3. белки
— А вот товарищ Амперян говорит, что без белка жить нельзя, — сказал Витька, заставляя струю табачного дыма сворачиваться в смерч и ходить по комнате, огибая предметы.— Я говорю, что жизнь — это белок, — возразил Эдик.— Не ощущаю разницы, — сказал Витька. — Ты говоришь, что если нет белка, то нет и жизни.Аркадий и Борис Стругацкие. Понедельник начинается в субботу
В разговорах о современной биологии слово “белок” звучит очень часто. Все знают, что белки — важнейшие питательные вещества. Но одновременно это еще и сложные биохимические машины, выполняющие в организме множество самых разных функций: дыхание, пищеварение, считывание наследственной информации, сокращение мышц, защита, восприятие света, передача сигналов... проще сказать, чего белки не делают. Что же это, в конце концов, такое?
Начнем с того, что белки, или протеины, — это огромные молекулы, входящие в состав абсолютно всех современных живых организмов. История их названия, честно говоря, довольно запутанна. Сам термин “белок” (albumin) вошел в употребление еще в XVIII веке, и относился он тогда к веществам, подобным всем известному белку куриного яйца. Что касается термина “протеин” (protein), то его придумал знаменитый шведский химик Йёнс Якоб Берцелиус. Кроме этого, Берцелиус открыл несколько новых химических элементов, установил формулы ряда органических кислот, разобрался в явлении электролиза и сделал еще много другого. В частности, именно Берцелиус открыл явление изомерии и ввел само понятие “изомеры”, уже нам знакомое (см. главу 1). Да и термин “органическая химия” тоже принадлежит ему.
Так вот, в 1838 году Берцелиус предложил назвать некоторые органические вещества “протеинами”. Слово это произведено от греческого πρώτειος, “первичный”. Придумывая свой термин, Берцелиус предполагал, что “протеины” — это некие первичные строительные блоки живых организмов, и был, как мы сейчас знаем, совершенно прав.
В русском языке “белок” и “протеин” — строгие синонимы. По буквальному смыслу “протеин”, конечно, точнее, чем “белок”. Но как-то уж так сложилось, что в нашей научной литературе слово “белок” употребляется гораздо чаще, и мы будем этому следовать. Скорее всего, дело тут просто в том, что слово “белок” проще для восприятия и привычнее на слух.
Полимеры
В состав белков входят углерод, водород, кислород, азот и, как правило, еще сера. Ничего особенного в таком химическом составе нет. Гораздо удивительнее другое. Еще в XIX веке химики обнаружили, что молекулы белков буквально гигантские. По размеру, то есть по количеству атомов, они в сотни раз превосходят молекулы большинства других органических веществ. Дело в том, что белки относятся к полимерам — молекулам, состоящим из множества однотипных (но не обязательно совершенно одинаковых) повторяющихся звеньев, ковалентными связями соединенных друг с другом (см. рис. 3.1А). Такие звенья называются мономерами. Полимеры могут распадаться на отдельные мономеры, а могут и собираться из них вновь. Эти процессы играют огромную роль в биологическом обмене веществ, в ходе которого то и дело одни полимеры расщепляются, а другие строятся из мономеров заново. Иногда полимеры называют макромолекулами, то есть попросту “большими молекулами”.
Очень простой пример полимера — полиэтилен, тот самый, из которого делают упаковочную пленку, пакеты, изоленту и тому подобные вещи. Это обычный углеводород, имеющий, однако, очень длинные молекулы (гораздо более длинные, чем все, что мы видели до сих пор). Формула полиэтилена следующая: CH3–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2–... За многоточием тут может скрываться цепочка из нескольких тысяч атомов углерода (разумеется, с присоединенными к ним атомами водорода, которые всегда заполняют все валентности, не заполненные другими атомами). Повторяющимся звеном в полиэтилене является группа –CH2–CH2– (с двумя атомами углерода, а не с одним, потому что при получении полиэтилена он “сшивается” именно из двухуглеродных молекул). Это и есть его мономер.
И в живой, и в неживой природе есть много всевозможных полимеров, состоящих из самых разных типов мономерных звеньев. И как правило, эти звенья гораздо сложнее, чем в полиэтилене. Например, во многих биологических полимерах мономерами являются сахара (см. главу 6). В белках же мономеры особые, свойственные только им. Это — аминокислоты.
Альфа, бета, гамма...
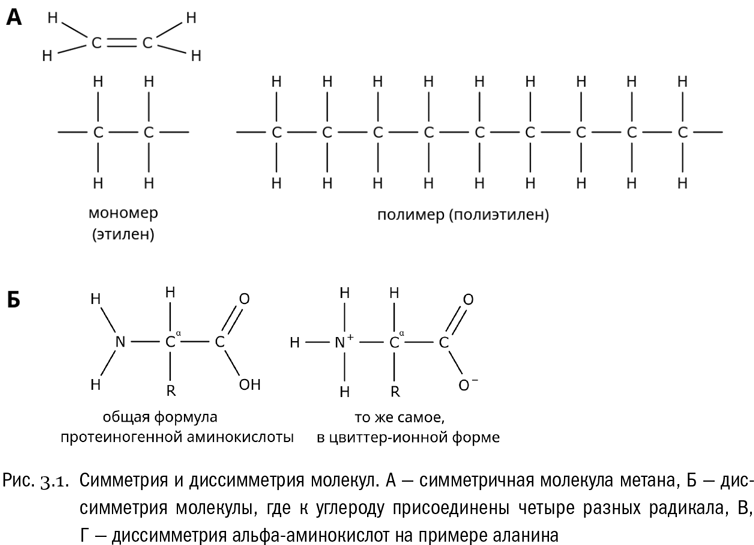
Аминокислота, как мы знаем, — это вещество, в молекуле которого одновременно есть карбоксильная группа и аминогруппа (см. главу 1). Особенность аминокислот, входящих в состав белков, в том, что эти две группы обязательно присоединены к одному и тому же атому углерода. Такие аминокислоты называются альфа-аминокислотами (см. рис. 3.1Б). Если в какой-нибудь аминокислоте карбоксильная группа и аминогруппа связаны с разными атомами углерода, то это не альфа-аминокислота и в состав белков она входить не может.
Почему альфа-аминокислоты называются “альфа” и при чем тут вообще греческие буквы? Дело вот в чем. Атомы углерода, образующие аминокислоту, принято обозначать греческими буквами по порядку, считая от карбоксильной группы (сама она в счет не идет). Таким образом, первый атом углерода после карбоксила — это альфа-атом, второй — бета-атом, третий — гамма-атом и т.д. И в зависимости от того, к какому атому углерода у них присоединена аминогруппа, аминокислоты делятся на альфа-аминокислоты, бета-аминокислоты и прочие.
Например, представим себе аминокислоту с формулой NH2–CH2–CH2–CH2–COOH (см. далее рис. 3.8). Аминогруппа тут присоединена к третьему атому углерода, считая от карбоксильной группы, то есть к гамма-атому. Значит, это гамма-аминокислота. В данном случае она называется гамма-аминомасляной кислотой (ГАМК). Эта аминокислота есть в организмах большинства животных. Во-первых, она является промежуточным продуктом обмена веществ, а во-вторых, служит нейротрансмиттером, то есть веществом, передающим сигнал между нервными клетками. Именно с нарушением ГАМК-эргической (то есть обусловленной ГАМК) передачи нервных импульсов связано действие одного из самых сильных растительных ядов — яда цикуты, которым в свое время отравили Сократа. Но вот в состав белков ГАМК, в отличие от альфа-аминокислот, никогда не входит.
Пептидная связь
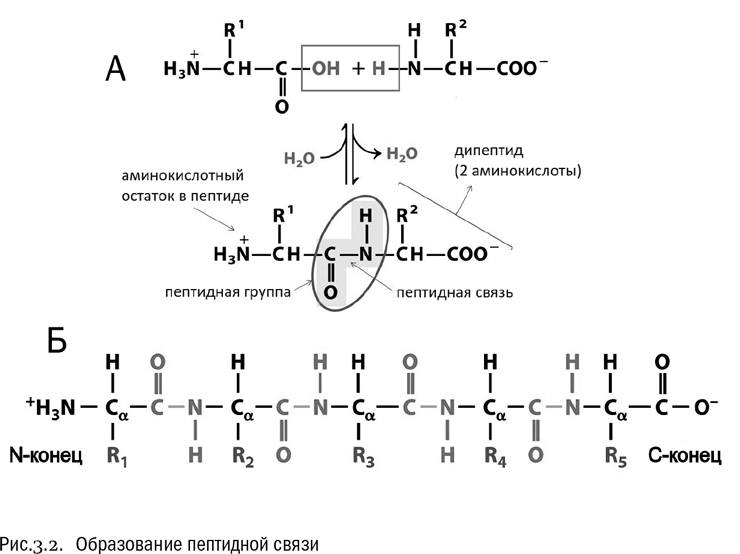
Как альфа-аминокислоты объединяются в белок? Очень просто: карбоксильная группа одной аминокислоты связывается с аминогруппой другой (см. рис. 3.2А). От карбоксильной группы отщепляется гидроксил (–OH), а от аминогруппы — водород (–H). Эти отщепленные фрагменты тут же соединяются и дают воду (H–O–H), а остатки карбоксильной группы и аминогруппы замыкаются по освободившимся валентностям друг на друга, образуя новую группу –CO–NH–. Вот через нее-то две аминокислоты и соединяются между собой.
Группа –CO–NH– называется пептидной группой, а связь между углеродом и азотом в ней — пептидной связью. Цепочка аминокислот, соединенных пептидными связями, называется пептидом (см. рис. 3.2Б). Это более широкое понятие, чем белок. Все белки — пептиды, но не все пептиды — белки.
Реакция образования пептида в принципе обратима: он может как синтезироваться, так и распадаться обратно на отдельные аминокислоты. На одном конце пептида находится свободная аминогруппа, на другом — свободная карбоксильная группа. Для краткости конец пептида со свободной аминогруппой принято называть N-концом, а конец со свободной карбоксильной группой — C-концом.
Короткие пептиды называют или по числу аминокислотных остатков (два остатка — дипептид, три — трипептид, четыре — тетрапептид...), или собирательно — олигопептидами. Длинные пептиды с многими десятками аминокислотных остатков называют полипептидами. Все белки — полипептиды. Аминокислотных остатков в них обычно даже не десятки, а сотни. “Средний” белок, типичный для живой природы, включает примерно 300–350 аминокислот. Белок из 200 аминокислот считается небольшим. Неудивительно, что белковые молекулы поразили когда-то химиков своими размерами.
Разнообразие и единство
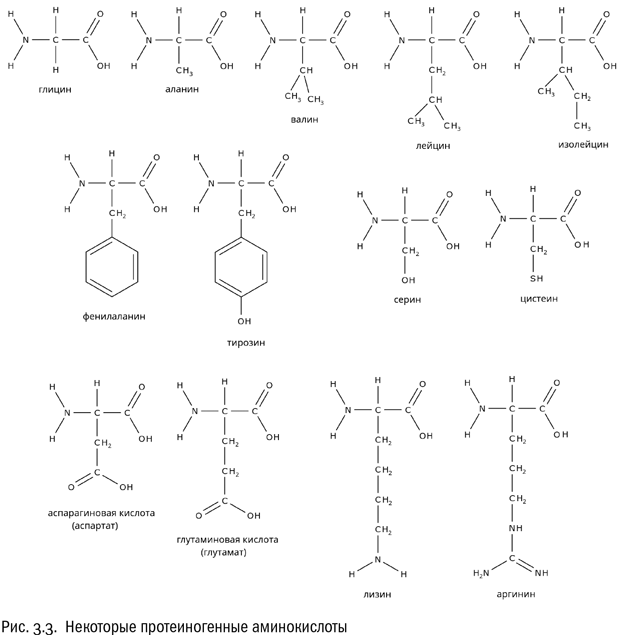
В состав белков входит 20 стандартных аминокислот, одних и тех же у всех живых организмов. Как мы уже знаем, все они — альфа-аминокислоты, а это значит, что их общую формулу можно записать вот как: R–CH(NH2)–COOH. Буква R тут, как всегда, обозначает радикал, то есть изменяемую часть молекулы.
Аминокислоты, образующие белки, называют протеиногенными, от уже знакомого нам слова “протеин”. Две самые простые протеиногенные аминокислоты — глицин (где радикал — атом водорода) и аланин (где радикалом служит метильная группа –CH3). У других аминокислот радикалы сложнее. Для читателей-эрудитов добавим, что все нестандартные аминокислоты (селеноцистеин, пирролизин, гидроксилизин, гидроксипролин) так или иначе являются производными стандартных и нас пока не интересуют. А стандартных — ровно 20.
Свойства любого пептида зависят не только от того, какие аминокислоты в него входят, но и от того, в каком порядке они там расположены. Например, представим себе дипептид, состоящий из глицина и аланина. Как он будет выглядеть? Если в создании пептидной связи примут участие карбоксильная группа глицина и аминогруппа аланина, дипептид будет таким: NH2–CH2–CO–NH–CH(CH3)–COOH. Но возможен и другой случай, когда пептидную связь образуют, наоборот, карбоксильная группа аланина и аминогруппа глицина. Тогда пептид получится вот таким: NH2–CH(CH3)–CO–NH–CH2–COOH. Как видим, это два разных соединения. В белках, состоящих из сотен аминокислот, порядок расположения этих аминокислот не менее важен — только вот возможных вариантов там намного больше.
Глядя на формулы, легко убедиться, что два наших дипептида — не что иное, как изомеры (см. главу 1). То же относится к любым пептидам, отличающимся друг от друга порядком расположения одних и тех же аминокислотных остатков. И число таких изомеров в случае с длинными пептидами может быть огромным. Например, можно вычислить, что для декапептида, состоящего из 10 разных аминокислот, число возможных изомеров равно 3 628 800. А ведь декапептид — это даже не белок. Для любого крупного белка число изомеров будет в буквальном смысле астрономическим. Вот почему разных белков так много.
Порядок расположения аминокислот крайне важен для того, чтобы белок правильно выполнял свою функцию. Он должен быть таким же точным, как порядок букв в напечатанной фразе. Единственная замена или перестановка вполне может сделать белок совершенно “бессмысленным”, то есть бесполезным для организма. Между тем никакими чисто химическими средствами такую точность синтеза белка обеспечить невозможно. Первым, кто над этим всерьез задумался, был выдающийся русский биолог Николай Константинович Кольцов. “Молекула октокайдекапептида, состоящая из 18 аминокислот, может иметь около триллиона изомеров, а изомеры сложных белковых молекул должны исчисляться центильонами (число из 600 цифр). Представляется совершенно невероятным, чтобы синтез определенного изомера белков определялся исключительно внешними условиями реакции”, — писал Кольцов еще в 1927 году. Из этого следовал важнейший вывод: информация, задающая порядок аминокислот в белке, непременно должна храниться где-то в организме. Она должна копироваться, передаваясь из поколения в поколение, и считываться по первому запросу, когда тот или иной белок понадобится создать. Все это, как мы сейчас понимаем, совершенно верно. О том, как в действительности работают эти механизмы, мы узнаем из главы 9.
В отличие от некоторых других полимеров, белки никогда не ветвятся. Любой белок — это строго линейная цепочка аминокислот. Сами аминокислоты, входящие в состав белка, принято обозначать буквами: например, глицин обозначается буквой G, аланин — буквой A и т.д. Поэтому формулу любого белка можно записать в виде простой последовательности букв, соответствующих аминокислотам. На самом деле так обычно и делают.
“Кирпичики”, из которых состоит жизнь
Итак, мы видим, что аминокислоты, входящие в состав белков, построены по одной схеме. В любой из этих аминокислот есть карбоксильная группа и аминогруппа, присоединенные к центральному атому углерода (тому, который мы назвали альфа-атомом). Кроме того, к центральному атому углерода всегда присоединен атом водорода. Таким образом, из четырех валентностей альфа-углеродного атома три всегда заняты одними и теми же группами — карбоксильной, аминогруппой и атомом водорода. В этих частях молекул никакого разнообразия нет.
А вот четвертая валентность альфа-атома углерода занята изменчивой группой, которую мы для удобства назвали радикалом (–R). По ней-то аминокислоты и различаются (см. рис. 3.3).
Есть несколько аминокислот, у которых радикалы чисто углеводородные: аланин, валин, лейцин, изолейцин, фенилаланин. Две из них — лейцин и изолейцин — являются изомерами друг друга, потому так и названы. Они отличаются всего лишь положением одной метильной группы (–CH3). Чисто углеводородные радикалы плохо взаимодействуют с водой, но хорошо друг с другом. Иначе говоря, эти радикалы — гидрофобные.
Есть аминокислоты, у которых в состав радикала входит гидроксильная группа –OH (серин, тирозин) или аналогичная ей, но содержащая вместо атома кислорода атом серы тиольная группа –SH (цистеин). В таких радикалах есть полярные связи, а потому они взаимодействуют с водой гораздо лучше. Эти радикалы — гидрофильные.
Все аминокислоты, перечисленные нами до сих пор, называются нейтральными. Это означает, что в водном растворе их молекулы электрически не заряжены. Мы уже видели, что в любой аминокислоте есть карбоксильная группа, свойства которой кислотные, и аминогруппа, свойства которой, наоборот, основные (см. главу 1). Попадая в воду, карбоксильная группа отдает протон и становится заряжена отрицательно (–COO–), а аминогруппа присоединяет протон и становится заряжена положительно (–NH3+). Суммарный заряд молекулы аминокислоты в результате остается равным нулю. Это — нейтральная молекула.
А что, если карбоксильных групп две? Тогда эта аминокислота будет в растворе заряжена отрицательно. И действительно, в состав белков входит пара таких аминокислот — это аспарагиновая и глутаминовая кислоты. У них обеих есть карбоксильная группа не только при альфа-атоме, но еще и в радикале. И, соответственно, этот радикал несет дополнительный отрицательный заряд.
Для краткости аспарагиновую и глутаминовую кислоты часто называют, соответственно, аспартатом и глутаматом. Тут надо пояснить одну тонкость, связанную с названиями веществ. Аспартат и глутамат — на самом деле названия не кислот, а их анионов или (что то же самое) их солей. Например, глутамат — это соль глутаминовой кислоты. В биохимии этим сплошь и рядом пренебрегают, используя названия кислот и солей как синонимы. Ведь что такое соль? Это кислота, у которой на месте протона оказался любой другой катион. Если же она диссоциирована и не имеет никаких катионов вообще (а это бывает в растворах очень часто), то за ней обычно ради удобства оставляют название соответствующей соли. Именно это мы на примере аспартата с глутаматом и видим. Название соли — это название аниона, в виде которого молекула реально существует в воде.
Глутамат (будем отныне называть его так) интересен не только тем, что участвует в образовании белков. В организмах подавляющего большинства животных он служит еще и нейротрансмиттером, причем одним из важнейших. В нервной системе человека глутамат используют для передачи возбуждения примерно 40% нейронов — это очень много! Почти все основные информационные потоки в нашем мозге идут посредством выделения глутамата, служащего для нервных клеток возбуждающим сигналом. Выше мы упоминали, что нейротрансмиттером является и еще одна протеиногенная аминокислота, а именно глицин (см. главу 1). Но действие глицина тормозящее (то есть успокаивающее), а глутамата — именно возбуждающее. Поскольку глутамат входит в состав каких угодно белков, то его очень много в пище, но пищевой глутамат в мозг почти не попадает — нервные клетки синтезируют его сами.
Кроме того, к глутамату очень чувствительна наша вкусовая система. Обычно считается, что есть пять основных вкусов, для которых на языке существуют отдельные типы рецепторов: соленый, кислый, сладкий, горький и выделенный в начале XX века вкус умами. Соленый — это вкус поваренной соли, кислый — протонов (H+), сладкий — сахаров. Горький вкус — самый сложный, он не привязан к какому-то одному классу молекул и возникает как реакция на любое вещество из большой и разнообразной группы зачисленных мозгом в “ядовитые”, это эволюционно выработанный механизм защиты от токсичной пищи. Ну а умами — это не что иное, как вкус глутамата. Судя по всему, в ходе эволюции органов чувств позвоночных животных именно глутамат был выделен как индикатор вкуса белков (важнейших питательных веществ как-никак). Вот почему на языке для него есть особые рецепторы. Ощущение вкуса глутамата — это эволюционно выработанный сигнал о том, что в рот попало нечто белковое.
Глутамат часто добавляют в пищу, причем как в виде кислоты (пищевая добавка E620), так и в виде натриевой соли (пищевая добавка Е621). И раз уж мы заговорили об этом веществе, воспользуемся случаем, чтобы развеять несколько связанных с ним заблуждений. Может быть, кому-то пригодится.
Итак, во-первых, утверждение, что глутамат — усилитель вкуса, неточно. Выражение “усилитель вкуса” могло бы относиться к некоему (вымышленному) веществу без собственного вкуса и запаха, обладающему свойством обострять вкус любой еды. Глутамат этого не делает: у него просто есть свой вкус, точно так же, как у сахара или у соли. Механизмы восприятия вкуса глутамата и вкуса, допустим, того же сахара принципиально не отличаются друг от друга. Просто сахар воспринимается одними рецепторами, а глутамат — другими.
Во-вторых, неверно часто встречающееся мнение, будто “натуральный” глутамат (предположительно безобидный) — это совсем не то, что глутамат “искусственный” (предположительно вредный и опасный). Люди, которые так думают, просто не знают, о каком веществе идет речь. А мы с вами теперь знаем. Глутамат — это не какая-нибудь загадочная сложная смесь (состав которой действительно мог бы варьироваться), а одно-единственное химическое соединение, описываемое незатейливой формулой. Вот она, эта формула: HOOC–CH2–CH2–CH(NH2)–COOH. Только и всего. Глутамат, полученный искусственно, не отличается от полученного готовым из природных продуктов, потому что отличаться там нечему.
В-третьих, глутамат, получаемый с пищей, едва ли опасен для нервной системы прежде всего потому, что он в нее почти не проникает — это обеспечивается специальным физиологическим барьером. Нервные клетки синтезируют глутамат самостоятельно. К тому же в белковых продуктах (таких, как творог, мясо или соя) глутамата наверняка больше, чем попадает его в еду в качестве пищевой добавки, — просто потому, что это составная часть любых белков.
В-четвертых, на глутамат нет аллергии. Аллергию вызывают чужеродные вещества, а не такие, которые жизненно необходимы и всегда присутствуют в организме в высоких концентрациях, — а глутамат как раз относится к последним.
Так что опасность глутамата — это, судя по всему, типичный современный миф.
Однако вернемся к другим аминокислотам. Если есть отрицательно заряженные аминокислоты, логично ожидать, что существуют и положительно заряженные. Это действительно так. Пример положительно заряженной аминокислоты — лизин, имеющий формулу NH2–CH2–CH2–CH2–CH2–CH(NH2)–COOH. Как видим, у лизина в радикале есть дополнительная аминогруппа, которая ведет себя так, как аминогруппе и положено: приобретает протон. Еще одна положительно заряженная аминокислота — аргинин, радикал которого включает довольно редкую (больше она нам нигде не встретится) гуанидиновую группу –NH–C(NH)–NH2, тоже охотно присоединяющую протон.
Таким образом, аминокислоты, из которых состоят белки, можно поделить на четыре категории: гидрофобные, гидрофильные нейтральные, положительно заряженные и отрицательно заряженные. Разумеется, все эти различия касаются только той части молекулы аминокислоты, которую мы назвали радикалом (R). “Базовая” часть (включающая атом углерода, атом водорода, карбоксильную группу и аминогруппу) во всех рассмотренных нами протеиногенных аминокислотах одна и та же. Особое положение занимает разве что самая простая из всех возможных аминокислот — глицин, у которого вместо радикала атом водорода.
Аминокислоты и связи
Итак, любой белок — это полипептид, то есть цепочка аминокислот, соединенных пептидными связями. На самом деле теоретически можно придумать полипептид, не являющийся белком, но мы сейчас поступим проще и будем считать, что эти слова — синонимы. В подавляющем большинстве тех случаев, которые нам могут встретиться, так оно и есть.
Для начала представим, что молекулу полипептида бросили в воду. Очевидно, что она не останется там вытянутой в прямую линию, а будет как-то сворачиваться. Это сворачивание будет зависеть от взаимодействия аминокислотных остатков как с молекулами воды, так и между собой. В целом пептидная цепь устроена довольно просто: ее “скелет” образуют пептидные группы, соединяющие между собой альфа-атомы углерода, а радикалы торчат в стороны. Все эти части огромной молекулы как-то размещаются в пространстве относительно друг друга, и в результате белок приобретает свою трехмерную форму — как обычно говорят, конформацию. Белок с нарушенной конформацией, как правило, совершенно бесполезен для организма. Поэтому соблюдение конформации — это очень важно.
Как же она складывается? Есть четыре типа взаимодействий между аминокислотами, определяющих объемную структуру белка, в который они входят.
Во-первых, это водородные связи (см. главу 2). В белке их обычно много, они возникают и между пептидными группами, и между боковыми цепями аминокислот (“боковая цепь” и “радикал” — в данном случае синонимы). Особенно это относится к тем аминокислотам, радикалы которых нейтральны и гидрофильны — вроде, например, серина или тирозина.
Во-вторых, это гидрофобное притяжение между углеводородными радикалами, принадлежащими таким аминокислотам, как валин, лейцин или фенилаланин. Вода выталкивает эти радикалы точно так же, как вытолкнула бы обычные молекулы углеводородов, и они отлично слипаются вместе, если оказываются при сворачивании белковой молекулы близко друг к другу. А тем самым они это сворачивание и закрепляют.
В-третьих, существует электростатическое притяжение между положительно и отрицательно заряженными боковыми цепями. Если, например, глутамат (радикал которого заряжен отрицательно) окажется при сворачивании белка рядом с лизином (радикал которого заряжен положительно), между ними тут же возникнет самая настоящая ионная связь.
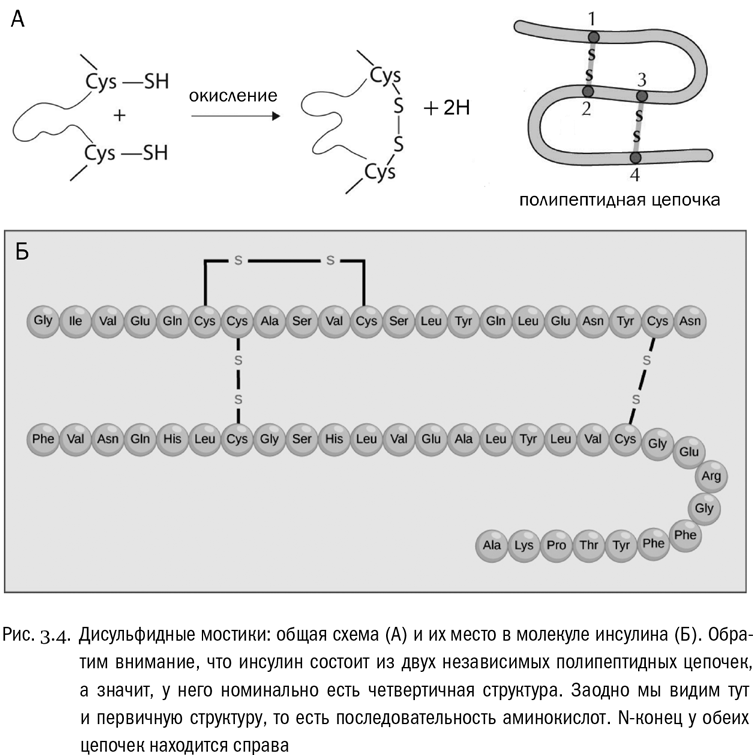
Есть и четвертый тип взаимодействий. Он зависит от единственной аминокислоты, радикалы которой могут образовать между собой аж ковалентные связи (не имеющие никакого отношения к пептидным). Эта аминокислота — цистеин. В радикале цистеина есть сульфгидрильная группа –SH, аналогичная спиртовой группе (–OH), но с атомом серы вместо атома кислорода. Целиком радикал цистеина имеет вид –CH2–SH. Так вот, уже в готовом белке может произойти реакция, при которой у двух таких радикалов будет отобран водород (его унесут специальные молекулы-переносчики), а свободные валентности атомов серы замкнутся друг на друга и образуют между остатками цистеина связь –S–S–. Это называется дисульфидным мостиком (см. рис. 3.4А). Белок вполне может быть в нескольких местах “сшит” такими мостиками (см. рис. 3.4Б). Причем реакция их образования обратима: дисульфидные мостики могут возникать и рваться, и это бывает важно для регуляции некоторых физиологических процессов.
Связи и уровни
Для удобства принято выделять четыре уровня структуры белка. Они так и называются: первичная структура, вторичная, третичная и четвертичная.
Первичная структура — это просто последовательность аминокислот, соединенных пептидными связями (см. рис. 3.4Б). Она всегда линейна, ибо белки не ветвятся. Перечислять аминокислоты в белке принято от N-конца (свободная аминогруппа) к C-концу (свободная карбоксильная группа). Множество таких перечислений, то есть записей первичной структуры белков, есть в современных электронных базах данных, доступных в сети. Можно сказать, что первичная структура белка одномерна, в то время как все остальные уровни — трехмерны. К первичной структуре относятся только пептидные связи, а к остальным уровням — любые другие взаимодействия между аминокислотами, входящими в один и тот же белок.
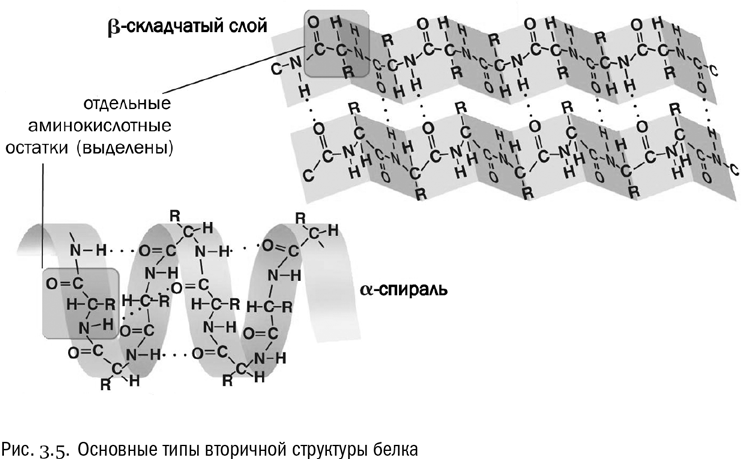
Вторичная структура — это система взаимодействий между аминокислотами в составе одной и той же полипептидной цепочки, расположенными близко (через считаные остатки друг от друга). Вторичная структура держится в основном на водородных связях (см. рис. 3.5). Причем в данном случае это связи между пептидными группами, а не боковыми цепями. А поскольку все пептидные группы одинаковы, то вторичная структура обладает высокой регулярностью, в ней часто повторяется один и тот же “узор”.
Два самых распространенных типа вторичной структуры белка — альфа-спираль и бета-слой. В альфа-спирали водородные связи постоянно образуются между аминокислотными остатками с номерами n и (n+4), то есть каждая аминокислота образует водородную связь с аминокислотой, четвертой по счету от нее. В результате получается компактная спираль, внутри которой находятся пептидные группы, а радикалы торчат в стороны. Альфа-спираль очень устойчива, в том числе и потому, что внутри нее в образовании водородных связей принимают участие все пептидные группы без исключения. В бета-слое полипептидная цепочка несколько раз перегибается, и водородные связи образуются между ее противоположно направленными отрезками.
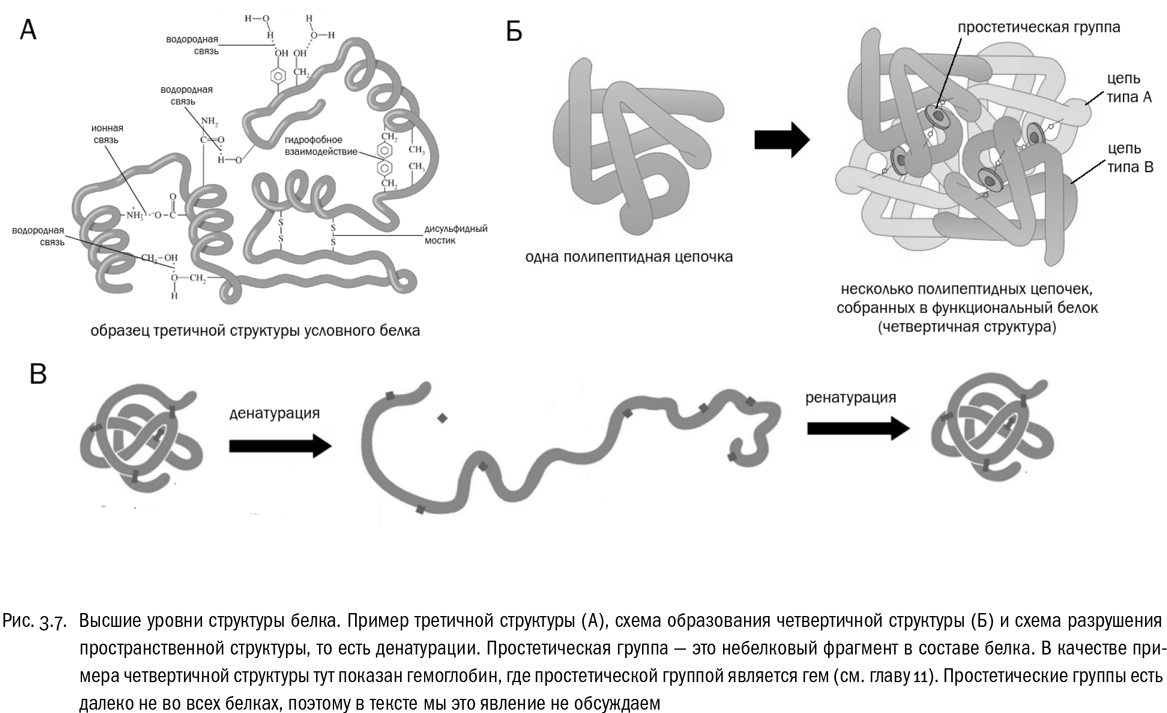
Третичная структура белка — это система взаимодействий между сколь угодно далекими (но принадлежащими к одной и той же полипептидной цепи) остатками аминокислот (см. рис. 3.6, 3.7А). Она определяет, какую форму будет иметь молекула белка целиком. Если вторичная структура — это ближний порядок, то третичная — дальний порядок. В образовании третичной структуры участвуют водородные связи между боковыми цепями, гидрофобные взаимодействия (очень частый случай) и ионные связи между заряженными боковыми цепями. И дисульфидные мостики тоже вносят в третичную структуру свой вклад.
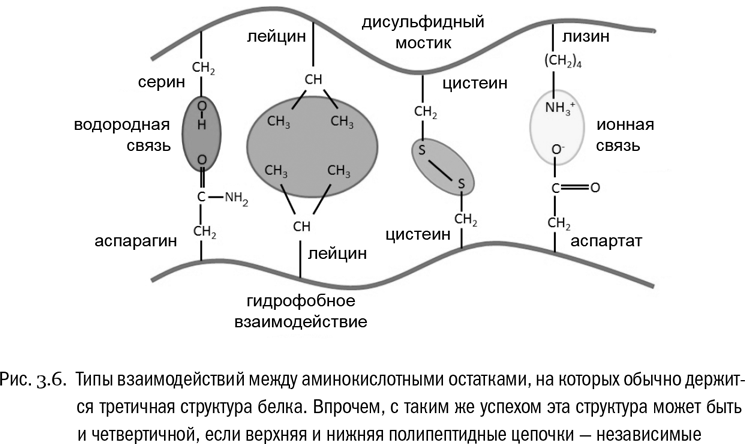
Наконец, четвертичная структура возникает в том случае, если функциональный белок собирается из нескольких отдельных полипептидных цепей (см. рис. 3.7Б). Если белок состоит из одной полипептидной цепи, значит, четвертичной структуры у него нет. Взаимодействия, создающие четвертичную структуру, те же самые, что и в третичной структуре, только не внутри одной полипептидной цепи, а между разными цепями.
Типичный белок с четвертичной структурой — гемоглобин, переносящий кислород в нашей крови. Его молекула состоит из четырех полипептидных цепочек, которые синтезируются отдельно, но свою функцию выполняют только вместе. Объединяются они в основном за счет гидрофобных взаимодействий. Всего молекула нормального гемоглобина взрослого человека включает 574 аминокислоты.
Потеря белком своей пространственной структуры без разрушения пептидных связей (то есть первичной структуры) называется денатурацией, что буквально значит “потеря природы” (см. рис. 3.7В). Самый простой способ денатурировать белок — как следует нагреть его. Именно частичная денатурация белков является основной целью любой тепловой обработки пищи. Причем иногда этот процесс до некоторой степени обратим (при кипячении молока, например). Восстановление пространственной структуры денатурированного белка называется ренатурацией. Но бывает и необратимая денатурация. Например, белок крутого яйца после полной необратимой денатурации растворенных там молекул белков становится твердым, потому что раскрученные полипептидные цепочки перепутываются между собой. Денатурация большинства белков (но не всех!) происходит при температуре 40–50 °С. Это определяет верхний температурный предел для жизни большинства земных живых существ.
Чтобы белок выполнял свою биологическую функцию, нужна, как правило, тончайшая и очень точная “настройка” его пространственной структуры. Нарушения аминокислотной последовательности тем и опасны, что они эту структуру разрушают. Например, существует генетическое нарушение, при котором в строго определенной точке одной из цепей гемоглобина глутамат заменяется на валин. Казалось бы, всего лишь одна аминокислота заменяется на другую. Но здесь это имеет неожиданно серьезные последствия. Глутамат — аминокислота, боковая цепь которой несет отрицательный заряд, валин же нейтрален и гидрофобен. Если рядом окажутся два остатки глутамата, они будут отталкиваться. А если два остатка валина, то, наоборот, слипаться. В данном случае замена глутамата на валин приводит к тому, что слипаться начинают целые молекулы гемоглобина. А это деформирует красные кровяные клетки, в которых он содержится, и вызывает тяжелую болезнь — серповидноклеточную анемию. Именно таков ее молекулярный механизм.
Очевидно, что взаимодействия между аминокислотами в белке неслучайны. Сворачивание белковой молекулы зависит от ее первичной структуры, то есть от того, в каком порядке аминокислоты расположены в цепочке. Иногда говорят, что если бы можно было взять полипептидную цепь за концы, растянуть ее и потом отпустить, то она каждый раз свертывалась бы совершенно одинаково. На самом деле в живой клетке все происходит несколько иначе: там белок синтезируется последовательно, аминокислота за аминокислотой (от N-конца к C-концу), и части молекулы белка, синтезированные раньше, успевают свернуться в трехмерную структуру до того, как будут синтезированы остальные части. Но в итоге все молекулы данного белка сворачиваются строго одинаково. Зная аминокислотную последовательность белка, теоретически можно рассчитать его пространственную структуру всех уровней. Часто это успешно делают и на практике, используя методы таких наук, как биофизика и биоинформатика. В идеале последовательность аминокислот (которую можно записать в строчку, обозначив аминокислоты буквами) должна однозначно определять собой все свойства белка. И более того, изучаемая биологами реальность к этому идеалу очень близка.
На заре жизни... и раньше
Биохимическая эволюция началась еще до образования Земли как планеты. Современные ученые уверены, что синтез веществ, ставших потом биологически активными, шел уже на частицах протопланетного газопылевого облака. Об этом свидетельствует многое — например, химический состав углистых метеоритов, которые потому так и называются, что богаты углеродом (в научной литературе их часто называют углистыми хондритами). Метеориты этого типа никогда не входили в состав планет, поэтому их химический состав не искажен действием высоких температур и давлений, господствующих в планетных недрах. Это своего рода химический “заповедник” очень древней эпохи Солнечной системы — эпохи, когда синтез органических веществ только начинался. Химический анализ углистых метеоритов дал ученым поразительную возможность заглянуть во времена, непосредственно предшествовавшие зарождению жизни.
И ученые не разочаровались. Углистые метеориты оказались удивительно богаты органикой. Например, в них найдены углеводороды, спирты, альдегиды, кетоны, карбоновые кислоты, оксикислоты, амины и углеводы (см. главу 1). Есть там и аминокислоты. Причем очень разные.
В знаменитом Мурчисонском метеорите, упавшем в 1969 году в Австралии, химический анализ обнаружил в общей сложности 52 аминокислоты(см. рис. 3.8). И они необыкновенно разнообразны. Например, среди этих аминокислот есть бета-аланин — изомер аланина, являющийся не альфа-, а бета-аминокислотой (NH2–CH2–CH2–COOH). Есть там гамма- и даже дельта-аминокислоты, например дельта-аминовалериановая кислота (NH2–CH2–CH2–CH2–CH2–COOH). Есть изовалин и метилнорвалин — аминокислоты, где к альфа-углеродному атому присоединены сразу два углеводородных радикала (как мы помним, в “белковых” аминокислотах на месте одного из них всегда атом водорода). Есть циклолейцин, где карбоксильная группа и аминогруппа присоединены к замкнутому пятичленному углеводородному кольцу. Перечислять странные аминокислоты, многие из которых встречаются исключительно в метеоритах, можно еще долго. Есть там, впрочем, и аминокислоты, входящие на Земле в двадцатку “белковых”, — глицин, аланин, валин, лейцин, изолейцин, аспартат, глутамат и пролин.
И вот тут возникает закономерный вопрос: почему одни аминокислоты вошли в состав белков, а другие нет? Кое о чем вполне можно догадаться. Скорее всего, “выбор” аминокислот на роль протеиногенных сильно зависел от формы их молекул. Например, аминокислота с двумя крупными радикалами при альфа-атоме должна ограничивать число возможных конформаций полипептидной цепочки, делая ее менее гибкой, — просто потому, что радикалы будут торчать в разные стороны, мешая изгибам и поворотам. У всех без исключения протеиногенных аминокислот одна из валентностей альфа-атома занята простым водородом. И это, конечно, не случайность.
Примерно так же, скорее всего, объясняется и тот факт, что все белки состоят исключительно из альфа-аминокислот. В результате этого остов полипептидной цепи получается легким, гибким и однородным. Все “тяжелые” группы переводятся в боковые цепи, а все промежутки между пептидными связями имеют строго одинаковую длину. Если бы, например, в состав белка входили вперемешку альфа- и бета-аминокислоты, последнее условие не соблюдалось бы и это сделало бы невозможными регулярные конформации — такие как широко распространенная в реально существующих белках альфа-спираль.
А теперь отвлечемся от белков и подумаем про общие принципы развития Вселенной. Благо повод для этого сейчас есть. Не раз упоминавшийся тут Станислав Лем писал, что творческий потенциал неживой природы больше, чем у живой, по одной простой причине: она не ограничена требованиями естественного отбора. “Лишь там, где царит смерть, вечная, спокойная, где не действуют ни сита, ни жернова естественного отбора, формирующие любое создание по законам бытия, открывается простор для удивительных произведений материи, которая, ничему не подражая, никому не подчиняясь, выходит за границы человеческого воображения”, — пишет Лем в замечательном романе “Фиаско”. И добавляет: на планетах, освоенных жизнью, химические соединения попали “в рабство к биологической эволюции”. Они усложнились, но потеряли свое огромное исходное разнообразие.
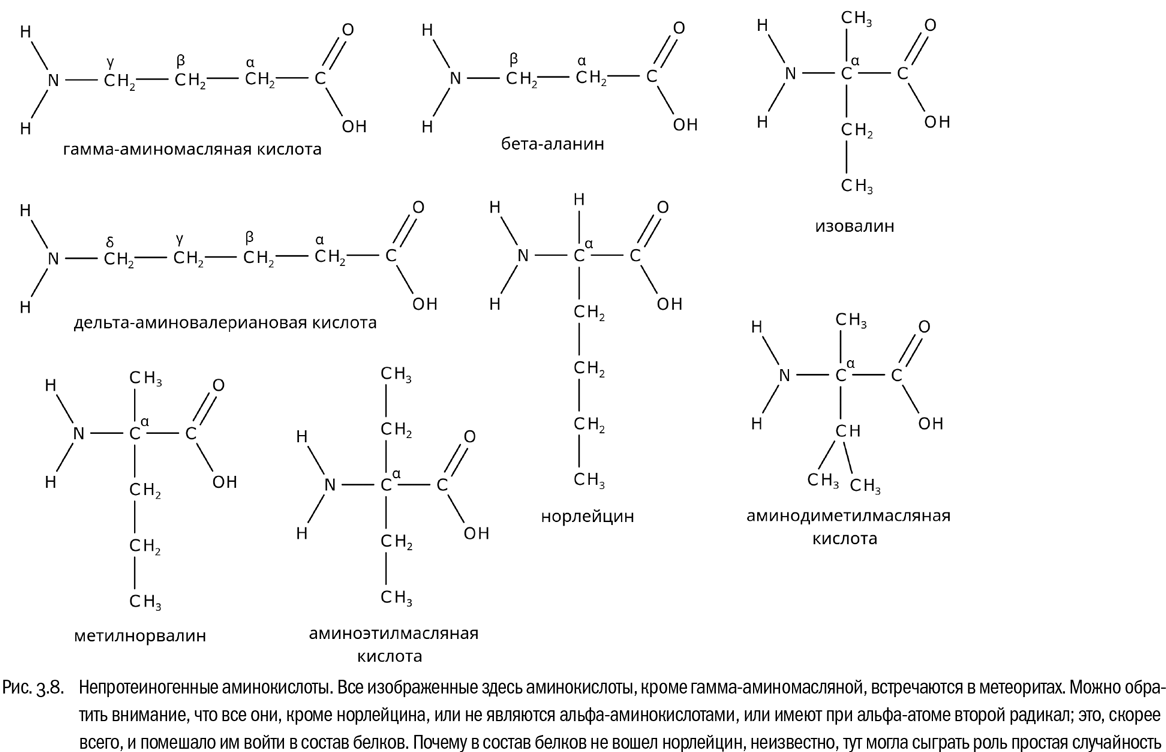
То, что мы знаем об аминокислотах Солнечной системы, превосходно иллюстрирует эту мысль. Пока никакой жизни не существовало, самые разные аминокислоты синтезировались более-менее одинаково легко. Их было много десятков. После возникновения жизни ситуация резко изменилась: аминокислоты, вошедшие в состав белков, стали активно синтезироваться живыми организмами, и на Земле (о жизни в других местах мы пока ничего не знаем) их концентрации колоссально выросли. Аминокислоты, не прошедшие отбора на протеиногенность, наоборот, стали редкими. Некоторые из них, например изовалин, не встречаются на Земле вообще никогда. Между прочим, именно открытие в Мурчисонском метеорите изовалина стало самым сильным доводом в пользу того, что аминокислоты не были занесены в этот метеорит земными микробами после его падения на Землю, а присутствовали там изначально. Изовалина на Земле просто нет, так что никакие микробы занести его не могли. Он синтезировался в космосе совершенно самостоятельно. Но поскольку у изовалина два углеводородных радикала, то на роль составной части белков он не подошел. Со многими другими аминокислотами произошло то же самое. И это, конечно, не случайность. Уже знакомая нам общая формула протеиногенной аминокислоты — это не изолированный факт, который можно только зазубрить, а вполне осмысленный продукт эволюции.
Функции белков
Любой отдельный белок — тоже продукт биологической эволюции. Его аминокислотная последовательность, как и вся структура, всегда приспособлена под какую-нибудь строго определенную функцию. Известный биофизик Лев Александрович Блюменфельд писал: “Если бы для описания клетки нам пришлось выбирать между двумя крайними моделями — часовым механизмом и гомогенной химической реакцией в газовой фазе, — выбор был бы однозначен: клетка несравненно ближе к часовому механизму, чем к чисто статистической системе”. Можно добавить, что это относится не только к целой клетке, но и к отдельным макромолекулам, то есть в первую очередь к белкам. Блюменфельд как раз и начинает вышеприведенными словами главу своей книги, посвященную биофизике молекул белка.
Функционирующий белок можно в самом что ни на есть буквальном смысле рассматривать как молекулярную машину, то есть как машину размером с молекулу. По определению Блюменфельда, машина — это конструкция с выделенными внутренними степенями свободы (то есть, попросту говоря, с подвижными частями), использующая собственное механическое движение для передачи силы от одной части системы к другой. Белковая молекула этому определению, безусловно, соответствует. В ней хватает внутренних степеней свободы (множество ковалентных связей, вокруг которых возможны повороты), и она вполне может передавать силу с помощью своих подвижных частей. И многие белки — например, мышечные — постоянно используют это, совершая настоящую механическую работу. Но это далеко не единственное, что белки могут делать.
А что же, собственно, они делают? Самый близкий к истине ответ — да все! Или, во всяком случае, почти все. Функции белков настолько многообразны, что никакое их перечисление, скорее всего, не будет абсолютно полным. Но мы все-таки попробуем назвать главные функции белков, помня про эту оговорку.
* Структурная функция относится к белкам, из которых сделаны те или иные части живых тел. Например, коллаген — белок, образующий механическую основу костей, хрящей и соединительнотканного слоя кожи. Кератин — белок, из которого состоят волосы, ногти и наружный роговой слой кожи. Кристаллины — белки, из которых в основном состоит хрусталик глаза. И так далее.* Каталитическая функция, связанная с ускорением химических реакций. О ней — чуть ниже.* Сигнальная функция: белки, предназначенные для передачи информации. Эта функция на редкость многолика. Бывают белки-нейротрансмиттеры, передающие сигналы между нервными клетками. Бывают белки-гормоны, передающие сигналы примерно тем же способом, но через кровь, по всему организму сразу. Бывают белки-рецепторы, которые, наоборот, принимают сигналы, сидя на поверхности клетки. Бывают белки-посредники, обеспечивающие проведение сигнала уже внутри клетки. И это далеко не все возможности, но вникать в детали мы сейчас не будем.* Транспортная функция. Например, известный всем гемоглобин — это белок, переносящий молекулы кислорода из одной части организма в другую.* Двигательная функция свойственна белкам, от которых зависит сокращение мышечных клеток животных, но не только им. Например, двигательные структуры одноклеточных организмов — жгутики, реснички, ложноножки — тоже обязательно содержат специальные моторные белки.* Защитная функция. Это всевозможные яды, а также антитела, то есть белки, выделяемые клетками иммунной системы и убивающие опасных “гостей” организма (например, попавших туда бактерий).
Эти функции, пожалуй, главные. Ясно, что живым существам нужны они все. И тем не менее среди них можно выделить одну совершенно особую функцию, настолько распространенную и важную, что без нее белки как природное явление вообще невозможно представить. Эта функция — каталитическая. Вот о ней стоит поговорить подробнее.
Ферменты
Начнем с простых определений. Вещество, ускоряющее химическую реакцию, но само не претерпевающее в ней стойких изменений, называется катализатором. А катализатор, являющийся белком, называется ферментом. Ускорять он может все что угодно. Все биохимические реакции идут не сами по себе, а с помощью ферментов. Например, даже такой предельно простой процесс, как слияние углекислоты (CO2) и воды (H2O) в молекулу угольной кислоты (H2CO3), все равно катализируется специальным ферментом — карбоангидразой, которая ускоряет его примерно в миллион раз. А для более сложных реакций ферменты тем более необходимы. Можно без особого преувеличения сказать, что ферменты контролируют в живом организме вообще все.
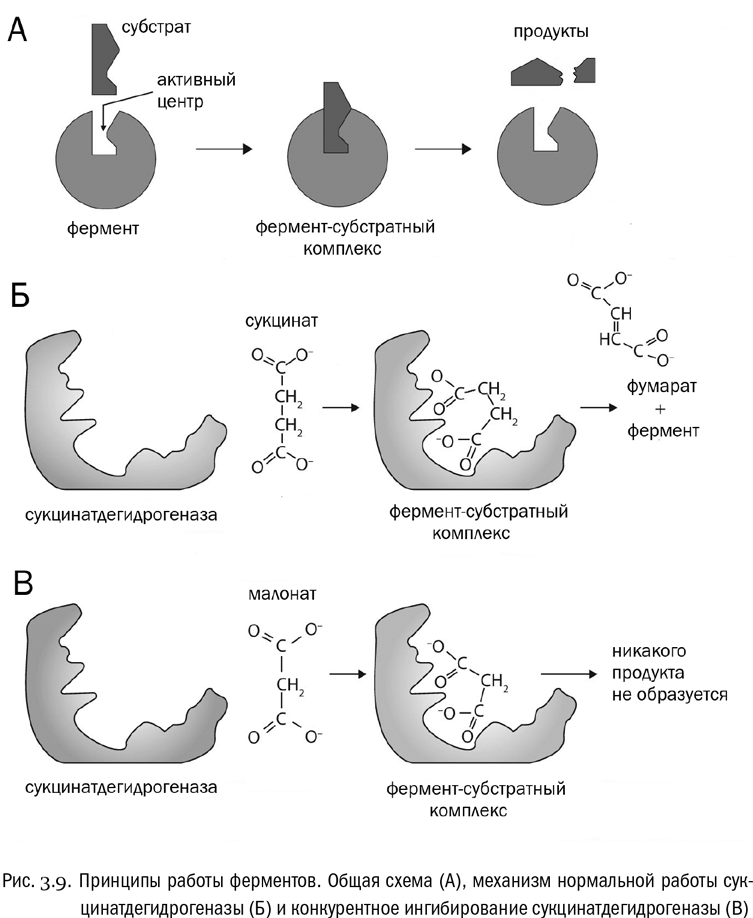
Вещество, являющееся исходным для той реакции, которую катализирует данный фермент, называется его субстратом. Молекула фермента должна войти в контакт с молекулой субстрата и подвергнуть ее некоему действию — например, расщепить надвое, или поменять в ней местами функциональные группы, или сшить что-нибудь ковалентной связью, или разорвать эту связь, — вариантов тут множество. Но в любом случае молекула фермента должна сначала захватить молекулу субстрата, а потом преобразовать ее и высвободить. Часть молекулы фермента, непосредственно контактирующая с молекулой субстрата, называется активным центром. Ферменты — это обычно довольно крупные белки, а в активном центре может быть всего-навсего несколько аминокислот. Поэтому, как правило, активный центр занимает только небольшую часть молекулы фермента (см. рис. 3.9А).
Если говорить совсем примитивно, активный центр — это такое гнездо в молекуле фермента, куда молекула субстрата должна войти, как ключ в замок. Как только она туда попадет, молекула фермента ее захватит и преобразует. Очевидно, что для этого конформация активного центра должна очень точно совпадать с очертаниями молекулы субстрата — в самом деле как замочная скважина с ключом. Это прямо так и называют “моделью ключа и замка”. Правда, на самом деле активный центр фермента, в отличие от механизма замочной скважины, является скорее гибким, чем жестким. При взаимодействии с субстратом его конформация всегда меняется — примерно так, как меняется форма перчатки, когда ее надевают на руку. Модель работы ферментов, учитывающая это, называется “моделью индуцированного соответствия”. Когда реакция завершается, конформация фермента возвращается к прежней.
Биофизики уверены, что во всех этих процессах молекула фермента действует как сложная механическая машина, имеющая множество шарниров, сочленений, поворачивающихся частей и т.п. И это, конечно, впечатляет. “Самонадеянно скажет иной: “Сколочу-ка телегу!” // Но ведь в телеге-то сотня частей! Иль не знает он, дурень?” — писал в поэме “Труды и дни” великий древнегреческий поэт Гесиод. А ведь молекула любого фермента (пусть даже и небольшого) устроена намного сложнее гесиодовой телеги. Причем это будет верно, даже если мы станем рассматривать ее исключительно как механическую машину, игнорируя всю тонкую структуру атомного уровня.
Номенклатура ферментов довольно сложна, потому что их очень много. Но в большинстве случаев название фермента включает, во-первых, название его субстрата и, во-вторых, характерное окончание “-аза”. Если мы видим где-то слово с таким окончанием, это наверняка название какого-нибудь фермента.
У термина “фермент” есть синоним. Это слово “энзим”. Например, в английском языке ферменты называют почти исключительно энзимами (enzyme), и в русские переводы с английского это тоже иногда проникает. Но все-таки по-русски гораздо чаще говорят именно “фермент”. На всякий случай запомним, что “фермент” и “энзим” — синонимы, и употребление одного из этих слов вместо другого большой ошибкой не будет.
Очень важное свойство ферментов — специфичность. Это значит, что каждый фермент приспособлен к одной строго определенной химической реакции. Субстрат должен точно подойти к его активному центру, иначе реакция не пойдет.
Например, фермент сукцинатдегидрогеназа захватывает молекулу янтарной кислоты (HOOC–CH2–CH2–COOH) и превращает ее в молекулу фумаровой кислоты (HOOC–CH=CH–COOH). Янтарная кислота при этом, как видим, теряет два атома водорода. Водород по-латыни “гидроген”, а “дегидрогеназа” — фермент, его отнимающий. Субстратом же этого фермента является янтарная кислота. В растворе она диссоциирует, и от нее остается анион, который называется сукцинатом. Таким образом, сукцинатдегидрогеназа — это фермент, отнимающий водород у сукцината (см. рис. 3.9Б).
Однако возможна ситуация, когда на месте сукцината окажется малонат — анион малоновой кислоты, отличающейся от янтарной на один атом углерода (HOOC–CH2–COOH). Молекула малоната приблизительно подходит по форме к активному центру сукцинатдегидрогеназы и может его занять. Но поскольку соответствие все-таки неточное, никакой реакции в этом случае не произойдет (см. рис. 3.9В). Активный центр сукцинатдегидрогеназы будет просто заблокирован. Это называется конкурентным ингибированием. Конкурентное ингибирование — очень эффективный механизм “выключения” ферментов, на нем основано действие многих лекарств и ядов.
В активных центрах ферментов бывает очень полезным уже знакомое нам разнообразие аминокислот. Чтобы молекула субстрата встала на свое место и повела себя как надо, очертания активного центра должны быть под этот субстрат идеально подогнаны. Для этого где-то можно разместить гидрофобные карманы, образованные боковыми цепями аланина или валина, где-то — отрицательные заряды аспартата или глутамата, где-то — нейтральные гидрофильные аминокислоты, которые образуют с субстратом водородные связи, и так далее. Если, например, в субстрате есть положительный заряд, то в этом-то месте как раз и можно выставить отрицательно заряженный радикал какого-нибудь аспартата, чтобы электростатическим притяжением удержать молекулу субстрата в нужном положении. Таких примеров можно привести множество.
Интересно, что аминокислоты, оказывающиеся рядом в активном центре, запросто могут в первичной последовательности находиться очень далеко — например, за 300 остатков друг от друга. Их “правильное” взаимное расположение достигается за счет очень точного объемного сворачивания полипептидной цепи, то есть за счет третичной структуры. Можно представить, насколько сложной биохимической машиной является такой фермент! А между тем в типичной живой клетке ферментов несколько тысяч.
Любой фермент работает только в достаточно строго определенном диапазоне внешних условий — например, таких, как температура и кислотность. Если температура слишком высока, фермент может просто денатурировать, то есть потерять вторичную и третичную структуру. Понятное дело, что его активный центр при этом развалится. Если же слишком высока кислотность, это может повлиять на поведение радикалов некоторых аминокислот. Мы знаем, что кислотность — это концентрация протонов (H+). Если в растворе будет слишком много протонов, боковые цепи отрицательно заряженных аминокислот (аспартата и глутамата) волей-неволей присоединят их. Например, радикал глутамата, обычно имеющий вид –CH2–CH2–COO–, в этом случае перейдет в состояние –CH2–CH2–COOH. Отрицательный заряд в нем исчезнет, и он больше не сможет выполнять свою функцию в активном центре. Поэтому слишком высокая кислотность вредна. Живым организмам очень важно поддерживать постоянство своей внутренней среды не в последнюю очередь потому, что иначе работа ферментов разладится.
“Белковая Вселенная”
Представим себе дипептид, в котором всего-навсего две аминокислоты. На первой позиции в этом дипептиде может быть любая из 20 стандартных протеиногенных аминокислот. Проведем ось, на которой каждая аминокислота будет отмечена черточкой. Это — 20 возможных вариантов.
На второй позиции в дипептиде тоже может быть любая из 20 стандартных протеиногенных аминокислот. Проведем вторую ось, перпендикулярную первой, и тоже отметим на ней черточкой каждую аминокислоту. У нас получится плоский график. Между двумя осями образуется пространство, состоящее из 400 точек (202). Каждая точка соответствует одному возможному дипептиду. А всего таких дипептидов может быть ровно 400.
Поздравим себя: мы только что познакомились с важнейшей концепцией, которая называется “белковая Вселенная”. Это воображаемое пространство, в каждой точке которого находится один белок. “Белковая Вселенная” — не поэтическая метафора, а на сегодняшний день уже повседневный инструмент работы биоинформатиков, примерно как геометрическое пространство декартовых координат для обычного инженера.
“Белковая Вселенная”, состоящая всего из 400 точек, — очень маленькая и простая. Но стоит добавить в наш пептид хотя бы третью аминокислоту, как пространство превратится в трехмерное и точек в нем станет уже 8000 (203). Дальше можно добавить и четвертую аминокислоту, и пятую — и так, пока не дойдет до обычных белков, число аминокислот в которых измеряется как минимум десятками, а обычно сотнями. Число измерений пространства “белковой Вселенной” будет в каждый момент равно числу аминокислотных остатков, сколько бы их ни было. Это пространство будет многомерным. Наглядно представить себе многомерное пространство человек не может, но это от него и не требуется, достаточно того, что модель в принципе годится для расчетов.
Важно понимать, что с ростом длины белка объем “белковой Вселенной” растет чрезвычайно быстро. Невообразимо быстро. Для белка из 300 аминокислот (а это типичный размер) “Вселенная” является 300-мерной и включает 20300 возможных аминокислотных последовательностей. Это гораздо больше, чем, например, общее число протонов и нейтронов в наблюдаемой части физической Вселенной (1080). Совершенно очевидно, что до сих пор биологическая эволюция исчерпала лишь ничтожную часть этого объема. Подавляющее большинство белков, последовательности которых существуют где-то в многомерном идеальном пространстве, еще ни разу не были воплощены, то есть физически синтезированы.
Более того, показано, что “белковая Вселенная” (вернее, ее реально освоенная часть) расширяется. С ходом эволюции белки разных организмов отличаются друг от друга все сильнее и сильнее. В этом “белковая Вселенная” подобна нашей обычной Вселенной, которая тоже расширяется, — в свое время это математически показал Александр Фридман и (независимо от Фридмана) вывел из астрономических наблюдений Эдвин Хаббл. Проводя свои наблюдения, Хаббл обнаружил, что галактики удаляются друг от друга, причем скорость их разлетания тем выше, чем больше между ними расстояние. Мысленно продолжив эту тенденцию в далекое прошлое, Хаббл пришел к выводу, что движение галактик должно было начаться из одной точки. Это послужило прекрасным подтверждением теории Большого взрыва, которая как раз в те годы начинала создаваться. Ну, а теперь мы понимаем, что примерно так же устроена и “белковая Вселенная”, в которой идет биологическая эволюция. Белки всех живых организмов стремительно (в эволюционном масштабе времени) “разбегаются” в своем многомерном пространстве от исходной точки — ограниченного набора белков единственного общего предка.
Рассматривая карту окружающей нас части физической Вселенной, можно подобрать такой масштаб, в котором не только галактика Млечный Путь, но и весь суперкластер Девы (куда входит Млечный Путь вместе с еще примерно 30 000 галактик) будет выглядеть как ничтожно малые точки. Так вот, соотношение освоенных и неосвоенных областей “белковой Вселенной” должно выглядеть как-то очень похоже. Белки всех живых организмов, вместе взятых, возникшие за несколько миллиардов лет истории Земли, до сих пор занимают в “белковой Вселенной” лишь ничтожно малую область. В этом смысле эволюция только началась! А ведь могут быть еще и другие, небелковые биологические Вселенные.
Назад: 2. вода
Дальше: 4. cимметрия

