Книга: Что значит быть собакой. И другие открытия в области нейробиологии животных
Назад: Глава 5 Зачатки способностей
Дальше: Глава 7 Буриданов осел
Глава 6
Рисование звуком
Моя давняя коллега по Университету Эмори, Лори Марино, знает о дельфинах все. В своей диссертации она сравнивала анатомию черепа китообразных и приматов и пришла к неожиданному выводу: зубатые киты, к которым относятся и дельфины, обладают одним из самых высоких коэффициентов энцефализации в животном мире.
Почти на всем протяжении нулевых Лори занималась изучением мозга дельфинов. Она подключилась к сети оповещений о выбрасывании морских млекопитающих на берег и, когда на атлантическом побережье погибал дельфин, всеми правдами и неправдами старалась раздобыть его мозг. Так, упорными усилиями ей удалось собрать небольшую коллекцию образцов, которые она начала исследовать с помощью МРТ. Именно ей, Лори Марино, мы во многом обязаны современными представлениями об анатомии мозга дельфинов.
Помимо анатомических исследований Лори прославилась сотрудничеством с Дианой Рейс, профессором психологии из Городского университета Нью-Йорка, — в ходе совместной работы они выясняли, узнаю́т ли дельфины себя в зеркале. Зеркальный тест еще в 1960-х разработал психолог Гордон Гэллап, исследовавший степень самосознания у шимпанзе. Для этого на лбу шимпанзе делали отметку мелом и наблюдали за реакцией подопытного, когда тот смотрел в зеркало. Касание отметки пальцем свидетельствовало о наличии самосознания. Шимпанзе у Гэллапа этот тест успешно проходили, как проходят его человеческие дети с полутора лет.
Лори и Рейс протестировали двух живших в неволе дельфинов и обнаружили, что при наличии метки на какой-либо части тела, в том числе по бокам головы, они проводили перед зеркалом больше времени, чем при отсутствии метки. Это открытие сильно повлияло на Лори. Если дельфины обладают самосознанием, может быть, у них и когнитивная сфера, как у человека? Какое тогда мы имеем право держать их в неволе?
Когда я обратился к Лори по поводу нейровизуализации мозга дельфинов, она сразу осознала огромный потенциал диффузионной МРТ как способа ответить на давние вопросы о психическом опыте дельфинов. Эти животные поражают невероятной широтой диапазона издаваемых звуков. Часть из них выполняет коммуникативную функцию, остальные используются для подводной эхолокации. Однако, несмотря на то что эхолокационную способность дельфинов изучали уже не первое десятилетие, по-прежнему оставалось неясным, как их мозг обрабатывает эти данные, создавая когнитивную карту окружающей морской среды.
Томас Нагель счел бы нашу затею заведомо бессмысленной, поскольку различия между дельфином и человеком слишком велики и мы никогда не поймем, каково это — жить в море или пользоваться эхолокацией. Я считал иначе. Если мы выясним, как работает мозг дельфина при эхолокации, то еще на шаг приблизимся к пониманию субъективных ощущений этих животных. Кроме того, зачаточная способность к эхолокации у человека имеется. А значит, мозг дельфина может что-то рассказать нам и о человеке.
— Замечательно! — обрадовалась Лори. — Никто еще не делал диффузионную МРТ мозга дельфина.
Разделив ее восторг, я перешел к более насущным вопросам.
— Может быть, вы подскажете, где раздобыть мозг дельфина?
— Конечно. У меня остались все образцы, которые я сканировала десять лет назад. Они в вашем полном распоряжении.
Все складывалось даже удачнее, чем я ожидал. Но меня смущал возраст образцов. Диффузионную МРТ на мертвом мозге проводить и без того довольно трудно, а уж тут… Кто знает, какой сигнал мы получим от препарата, более десяти лет пролежавшего в формалине.
Мои размышления о возможных трудностях прервала Лори:
— Вы же знаете, я ухожу, — сообщила она, понизив голос.
В Университете Эмори она постоянно переводилась с кафедры на кафедру, собственно, как и я. Когда твоя исследовательская работа не вписывается в четкие рамки, найти постоянное прибежище зачастую бывает сложно. Это не значит, что коллеги принимают тебя в штыки, но собеседника иногда не хватает.
У Лори все больше и больше времени отнимала зоозащитная деятельность, которой она занялась, когда узнала о ежегодном жестоком истреблении дельфинов в японской бухте Тайдзи. Лори включилась в борьбу за китов и дельфинов, а затем выступила в документальном фильме 2013 года под названием «Черный плавник». В итоге она оказалась на распутье: либо целиком и полностью посвятить себя защите дельфинов и других животных, либо продолжить преподавать основы нейронауки и пытаться выкроить время на исследования. Выбор был очевиден.
Только теперь я заметил стоящие в кабинете коробки. Она уезжала через неделю.
— И куда вы? — спросил я.
— В Юту.
В Юту? Странно. А что там?
— У меня большие планы. Я учреждаю новый фонд — зоозащитный центр «Киммела». Мы намерены постепенно стирать границу между изучением животных и их защитой. Эти две области не должны противопоставляться.
Я мысленно снял шляпу перед Лори. Чтобы принять такое радикальное решение, нужно огромное мужество. И хотя сам я мыслить подобным образом еще не привык, когда-нибудь и я к этому приду, это лишь вопрос времени.
Согласно полученным от Лори инструкциям, за образцами мозга предстояло наведаться в подвалы одного из самых мрачных зданий кампуса. В обиходе оно было известно как «старая стоматология», поскольку именно там до закрытия в 1990 году располагался стоматологический факультет Эмори. Антисемитская политика факультета получила широкую огласку, руководство принесло официальные извинения в документальном фильме, снятом, чтобы рассказать о темном прошлом университета. Однако, сколько бы факультет ни извинялся, на его здании все равно останется клеймо. Если приведения существуют, то по коридорам старой стоматологии они слоняются наверняка.
Мы с Питером отправились за образцами. Кабинет, где они хранились, никаких опознавательных знаков не имел, и, если бы Лори не дала нам подробных указаний, мы бы его ни за что не нашли. Открыв тугую дверь, мы включили свет.
Перед нами оказалась бывшая лаборатория, превращенная в банальную кладовку. На скамейках громоздились коробки, все было покрыто слоем пыли. Вытяжной шкаф, служащий для поглощения опасных газов и едких запахов, стоял приоткрытым, но характерного гудения, которое издает вытяжка в рабочем режиме, я не услышал. Внутри виднелся десяток поставленных друг на друга пластиковых ведерок. Сквозь пожелтевшие от времени стенки просматривалась налитая до половины темно-коричневая жидкость. Бумажные этикетки, прилепленные много лет назад, держались на последних молекулах клея.
— Кажется, это оно, — сказал я.
Питер кивнул.
Перчатки мы взять не додумались, но посмотреть, что в ведерках, было необходимо. Приподняв стеклянную дверцу шкафа, я вытащил одно из ведерок. В коричневой жидкости за выцветшим пластиком явно что-то плавало.
Когда я открыл крышку, в нос мне ударил запах формальдегида, глаза моментально заслезились, но даже сквозь пелену я разглядел внутри большой мозг. Не похожий ни на какой из виденных мной прежде. Крупный — по крайней мере раза в полтора крупнее человеческого. И круглый, как футбольный мяч. На этикетке значилось: «афалина». Перед нами был мозг бутылконосого дельфина, или афалины. Если не обращать внимания на мерзкую коричневую жидкость, выглядел он вполне прилично сохранившимся, особенно для образца, пролежавшего в этой гадости больше десяти лет.
— Неплохо, — сказал я. — Формалин нужно будет поменять, но, по крайней мере, мозг не расползся в кашу.
Мы вскрыли все контейнеры и отметили самые перспективные на вид экземпляры. Намереваясь проводить посмертную магнитно-резонансную томографию на образцах такой давности, мы вступали на неизведанную территорию. Ничего, скоро будет ясно, обогатит ли она нас сведениями об устройстве мозга дельфинов.
Погрузив полдесятка ведерок на тележку, мы с Питером покатили их в лабораторию, чтобы рассмотреть получше. Размер образцов в коллекции Лори варьировался от грейпфрута до футбольного мяча. Самые впечатляющие принадлежали афалинам: мало того что крупные, но и рисунок извилин гораздо более замысловатый, чем у человеческого мозга. Чем больше складок, тем больше площадь поверхности, а значит, эти извилины заключают в себе еще больше нервной ткани, однако среди них не было ничего похожего на центральную борозду — основной ориентир в мозге приматов, отделяющий лобные доли от всего остального.
До меня постепенно доходило, насколько трудную мы поставили перед собой задачу, и первоначальное мое воодушевление начало таять. Если мы не можем распознать даже основные части дельфиньего мозга, то как будем изучать его работу?
Как показало строение лобных долей, мозг дельфина лучше сравнивать с мозгом хищников, а не приматов. Присмотревшись повнимательнее, мы почти сразу разглядели крестообразную борозду, однако настолько сдвинутую вперед, что на лобную долю приходилось не более 10% общего объема. Это озадачивало. Как может животное, обладающее невероятным умом, иметь такие маленькие лобные доли? Либо специалисты по анатомии китообразных что-то напутали, либо мы ничего не знаем об отделах мозга, выполняющих когнитивные функции.
У нас было четыре образца мозга афалин и два поменьше — от дельфина-белобочки и пятнистого продельфина. Афалины считаются самыми умными и общительными из всех дельфинов, так что их мозг мы собирались сканировать в первую очередь. И хотя снаружи они выглядели вполне сохранными, главное скрывалось внутри. Быстрое сканирование покажет, не разжижена ли сердцевина.
Питер достал образцы мозга афалин из ведерок и подождал, пока стечет вязкая жидкость. Мы упаковали каждый в два запаивающихся пакета и направились к томографу, прижимая образцы к себе, словно жутковатые мячи для регби.
Выбрав самый приличный на вид, я уложил его в головную катушку томографа. Оно был хоть и крупнее человеческого, но зато без черепа, поэтому отлично поместился. Питер подложил под него куски пенопласта для устойчивости, и мы отправили мозг в тоннель томографа.
Я провел быстрое локализационное сканирование.
— Вроде неплохо. Серое и белое вещество видно по всей толщине.
Меня охватил азарт. Эти образцы, больше десяти лет пролежавшие в формалине, по-прежнему давали различимый сигнал. Воспрянув духом, я задал настройки для сканирования в высоком разрешении и нажал «пуск».
Аппарат начал предварительное сканирование. С щелчками и жужжанием он перестраивался на непривычную форму объекта, находящегося в его недрах. Спустя полминуты громкий гул возвестил о переходе к полноценному сканированию. Обычно оно длится минуты две, но, поскольку мозг афалины слишком большой и круглый, на него уйдет минут пять, не меньше. Нам оставалось только запастись терпением.
Наконец, по прошествии невероятно долгих пяти минут на экране появились снимки. Я принялся листать срезы, продвигаясь с самой нижней части мозга вверх.
— Вот мозжечок, — определил Питер. — Потрясающе подробно.
И действительно. Мозжечок выглядел как покрытый мелкой сеткой папоротник.
Когда мы дошли до коры, у меня снова опустились руки. Дыра на дыре. Даже невеликий специалист в анатомии мозга заметил бы, что образец явно некондиционный. Проследить нейронные связи в этом швейцарском сыре не представлялось возможным.
Мы просканировали еще два мозга афалины — в обоих обнаружилась такая же дистрофия тканей. Неудивительно, конечно, учитывая возраст образцов. Изъеденная дырами область располагалась ровно посередине между внешней поверхностью и желудочками. Судя по всему, из-за большого размера препарата формалин не смог пропитать его равномерно и выполнить свою консервирующую функцию, поэтому мозг со временем подвергся разложению.
Оставался только образец мозга белобочки. Примерно вдвое меньше мозга афалины, круглый, компактный, но заметно крупнее мозга морского льва. Мы поместили его в головную катушку и просканировали. К нашему огромному восторгу, он оказался целым и сохранным. Никаких дыр! Тогда мы запустили ДТВ — на всю ночь, до утра, так как процедура эта долгая.
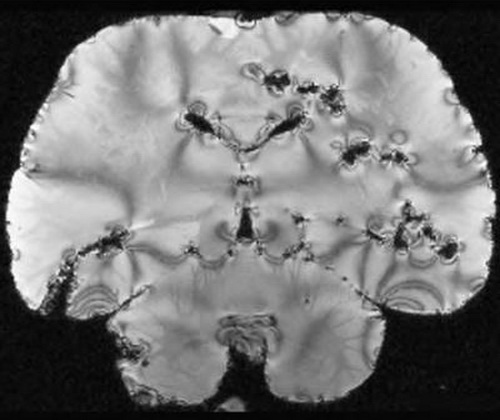
Покрытый дырами мозг афалины (Грегори Бернс)
Поскольку мозг был крупнее, чем у морского льва, на анализ данных потребовалось около недели. В первую очередь, проверяя, не ошиблись ли мы где-нибудь, проанализировали векторные карты. Мозг дельфина был настолько не похож на мозг сухопутных животных, что с таким же успехом мог принадлежать космическому пришельцу. Все сбито в большой круглый ком, все привычные ориентиры сдвинуты. Даже мозолистое тело выглядит странно. Неожиданно тонкое для мозга таких размеров, то есть связи между полушариями относительно немногочисленны.
Чтобы легче было сориентироваться, мы отобразили векторные поля в трех измерениях, убирая со снимков все, кроме белого вещества, и оставляя только подробную карту соединений. Заключенные в сферу нейронные тракты сложно было бы разглядеть без программного оборудования, способного сделать виртуальную «прозрачную» модель. И мы ее сделали.
Я смотрел на экран, где вращалось анимированное изображение, и испытывал благоговейный трепет — возбуждение ученого, которому удалось проникнуть в глубины неведомого. Вот нервные волокна, соединяющие кору со стволом мозга, они тянутся вертикально и на снимке раскрашены синим. Вот модель развернулась анфас, расправляя перед нами крылья височных долей, и мы увидели красные побеги черепных нервов, ответвляющиеся от ствола.
— Кажется, получилось, — сказал Питер.
Меня хватило только на то, чтобы кивнуть.
Убедившись, что результаты ДТВ выглядят многообещающе, мы обратились к волнующим нас научным вопросам. Мозг дельфина настолько отличался от всего виденного нами прежде, что мы с Питером слабо представляли, с чего начать и с какой стороны подступиться. Даже с мозгом морских львов было проще. Поскольку морские львы много времени проводят на суше, их мозг больше похож на мозг сухопутных млекопитающих, и мы не заметили там ничего непривычного по сравнению с собачьим. А вот дельфиний — это нечто совершенно иное.
Эхолокация не такое уж чуждое нам явление, как может показаться. Для нее необходимы две составляющие, и обе они у человека имеются — это производство звука и слух. Дельфины, разумеется, пользуются ею более виртуозно, чем человек, но за последние двадцать лет до нашего с Питером проекта в изучении эхолокации у дельфинов наметился большой прогресс, и теперь эта способность представлялась уже не такой загадочной, как прежде.

Трехмерная модель трактов в белом веществе мозга дельфина. Вид сбоку, мозг направлен влево (Грегори Бернс)
Эхолокация — биологический аналог системы гидролокации. Принцип ее довольно прост: издаваемый акустический сигнал отражается от подводных объектов и возвращается в виде эха. Время возвращения позволяет оценить расстояние до объекта, а по звуку эха можно определить размер объекта и его структуру. Искусственные гидролокационные системы достигли высокой степени технического совершенства, однако до точности дельфиньей эхолокации им по-прежнему далеко. Поэтому неудивительно, что ВМС США уже не первое десятилетие усиленно изучают дельфинов.
Дельфины и киты издают звук точно так же, как мы, с участием воздуха, но есть одно существенное отличие. Мы, как и остальные сухопутные животные, порождаем звук за счет вибрации воздуха в гортани. На выдохе воздушный столб проходит через голосовые связки, которые смыкаются и размыкаются, выпуская струи воздуха. Дальнейшее оформление этих потоков происходит при помощи горла, языка и губ. У дельфинов тоже есть гортань и голосовые связки, способные производить звук, однако в их случае это не главный механизм звукообразования. У всех зубатых китов, к которым относятся и дельфины, под дыхалом имеется пара органов, фамильярно называемых «обезьяньи губы» (или, в официальной терминологии, воздушные мешки). Когда дельфин выталкивает воздух через дыхало, воздушные мешки открываются и закрываются, вибрируя примерно как голосовые связки у сухопутных млекопитающих. Но в данном случае вибрирующий поток воздуха подается не в ротовую полость, а на жировую прослойку в лобной части головы, так называемый мелон. Он работает как акустическая линза, фокусируя и усиливая звуковой луч. Репертуар дельфинов поражает богатством и разнообразием. Помимо щелчков — коротких связок высокочастотных звуков, используемых для эхолокации, — в нем присутствуют свист и жужжание, которые, судя по всему, служат дельфинам для общения друг с другом.
Дельфинье щелканье попадает в ту часть звукового спектра, которая находится за пределами слышимости человека. Подросток способен различать частоты до 20 килогерц (кГц), тогда как преобладающая частота дельфиньих щелчков — более 100 кГц. Даже у кошек и собак предел слышимости ультразвука наступает где-то на 40 кГц. Однако при подводном звукоулавливании без высоких частот не обойтись. Если в воздухе звук распространяется со скоростью 340 м/с, то в морской воде — с запредельной 1500 м/с. Сухопутные млекопитающие определяют местонахождение источника звука по разнице во времени его улавливания каждым из ушей, но под водой это время настолько ничтожно, что разницы между прибытием низкочастотных звуков фактически нет. Поэтому для человека звук под водой доносится словно со всех сторон сразу. А вот с ультразвуком, которым пользуются дельфины, такой проблемы не возникает.
У дельфинов имеются уши, но слуховое отверстие у них шириной с булавочный прокол. Слышат они челюстью — звук передается через кости. В этом на самом деле нет ничего странного. У человека такой механизм слуха тоже есть. Если приложить что-нибудь вибрирующее, телефон например, к челюстной дуге, вы разберете звук. Форма головы дельфина не только обеспечивает ему стремительность, но и фокусирует встречные звуковые волны на широкой части челюсти. Такое устройство наделяет дельфинов максимальной восприимчивостью к звуку, поступающему спереди.
Исследования эхолокации у дельфинов неоднократно демонстрировали невероятные способности этих животных к распознаванию. В одном из экспериментов они различали толщину алюминиевых сфер, даже когда разница составляла не более 0,3 мм. Подобную чувствительность обеспечивает не только использование ультразвука, но и молниеносная работа мозга. Скорость обработки слуховых сигналов можно проверить, например, с помощью двух щелчков, постепенно сокращая разрыв между ними до тех пор, пока они не сольются в восприятии в один. Величина разрыва при слиянии и выступает показателем времени, которое требуется нервной системе, чтобы обработать входящую информацию. У человека оно составляет от 30 до 50 миллисекунд. У дельфинов — 264 микросекунды, то есть дельфин обрабатывает звук в сто с лишним раз быстрее, чем человек.
Хотя пока еще не до конца понятно, как именно дельфинам удается различать звуки с таким микроскопическим временным интервалом, можно предположить, что до того, как сигнал доберется до коры или даже до таламуса, он проходит тщательную обработку в стволе мозга. Возьмем оркестр, который традиционно настраивается по ноте ля первой октавы. Эта нота обозначается как А440, поскольку ее частота — 440 Гц. Но даже музыканту с абсолютным слухом сложно будет определить без сопоставления с камертонным эталоном, не сползает ли его инструмент на 439 Гц. При совместной игре разница частот составляет 1 Гц и называется частотой биений, потому что мы слышим при этом легкое биение, в данном случае раз в секунду. Исходя из времени обработки звука у дельфинов, можно вычислить, что они различают частоту биения до 4 кГц.
Как видим, все эти компоненты слуховой системы, которые поначалу кажутся такими непривычными, вполне соотносятся с компонентами нашего собственного мозга. А значит, вопреки тому, что утверждал Нагель, не так уж и трудно представить себе, каково быть летучей мышью или дельфином.
По сути, эхолокация не только не делает дельфинов непостижимыми, а, наоборот, предоставляет нам идеальную возможность определить, что можно узнать о субъективных ощущениях животного по его мозгу. Еще в середине XX века анатомы установили, что проводящие слуховые пути в мозге дельфина достаточно обширны. Однако до сих пор было мало изучено, как дельфины при помощи отраженного звука создают когнитивную картину окружающего их мира, если, конечно, здесь уместно слово «картина».
Нам нужно было узнать, какая область коры головного мозга принимает звуковые данные. У сухопутных млекопитающих слуховой нерв передает всю акустическую информацию в ствол мозга. Там звуковой поток расщепляется надвое, одна часть остается на той же стороне, откуда поступила, другая направляется на противоположную сторону ствола. Затем, проходя через цепочку ядер, эти потоки движутся вверх к таламусу.
Непосредственно перед тем, как достичь таламуса, звуковые импульсы направляются в сферическое ядро под названием «нижнее двухолмие». У млекопитающих оно настолько крупное, что правая и левая его части образуют пару внушительных выпуклостей на тыльной части ствола. Прямо над ним находится верхнее двухолмие, принимающее зрительную информацию, и по соотношению размеров нижнего и верхнего двухолмия можно примерно представить себе относительную роль слухового и зрительного восприятия у данного животного. У дельфинов нижнее двухолмие очень крупное, и оттуда уже совсем близко до таламуса.
Таламус располагается в центре мозга, между стволом и корой. У человека он величиной примерно с небольшую сливу. Таламус четко отделен от коры, а внутренняя структура представляет собой десятки отдельных ядер. Эти ядра служат промежуточными станциями при передаче сигналов между корой и другими отделами нервной системы. Часть ядер получает сигналы от спинного мозга, передающего сенсорные импульсы от органов тела к мозгу. Другие обеспечивают кольцевое взаимодействие импульсов, принимая информацию от базальных ядер и мозжечка и ретранслируя в кору, — так, судя по всему, мозгу проще координировать передачу. У приматов внушительная задняя часть таламуса, называющаяся подушкой, отвечает за зрение. Слуховая информация поступает в медиальное коленчатое тело.
Слуховые пути достаточно хорошо картированы у человека и крысы, однако у дельфинов к тому моменту, как мы с Питером ими занялись, в этом направлении сделано было мало. Федеральный Закон об охране морских млекопитающих жестко ограничивает научную работу с дельфинами и другими китообразными, и инвазивное исследование мозга дельфинов под эти ограничения тоже подпадало. В начале 1970-х годов советские ученые попытались картировать мозг дельфина, внедряя в него электроды и наблюдая, какие области реагируют на звук. Кроме того, они сделали попытку разметить связи между корой и таламусом у дельфинов, вводя им радиоактивные вещества и прослеживая путь этой радиоактивной метки вдоль аксонов.
В результате этих обрывочных исследований дельфиньих слуховых путей складывалась довольно странная картина. У всех сухопутных млекопитающих слуховая информация передается от медиального коленчатого тела в латеральном направлении, к височным долям. У человека, например, в верхней части височной доли имеется область под названием «извилина Гешля», которая эту информацию принимает. Существует даже карта волокон, несущих импульсы различных частот. Но, как свидетельствовали данные советских исследований, у дельфинов слуховая информация поступает в ту область коры, которая расположена ближе к макушке головы, почти в самой дальней части мозга, и у остальных обычно принимает поток зрительной информации. Согласно одной из теорий, поскольку слуховые данные имеют для дельфина первостепенное значение, в ходе эволюции слуховая кора его мозга значительно разрослась и сместилась в те области, которые у сухопутных млекопитающих отданы под обработку зрительных данных.
Мы с Питером не знали, где у дельфинов располагается слуховая кора, однако нижнее двухолмие распознали без труда. Его трудно было не заметить. Оно находилось непосредственно над стволом и представляло собой две четко выделяющиеся сферические структуры диаметром около сантиметра. Как и при работе с мозгом морских львов, мы прибегли к помощи цифровых меток, только теперь поместили их не в гиппокампе, а в нижнем двухолмии и дали программе задание воспроизвести отслеживаемые ими тракты.
Тракты шли в двух направлениях. Симулятор превосходно отследил нисходящие пути, устремляющиеся вниз по стволу обратно к слуховому нерву, тем самым подтвердив правильность нашего подхода. В противоположном направлении тянулся толстый пучок волокон, связывающий нижнее двухолмие с таламусом, — тоже в полном соответствии с нашими прогнозами. Но дальше тракты, повернув вбок, вели в латеральном направлении — к височным долям. В точности как у сухопутных млекопитающих.
Когда Питер показал мне снимки, я обрадовался: «Это же замечательно! Слуховые пути совсем как у остальных».
Однако Лори, которой мы отослали сканы, перезвонила мне в недоумении.
— Так не должно быть. У вас все не по учебнику.
— То есть?
— Слуховые тракты должны вести к темени, — объяснила она. — А не к височным долям.
Но я был уверен в правильности полученных результатов.
— Может, это учебники ошибаются?
Вопрос был принципиальный. «По учебникам» у дельфинов идет крупная нервная магистраль от слуховой системы в зрительную часть коры, что кардинальным образом отличается от устройства человеческого мозга. Однако результаты нашей ДТВ говорили о том, что разница не так велика. Мы обнаружили слуховой проводящий путь, идущий к височной доле, а это значит, что дельфины не «видят» звук, а слышат точно так же, как мы. Между тем эхолокация не сводится к одному лишь слуху. Эхолокация — это активный процесс, включающий и порождение, и восприятие сигнала. Дельфины звуком не видят, а «рисуют».
Пока мы просканировали только один мозг. И хотя результаты оказались сенсационными, нам требовалось убедиться, что это не случайность. Необходимо было исследовать еще один образец.
Единственный оставшийся в нашем распоряжении прилично сохранившийся мозг представлял собой экземпляр размером примерно с грейпфрут, принадлежавший пантропическому пятнистому продельфину. Как следует из названия, пантропические продельфины обитают в теплых водах по обе стороны экватора. В восточной части США они, следуя за Гольфстримом, иногда забираются достаточно далеко на север — к штату Мэн. Наряду с белобочкой пятнистый продельфин — самый многочисленный из обитающих здесь китообразных. Его популяция пережила даже массовое истребление в восточной части Тихого океана. С 1950-х до конца 1980-х годов миллионы дельфинов гибли при промышленном вылове тунца сетями. До применения этого способа популяция дельфинов исчислялась тремя или четырьмя миллионами. Сейчас она насчитывает пятьсот тысяч особей.
За время нашей работы с мозгом морских львов и дельфинов Питер достаточно поднаторел в заливке образцов желатином и запуске сканирования на всю ночь. На обработку мозга пятнистого продельфина ушло несколько дней. Сигнал получился не таким четким, как у белобочки, то есть диффузионные показатели сильнее искажались тепловым движением. И тем не менее, когда мы поместили виртуальные метки в нижнее двухолмие, симулятор выдал точно такой же путь к таламусу и височным долям, как у белобочки. Если рассматривать левую и правую стороны мозга по отдельности, то височный слуховой путь обнаруживался в четырех полушариях из четырех. Наше открытие подтверждалось.
Результаты ДТВ дали нам дорожную карту воспринимающей стороны слуха дельфинов. В обоих просканированных образцах мозга основной путь от таламуса шел к височным долям, а не к теменной части. В целом этот тракт достаточно типичен для мозга млекопитающих, и, соответственно, там, где он заканчивается, располагается слуховая кора.
Но куда поступает информация оттуда?
Чтобы это выяснить, мы попросту поместили очередную виртуальную метку в обнаруженную нами область слуховой коры и принялись отслеживать идущие оттуда тракты. Теперь они вели назад и вверх.
Получившаяся карта указывала на две слуховые области — одна в височных долях, как у сухопутных млекопитающих, а другая ближе к темени, рядом со зрительной корой. Среди млекопитающих схожая организация наблюдается только у летучих мышей. Поскольку летучие мыши пользуются эхолокацией на суше, механизм ее вполне понятен. У летучих мышей имеется первичная слуховая зона в височных долях, точно такая же, как у дельфинов, но кроме нее есть еще вторичная и третичная слуховая кора непосредственно над височными долями и позади. У некоторых видов летучих мышей содержащиеся в этих вспомогательных слуховых областях нейроны специфически активируются на разные интервалы возвращения эха, формируя в итоге когнитивную карту расстояний до окружающих объектов. У некоторых летучих мышей присутствует еще одна прилегающая область, которая реагирует на изменение звука эха. Эти мыши умеют регулировать издаваемый ими звук, подстраиваясь под объект эхолокации. Это называется «частотная модуляция», как в радиовещании в диапазоне УКВ (FM).
Самое примечательное, что при всем сходстве между мозгом летучих мышей и дельфинов эти животные не связаны близким родством. Чтобы отыскать у них общего предка, придется вернуться в прошлое минимум на восемьдесят миллионов лет. Ближайшие сухопутные родственники дельфинов — парнокопытные, то есть свиньи, коровы, козы, овцы, и у них эхолокации не обнаруживается (хотя другие представители копытных, гиппопотамы, общаются при помощи издаваемых под водой щелчков). В данном случае перед нами классический пример параллельной эволюции. Эхолокация развивалась у летучих мышей и дельфинов независимо, но, поскольку задача стояла одинаковая, решения тоже получились аналогичными, только одно — для воздушной среды, а другое — для водной. Эту картину подтверждает и исследование генома дельфинов и летучих мышей. Гены, связанные со слухом и зрением, роднят этих животных между собой гораздо больше, чем следовало бы ожидать при таком далеком общем предке.
Сознавать, что мы отыскали еще один кусочек мозаики, еще одно звено цепи, соединяющей дельфинов с сухопутными животными, было приятно. Отчасти нас воодушевляла просто радость познания. Древо земной жизни поражает великолепием, и, когда удается обнаружить очередную связь между его ветвями, становится яснее, где на нем располагаюсь лично я, представитель семейства гоминид. Результаты исследования мозга животных при помощи таких сложных инструментов, как диффузионная МРТ, говорят, что сходства между нами больше, чем различий. Даже такое чуждое, казалось бы, явление, как эхолокация, на самом деле не так уж и необычно, если разобрать его на составляющие. Нагель выступал против редукционизма, но ведь именно с помощью редукции к проводящим путям в белом веществе нам удалось обнаружить нечто общее между дельфинами и человеком.
А еще радостно было от того, что наша работа имела и философское значение. Философы любят рассуждать о «квалиа» — субъективном ощущении какого-то понятия, например красного цвета. Представьте себе орхидею, переливающуюся всеми оттенками от розового до фиолетового. Ни в одном языке не хватит слов, чтобы передать ощущения от этого цветка целиком и полностью. Даже названий оттенков не хватит, а уж о запахе и говорить нечего. Так что если квалиа существует, каждый из нас навеки заперт в мире собственных ощущений, не имея ни малейшей возможности проверить, ощущает ли он то же самое, что и остальные. Если в нашем распоряжении только слова, как убедиться, что мой «красный» не выглядит для вас «синим»?
Доводы любого, кто тянет из рукава козырь квалиа, всегда предсказуемы: как ни совершенствуй знания о физической стороне того или иного процесса, ощущения от него мы все равно не сможем передать. И не важно, что красный цвет точно и досконально описан как электромагнитное излучение с длиной волны 700 нанометров. Физика не передает ощущение красного.
Но я считал иначе. Физика и биология вполне способны передать то, что не выразишь словами. Свидетельством тому — нашумевший интернет-прикол 2015 года с платьем — то ли сине-черным, то ли бело-золотым. Диванные философы ухватились за него как за доказательство квалиа, ведь для каждого из нас платье существовало в своей собственной версии. Однако научное объяснение оказалось куда более прозаичным. Рэндалл Манро в своем блоге XKCD с помощью простой иллюстрации показал, как на наше восприятие платья влияет фоновый цвет изображения. Как и большинство систем организма, цветовое восприятие оперирует величинами относительными, а не абсолютными. Объект воспринимается как синий или золотой в сравнении с чем-то еще. И самое логичное объяснение разницы восприятия заключается в том, что смотрящие брали за ориентир разные элементы фона. Редукция к этим элементам уничтожает весь таинственный флер квалиа. Собственно, даже свое собственное восприятие вы можете изменить, если попробуете сосредоточить взгляд на разных участках изображения.
То же самое и с мозгом дельфина. Эхолокация — это не чужеродная способность, которую нам ни за что не осознать. Эхолокация — это всего-навсего усиленный вариант неких навыков восприятия, которые имеются и у человека. Однако не пора ли нам перейти от органов чувств к чему-нибудь более глубинному? Поискать межвидовое родство в когнитивных процессах? Например, в области общения и социальных отношений.
Назад: Глава 5 Зачатки способностей
Дальше: Глава 7 Буриданов осел

