Книга: Динозавры. 150 000 000 лет господства на Земле
Назад: Таз и задние конечности
Дальше: Кость динозавра под микроскопом
Функционирование скелета динозавра
Кости дают нам массу информации о строении тела животного, а также о том, как оно жило, как двигалось и на какие виды поведения было способно. Сочлененные скелеты не только показывают, как кости соединялись между собой при жизни, но и диапазон их возможных движений и какие позы мог принимать динозавр. Мы уже знаем, что отрасль биологии, которая изучает, как работают разные системы животного по его скелету, называется функциональной морфологией.
Важно помнить, что анатомия – это не только кости. На движения костей влияют хрящевые прокладки между ними, мышцы и веревки-сухожилия, которые соединяют кости. К большому разочарованию палеонтологов, почти ничего из этого не сохраняется в окаменелостях. Все они успевают разложиться до начала фоссилизации, потому их и называют «мягкими тканями», в отличие от «твердых тканей» – костей и зубов. По крайней мере мы можем получить некоторое представление о возможностях вымерших животных, сравнивая их скелеты со скелетами современных животных. Разработаны многочисленные методы, которые позволяют учитывать недостающие мягкие ткани и определять их влияние на передвижение и гибкость животных.
Самый простой и «традиционный» способ изучения функциональной морфологии у вымерших животных – двигать кости в суставах. Иногда для этого их моделируют на бумаге или при помощи математических расчетов. Это не очень точный метод, к тому же он обычно не учитывает влияние мягких тканей, о которых мы упоминали. Для него также нужен прямой доступ к самим окаменелостям, а они могут быть хрупкими или поврежденными. Благодаря технологическому прогрессу способы изучения функциональной морфологии сильно продвинулись. Регулярно используются цифровые модели скелетов, сделанные с помощью фотографий или сканеров, которые позволяют оцифровать форму костей. Пользоваться компьютерными моделями для изучения движения костей намного проще, чем работать с полноразмерными костями или их копиями. Порой результаты цифрового моделирования очень интересны и подтверждают некоторые противоречивые идеи о поведении динозавров. Давайте рассмотрим области анатомии динозавров, которые изучались с использованием как традиционных, так и современных методов.

Сегодня при изучении функциональной морфологии используются компьютерные модели и томография. В прошлом ученым часто приходилось манипулировать самими окаменелостями – или моделями окаменелостей, – чтобы увидеть, как животные могли бы двигаться при жизни. На фото: ученые изучают слепок черепа аллозавра
На протяжении десятилетий специалисты по динозаврам изучали челюсти гадрозавров и других игуанодонтов, чтобы понять, как эти животные жевали. Было предложено много идей, большинство из которых не объясняли, как мог возникнуть характерный рисунок износа зубов. В 1980-х гг. палеонтологи Дэвид Норман и Дэвид Вейшампел предложили новую идею. Они заметили, что через всю морду игуанодона, мантеллизавра и гадрозавров проходит зона, которая могла быть гибкой. Основываясь на том, как кости соединяются вокруг этой зоны, они предположили, что верхнечелюстная кость – крупная кость, которая образует большую часть морды и на которой расположены зубы верхней челюсти – могла смещаться внутрь и наружу, благодаря чему щечные зубы терлись друг о друга. Подвижные области в других частях черепа вроде бы тоже позволяли совершать такое движение. Согласно этой интерпретации, игуанодонты обладали черепным кинетизмом (мы уже встречались с этим термином). Это уникальное устройство черепа, позволяющее зубам тереться друг о друга, отсутствует у любого другого животного, современного или вымершего.
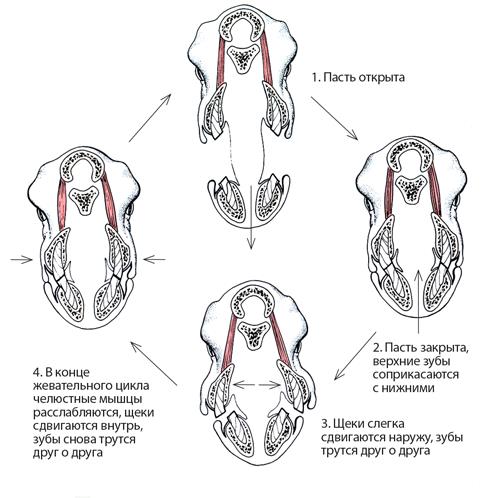
Согласно исследованиям Дэвида Нормана и модели плеврокинеза Дэвида Вейшампела, орнитоподы, например эдмонтозавр, обладали уникальным способом трения верхних зубов о нижние. На схеме показана последовательность движений челюстей, голова животного изображена в поперечном сечении
Вдохновленные этой гипотезой, палеонтологи стали предполагать, что у многих других нептичьих динозавров тоже были подвижные черепа. Высказывались идеи, что этим свойством обладали аллозавриды, тираннозавриды, альваресзавриды и др. С этим согласны не все. В 2008 г. анатомы Кейси Холлидей и Ларри Уитмер заявили, что на черепах всех нептичьих динозавров есть сцепленные друг с другом отростки, из-за которых кинетизм невозможен. К тому же в черепах динозавров не было подвижных, заполненных жидкостью суставов, которые есть у современных животных с черепным кинетизмом, таких как птицы и ящерицы. Если так, то почему у игуанодона и других динозавров есть признаки, допускающие по крайней мере некоторую степень подвижности черепа? Холлидей и Уитмер предположили, что это на самом деле были области роста костей, а не зоны гибкости.
Было немало споров и о расположении плечевого пояса у завропод и других четвероногих динозавров, например цератопсов. Здесь дискуссии ведутся насчет положения передних конечностей: были ли они расставлены в стороны, как у ящериц и черепах, или располагались под телом, как опорные колонны, на манер крупных млекопитающих вроде носорогов? Темы положения плечевого пояса и постановки передних конечностей взаимосвязаны, так как от положения плечевого пояса зависит расположение плечевого сустава. А оно, в свою очередь, влияет на положение передних конечностей.
Разные эксперты высказывали разные мнения, но сочлененные скелеты показывают, что плечевые пояса четвероногих динозавров располагались низко, причем настолько, что коракоиды почти соприкасались. Если принять это за реальное положение плечевого пояса, это значит, у цератопсов и других птицетазовых передние конечности были ориентированы вертикально, как колонны, а плечевые суставы были направлены назад, слегка наружу и вниз. Следовательно, их передние конечности не были расставлены, как у ящериц, но и не находились непосредственно под телом. Такое положение передних конечностей соответствует найденным следам, что говорит в его пользу.
Еще один спорный аспект анатомии динозавров, который тоже изучали с использованием и традиционных, и компьютерных методов, – это шеи завропод. Долгие годы палеонтологи не могли определиться, как завроподы держали свою шею, как ее использовали и насколько гибкой она была. Учитывая экстраординарную длину шеи, кажется логичным, что она была нужна завроподам, чтобы дотягиваться до верха деревьев и поедать пищу, недоступную другим растительноядным. Но подтверждают ли наши знания эту гипотезу?
Сопоставив шейные позвонки британского завропода цетиозавра, палеонтолог Джон Мартин заключил, что его шея, скорее всего, действовала как балка, которую динозавр держал прямо перед телом и не мог поднять выше уровня спины. Некоторые эксперты распространили эту гипотезу почти на всех завропод, в том числе брахиозавров и мамэньсизаврид с исключительно длинной шеей. Те же эксперты утверждают, что подвижность между смежными шейными позвонками была совсем небольшой и что завроподы в основном питались на уровне земли или плеч.
Для более тщательного изучения гибкости шеи программист Кент Стивенс и палеонтолог Майк Пэрриш построили цифровые модели апатозавра и диплодока. Было принято, что диапазон движения шеи этих цифровых динозавров такой же, как у современных животных. Стивенс и Пэрриш в основном пришли к тем же выводам, что и Мартин: шея действовала как горизонтальная балка, лучше изгибалась по бокам и вниз, чем вверх, и была малоподвижной у основания.
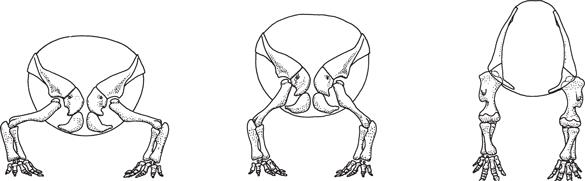
Эксперты выдвигали разные идеи о том, как были устроены скапулокоракоиды и передние конечности четвероногих птицетазовых. Две крайние – что они либо были полностью расставлены, либо полностью выпрямлены, как показано на схемах. Истина, скорее всего, где-то посередине
Вроде бы получается, что компьютерное моделирование подтверждает результаты, полученные при помощи традиционной работы с костями. Но гипотеза, что завроподы могли держать шею только прямо, в основном базируется на допущении, что суставы между шейными позвонками очень малоподвижны. Если же вспомнить, что нам известно о хряще в шейных суставах современных животных, то нужно предположить, что суставы между шейными позвонками завропод были способны к широкому диапазону движений – достаточному, чтобы сгибать шею далеко в сторону или вверх и вниз. Все дело в том, что компьютерное моделирование и работа с костями иногда могут давать неточные результаты, потому что они не учитывают мягкие ткани, которые обычно не сохраняются в окаменелостях, но учитывать их все равно нужно. Наличие мягких тканей означает, что животные в жизни могли выполнять действия, которые кажутся маловероятными, если использовать доказательства, представленные одними костями.

Если учесть хрящевые диски между шейными позвонками завроподовых динозавров, то окажется, что шея была очень гибкой и ее можно было сильно сгибать вверх (и вниз). Наверху показана шея апатозавра, внизу – диплодока
Другие исследователи тоже проводили компьютерное моделирование скелетов динозавров. Немецкий палеонтолог Хайнрих Маллисон разработал и изучил цифровые модели передних конечностей завроподоморфов, хвостов стегозавров и других частей тела динозавров. Его работа о передних лапах платеозавра позволила реконструировать диапазон их движения. Пальцы можно было довольно сильно сжать, лапы были зафиксированы в положении «ладонями внутрь», а запястье позволяло сгибать ладонь внутрь, но не вращать. Такая конфигурация должна показаться знакомой, ведь мы уже видели ее у теропод.
Построенная Маллисоном цифровая модель шипастого африканского кентрозавра показала, что хвост был достаточно гибким, чтобы им можно было размахивать в стороны и вверх. Такая подвижность согласуется с гипотезой, что шипы на хвосте стегозавров использовались как оружие и что стегозавры могли размахивать хвостом с большим диапазоном, защищаясь от крупных теропод. Компьютерные исследования, которые должным образом учитывают мягкие ткани между позвонками, показывают, что шеи и хвосты нептичьих динозавров часто были довольно гибкими.
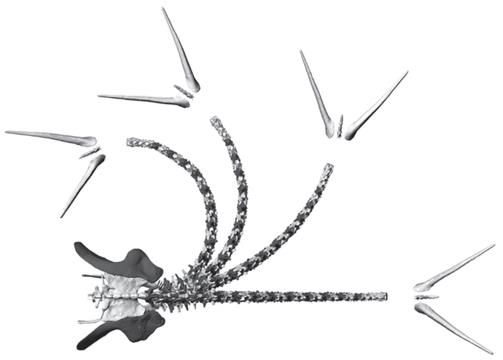
Определив диапазон движения между хвоствыми позвонками африканского стегозавра кентрозавра, ученые показали, что его хвост мог сгибаться очень сильно
Мы все еще находимся на ранних этапах изучения функциональной морфологии динозавров, и предстоит еще немало работы. Что особенно любопытно, повышенный интерес к функциональной морфологии нептичьих динозавров вдохновил многих ученых на изучение анатомии и функциональной морфологии нынешних животных, поскольку анатомия многих из них, как ни странно, никогда не изучалась подробно. Как видно из приведенных здесь примеров – черепного кинетизма, гибкости шеи завропод и т.д., – развивающиеся методы помогают палеонтологам изучать многие разделы анатомии динозавров и их функциональную морфологию гораздо глубже и подробнее, чем было возможно раньше.
Назад: Таз и задние конечности
Дальше: Кость динозавра под микроскопом

