходили нитевидные одноклеточные (или однорядные многоклеточные) выросты – ризоиды, которые служили для закрепления на субстрате и поглощения воды и питательных элементов. Настоящей сосудистой ткани еще не было: только удлиненные веретеновидные проводящие клетки – трахеиды, расположенные вдоль оси побега и подводящие воду ко всем органам. (Есть подобие таких образований – гидроидные клетки – и у мхов с печеночниками.) Различались древние растения по расположению, форме и строению спорангиев – органов, где развивались споры, причем спорофит и гаметофит у них, видимо, были соразмерными, хотя и не похожими друг на друга. У зостерофиллофитов на побеге возникли эмергенцы – мелкие игловидные выросты, а у тримерофитов и плаунов – микрофиллы (буквально – «мелкие листочки»), куда из побега вел тонкий сосудик, жилка. Основанием микрофиллы плотно прилегали к побегу. Так растения начали увеличивать площадь фотосинтезирующих органов. Появилось у них еще одно важное приобретение – устьица. Сквозь эту систему замыкающих клеток, расположенных на побегах, микрофиллах и спорангиях, углекислый газ и кислород проникали внутрь, под чехол, а вода испарялась. Устьица и водопроводящие клетки позволяли растениям окончательно освоиться на суше, поскольку испарение стало той силой, которое втягивало воду из почвы, где сидели ризомы, и тащило до самых замыкающих клеток.
Растения распространились по огромному континенту – Лавруссии, образовавшемуся после закрытия океана Япет и коллизии Лаврентии и Балтии. Лавруссия находилась в Северном полушарии, и на ее шельфе как раз накапливался древний красный песчаник. Южный континент Гондвану в конце силурийского периода облюбовали плауны, а самый северный материк – Ангарида (прежняя Сибирь и «пристыковавшиеся» к ней монгольские и казахстанские микроконтиненты) – из-за своего приполярного положения, видимо, еще не зазеленел. Обитали древние наземные растения в сырых, насыщенных влагой низинах и почти отмирали в засуху. В сухие сезоны современный плаун сворачивается, словно еж, припрятав внутри клубка зерна хлорофилла. Так он был способен «проспать» лет 15. Возможно, подобным образом выживали и растительные первопроходцы суши. Их побеги еще не стали надежной опорой, и растеньица поддерживали друг дружку, собираясь в плотные пучки, полупогруженные в воду.
В основном все силурийские и раннедевонские растения не превышали в высоту 0,1 м, лишь немногие дорастали до 2 м. В середине девонского периода «стартовала гонка за солнцем»: чтобы опередить соседей, не остаться в тени и не зачахнуть, растениям пришлось тянуться вверх. Но, чтобы подняться выше, нужно было обеспечить себе опору и овладеть гидравликой – наукой, изучающей законы движения и равновесия жидкостей. Согласно ее правилам, для водопровода лучше всего подходят цилиндрические трубы. Если у первых наземных растений роль проводящих сосудов играли отдельные веретеновидные трахеиды (1–4 мм длиной) с усиленными, но эластичными стенками, то к концу девонского периода у некоторых деревьев проводящие пучки уже состояли из трахеид на порядок длиннее (30–40 мм) и несколько шире. Еще сильнее увеличился диаметр трахеид у каменноугольных древовидных плаунов и семенных папоротников. А чем больше радиус трубы, тем выше ее проводимость и скорость текущего в ней потока: увеличение диаметра всего в два раза повышает проводимость трубки в 16 раз. Проводимость всей ткани зависит от числа отдельных трубок-сосудов. Однако труба большого диаметра должна выдерживать сильное давление, поэтому для укрепления ее стенок требуется пропитка, в качестве которой растения стали использовать лигнин (лат. lignum – древесина). Этот нерегулярный полимер формируется у наземных растений на основе довольно распространенного продукта обмена веществ – фенилпропанового радикала (С3-С6). В составе спорополленина тот же компонент защищает ткани от ультрафиолетового излучения наиболее опасного коротковолнового спектра. Оказалось, что это вещество пригодно и для укрепления (одеревенения) клеточных оболочек. Благодаря одеревенению и получилась водопроводящая ткань – ксилема. Попутно лигнин превратился в мощное препятствие для патогенных грибов и растительноядных животных, а также в накопитель углерода – 30 % всей современной растительной биомассы приходится на это вещество. Без укрепленных трубок-капилляров поднять самотеком воду на 10-метровую высоту растениям не удалось бы. Чтобы такая махина не падала от малейшего дуновения ветра, необходимо было «заякориться» в почве – пустить корни. Возраст древнейших ископаемых корней – 411 млн лет, известны они у раннедевонских плаунов. Лигнин появился позднее: раннедевонские растения, судя по их изотопной подписи углерода и вещественному анализу оболочек проводящих клеток, этот полимер для укрепления тканей еще не использовали.
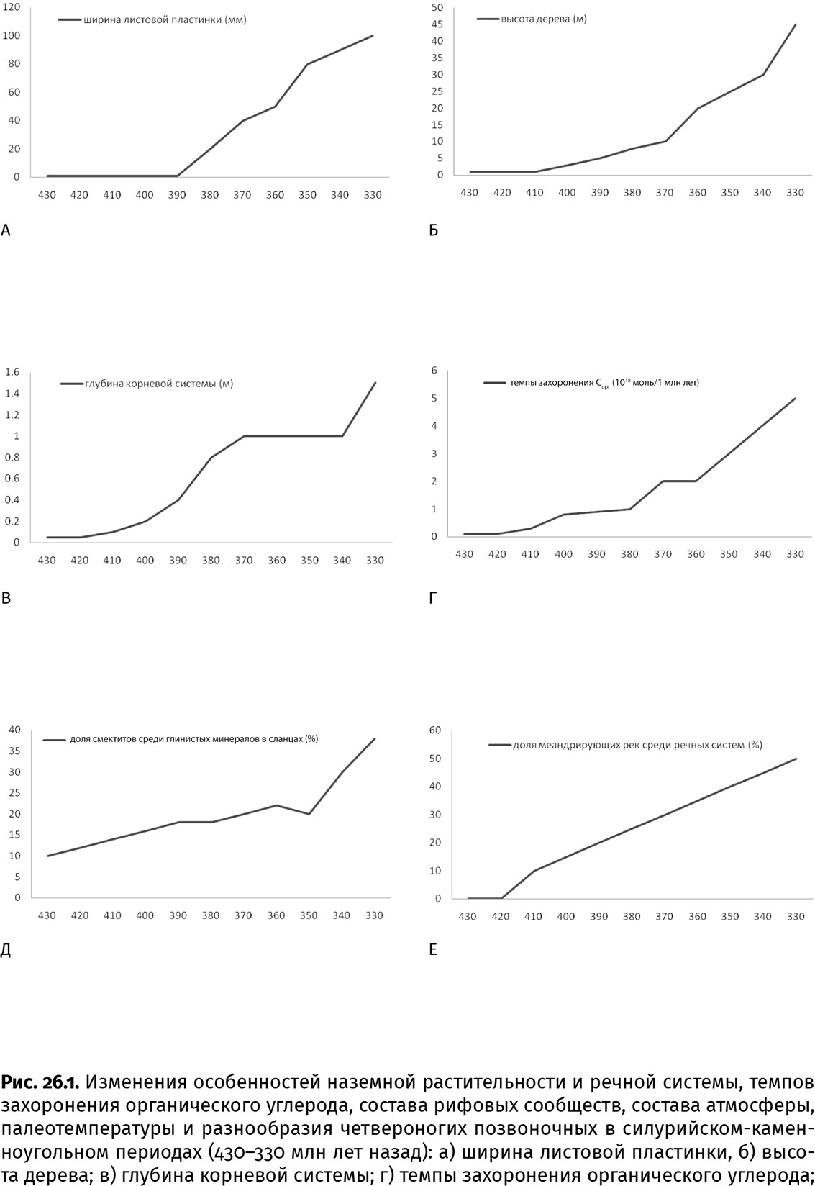
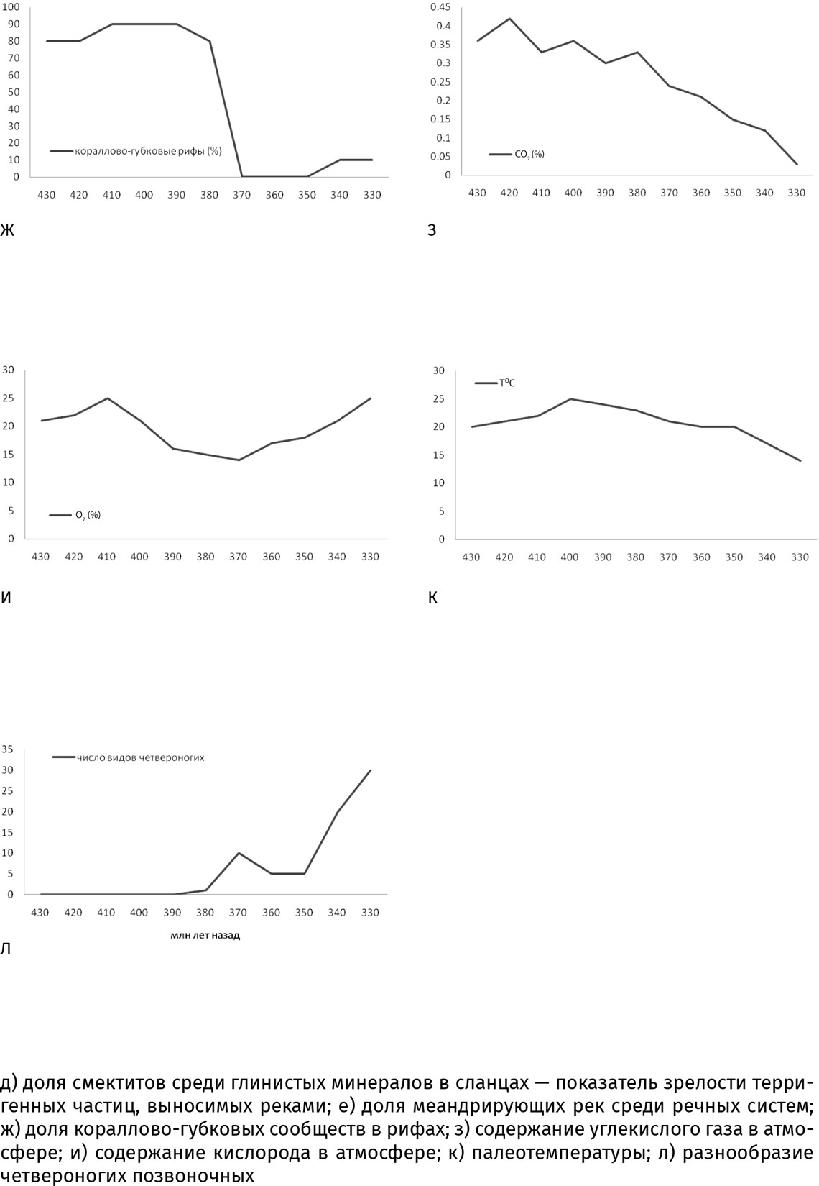
В течение последующих 50 млн лет своего существования наземные растения за счет всех усовершенствований увеличились в обхвате в 30 с лишним раз (до 3 м и более) и вытянулись по меньшей мере в 8 раз (до 8 м, возможно, и до 30 м). Так и возникли деревья. Растения древовидного облика – с глубокой корневой системой и широкими листьями – стали распространяться около 380 млн лет назад (рис. 26.1а – в). Настоящие листья, или макрофиллы, получились из пластинок, разросшихся между частыми ответвлениями побега. На дальнейшем увеличении площади листа сказалось снижение уровня углекислого газа: чтобы захватить достаточный объем ставших дефицитными молекул, понадобилось не только повысить плотность устьиц, но и расширить листовую пластину, чтобы их уместилось побольше.
Начиная со второго, пражского, века девонского периода (411–408 млн лет назад) растительный покров стремительно распространялся на все большее пространство: место низкорослых околоводных низинных «лугов» из голых риниофитов, зостерофиллофитов и тримерофитов сначала заняли травянистые и кустистые плауны и первые папоротники. Они все еще придерживались обводненных низин, но настоящие леса из древовидных плаунов, папоротников и некоторых других деревьев начали осваивать сухие возвышенности. (Все эти растения вели начало от тримерофитов, кроме плаунов, скорее всего произошедших от зостерофиллофитов.) Если в раннедевонскую эпоху растительность покрывала не более десятой части суши, то к концу периода уже занимала свыше двух третей ее площади. Распространение растительного покрова придало суше более темный цвет, что понизило альбедо планеты и смягчило климат в наступающую каменноугольную ледниковую эру.
В лесах, сложившихся к концу девонского периода (около 360 млн лет назад), существенную роль стали играть несколько новых групп, в первую очередь прогимноспермы (Progymnospermophyta), такие как археоптерис (Archaeopteris). Из-за сходства в строении перистых листьев – вай, расположенных правильной восходящей спиралью, – и спорангиев, сидевших на листовой пластине, подобные ископаемые растения долгое время считались папоротниками. Однако строение ствола с вторичной древесиной (дополнительные упрочняющие наслоения ксилемы) и усиленной проводящей тканью, состоявшей из мелких, но прочных трахеид, сближает их с голосеменными (Gymnospermaе), предками которых они могли быть. Плотная вторичная древесина позволяла наращивать ствол до 1,5 м в поперечнике и 25 м и более высотой, а пористые трахеиды помогали с максимальной пользой использовать капиллярные явления и поднимать воду на эту высоту; большая плотность этих клеток давала возможность прогимноспермам выживать в маловодных условиях, даже на солончаках. Еще они отличались от споровых современников развитой корневой системой – постоянно обновлявшейся, с обильной сетью тонких корневых волосков, способных проникнуть в любую пору, и уходившей на метровую глубину.
Надо отметить, что эффективную водопроводящую систему можно было организовать двумя основными способами. Во-первых, построить ее из относительно коротких, но обильных трахеид с многочисленными порами и сильно укрепленными (лигнифицированными) стенками. Такое сочетание высокой механической прочности и проводимости (благодаря многократному дублированию «каналов») позволяло выживать в маловодных условиях с сильными перепадами температур. Ксилема подобного типа появилась у прогимноспермов, что и позволило им распространиться на прежде неблагоприятные для растений территории, а также у кордаитов, глоссоптериевых (о них немного ниже) и хвойных (рис. 26.2). Поэтому хвойные по-прежнему прекрасно себя чувствуют в условиях «вечной мерзлоты», высокогорий и сухих средиземноморских ландшафтов.

Другой вариант ксилемы – большие проводящие клетки – обеспечивал более быстрый рост, но при условии постоянного бесперебойного подвода жидкости, что возможно только в безморозном климате, иначе закупорка трахеид или образование в них разреженностей приводили к гибели ткани. Функцию основной опоры при этом принимала на себя кора. Стволы с подобным строением известны у ряда голосеменных (тригонокарповые, цикадовые), семенных папоротников (таких, как лигиноптериевые – Lyginopteridales), а также в некоторой степени у древовидных плаунов, хвощей и папоротников (рис. 26.3). (Забегая вперед, нужно отметить, что наиболее совершенная проводящая ткань возникла у покрытосеменных: у них появились настоящие сосуды – открытые с обоих концов цилиндрические трубки, а опорную функцию стали выполнять специализированные сильно лигнифицированные непроводящие клетки; современная береза высотой 15 м, например, благодаря такому строению ствола «поднимает» по два десятка 10-литровых ведер воды на уровень шестого этажа ежедневно.)
Подробное освещение истории и разнообразия наземных растений потребовалось потому, что все приспособления (устьица, корни, сосудистые клетки, древесина, семя), которыми они обзавелись, чтобы выжить, позволили им стать новой геологической силой, способной преобразовывать планету не в меньшей степени, чем вулканические, а быть может, и тектонические процессы.



