Книга: Один день из жизни мозга. Нейробиология сознания от рассвета до заката
Назад: Нейронные ансамбли, пространство и время
Дальше: Метаансамбли?
Внутри ансамблей
Принято считать, что наименьшая функциональная структура для передачи информации в мозге – это синапс. Поэтому кажется естественным предположение, что динамика нейронного ансамбля будет определяться главным образом процессом синаптической передачи. Но если допустить, что ансамбль – лишь конгломерат синапсов, то общая картина была бы совсем иной, нежели та, что мы видим в реальности.
Границы ансамбля (см. рис. 2) простираются на расстояния, в несколько раз превышающие прогнозируемые согласно классическим представлениям о синаптической передаче. Что касается времени, мы знаем, что для затухания активности в пределах ансамбля до 20 % от максимального уровня, требуется около 300 миллисекунд и что потребуется еще больше времени, чтобы активность полностью исчезла. Но помимо этого мы знаем, что временные рамки синаптической активности не превышают 20 мс.
Наглядно иллюстрирует эти временные рамки рис. 7. Обратите внимание, что хотя передача сигнала от таламуса к коре путем классической синаптической передачи занимает всего пять миллисекунд на каждые два миллиметра, требуется еще двадцать миллисекунд, чтобы ансамбль распространился внутри коры. Здесь вступают в игру два особых способа общения между клетками мозга: один имеет преимущество в виде большего масштаба (объемная передача), в то время как другой гораздо более минималистичный (щелевые контакты). Давайте кратко рассмотрим каждый из них.
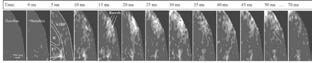
Рис. 7. Нейронные ансамбли в срезе крысиного мозга, визуализированные с помощью потенциал-чувствительных красителей (Fermani, Badin&Greenfield). В то время как нормальная синаптическая передача занимает 5 мс на 2 мм от таламуса до коры, последующее формирование ансамбля занимает в четыре раза больше времени (для достижения радиуса 0, 5 мм)
Объемная передача обеспечивает менее специфичное и значительно более медленное взаимодействие между нейронами, но ее преимущество состоит в том, что она затрагивает гораздо больше клеток. За последние тридцать лет этот прежде революционный механизм был тщательно исследован и теперь рассматривается как альтернатива классической синаптической передачи.
Фактически с 1970-х годов известно, что классические передатчики, такие как дофамин, могут выделяться из участков нейрона, напоминающих ветви дерева (дендриты). Обычно дендриты выступают в роли акцепторной зоны нейрона – мишени для сигналов от других нейронов. Однако теперь известно, что дендриты могут выделять вещества сами по себе. Более того, высвобождение веществ из дендритов происходит достаточно интенсивно, и это дает нам основания полагать, что это принципиально новый эффективный способ модуляции.
Третий процесс не связан с непосредственной передачей электрических сигналов через «щелевые контакты» – прямые контакты между нервными клетками. Интересно, что в нейронных сетях очень быстрые колебания активности (200 Гц) возникают именно посредством распространения электрических сигналов. И это широкое распространение активности осуществляется в том же масштабе, который характерен для ансамблей. Таким образом, ансамбли удовлетворяют пространственно-временным требованиям к сознанию, поскольку, в отличие от локализованных нейронных структур, они не являются ни устойчивыми во времени, ни пространственно ограниченными.
Эти три процесса, определяющие формирование, существование и затухание ансамбля, по всей вероятности, действуют согласованно. Но главный вопрос остается нерешенным: что происходит потом, что приводит к возникновению момента сознания? Ключ может скрываться во времени…
Назад: Нейронные ансамбли, пространство и время
Дальше: Метаансамбли?

