Книга: Вопрос жизни. Энергия, эволюция и происхождение сложности
Назад: Глава 5 Появление сложных клеток
Дальше: Часть IV Предсказания
Глава 6
Половое размножение и происхождение смерти
Природа не терпит пустоты, писал Аристотель. О том же две тысячи лет спустя рассуждал Исаак Ньютон. Оба ученых очень интересовались, чем же заполнено пространство. Ньютон полагал, что пространство заполнено загадочной субстанцией – эфиром. Физики в XX веке отказались от эфира, но в экологии идея horror vacui, “боязни пустоты”, жива и здравствует. Мысль, что все экологические ниши должны быть заняты, выражена в стихотворении Джонатана Свифта:
Под микроскопом он открыл, что на блохе
Живет блоху кусающая блошка;
На блошке той блошинка-крошка,
В блошинку же вонзает зуб сердито
Блошиночка, и так ad infinitum.
Любая экологическая ниша, которую только можно вообразить, оказывается занята видом, который наилучшим образом приспособлен к ней. Каждый организм (растение, животное, бактерия и т. д.) является пристанищем для паразитов, вирусов и мобильных генетических элементов всех мастей, не говоря уже о хищниках крупнее себя. И так всегда.
Но, оказывается, это видимость. Покрывало жизни кажется сплошным, но на самом деле в нем зияет прореха. Пришло время обратиться к главной загадке биологии: почему все организмы делятся на морфологически простых прокариот и эукариот с набором замысловатых черт, не встречающихся у прокариот. Их разделяет пропасть, пустота, которой природа действительно не терпит. С точки зрения морфологии у эукариот есть все, а у прокариот – ничего. Прекрасная иллюстрация самой несправедливой догмы: “Ибо кто имеет, тому дано будет, а кто не имеет, у того отнимется и то, что имеет”.
Мы видели, как эндосимбиотические отношения двух прокариот помогли им выбраться из тупика морфологической примитивности. Хотя одной бактерии непросто проникнуть в другую, некоторые примеры нам известны, и мы точно знаем, что эндосимбиоз прокариот возможен, пусть и очень редок. Но одна клетка, живущая внутри другой – это лишь начало, момент зачатия, не более того. Лишь клетка в клетке. От этого состояния каким-то образом нужно прийти к настоящей сложности: к клетке со всеми присущими эукариотам признаками. Начав с бактерий, у которых почти нет сложных морфологических черт, мы должны закончить эукариотами: клетками с ядром, множеством внутренних мембран и компартментов, динамическим цитоскелетом и сложным поведением – например сексом. Эукариоты на четыре-пять порядков увеличили размеры генома и физические размеры своих клеток. Последний общий предок эукариот уже обладал всеми этими признаками, в то время как у прокариотической клетки с прокариотическим эндосимбионтом (организма, с которого все началось) не было ни одного. Не осталось выживших промежуточных звеньев – ничего, что могло бы указать, как и почему эукариоты приобрели сложные черты.
Время от времени можно услышать, что эндосимбиоз, который предшествовал развитию эукариот, противоречит дарвиновскому сценарию эволюции: ведь это не градуализм, а неожиданный скачок, породивший “многообещающего урода”. Кое в чем замечание справедливо. Естественный отбор, действующий на бесконечную популяцию клеток в течение бесконечного времени, не сможет породить сложные клетки эукариотического типа иначе, чем путем эндосимбиоза. Такие процессы невозможно изобразить при помощи привычного нам бифуркационного дерева жизни. Эндосимбиоз противоположен бифуркации – в результате эндосимбиоза две ветви не расходятся, а напротив, сливаются. Но это единичное событие, один эволюционный эпизод, а ядро, как и другие черты из стандартного эукариотического набора, не могло возникнуть в один момент. Эндосимбиоз запустил цепочку эволюционных событий, которые привели к появлению эукариот, и этот процесс был уже в полном смысле дарвиновским.
Поэтому я не утверждаю, что зарождение эукариот происходило не по Дарвину. Единичный эндосимбиоз прокариот изменил адаптивный ландшафт, а уже после этого на нем смогла развернуться дарвиновская эволюция. Но как приобретение эндосимбионта изменило направленность естественного отбора? И было ли это изменение закономерным (что позволяет предположить похожее развитие событий и на других планетах)? Или исчезновение энергетических ограничений открыло просторы для бурного, непредсказуемого эволюционного процесса? Я беру на себя смелость утверждать, что хотя бы некоторые общие для эукариот признаки появились в результате тесного союза клетки-хозяина и ее эндосимбинта, и возникновение этих признаков можно предсказать из общих соображений. К таким признакам относится наличие ядра, половое размножение, наличие двух полов и разделение клеток тела на “бренные” соматические клетки и “бессмертные” клетки зародышевой линии.
С самого начала эндосимбиоз накладывает некоторые ограничения на порядок дальнейших событий. Так, ядро и система внутренних мембран должны были появиться лишь после эндосимбиоза. Кроме того, он ограничивает скорость эволюции. Эволюция по Дарвину подразумевает градуализм: когда отсутствуют резкие скачки, а закрепившиеся изменения дискретны и приносят небольшую выгоду. Однако это оказывается неверным в отношении самого генома, изменения которого могут представлять собой обширные делеции, дупликации, транспозиции или внезапную перестройку сигнальных путей из-за случайного включения или выключения регуляторных генов. Но сами по себе такие изменения неадаптивны. Подобно эндосимбиозу, они лишь прокладывают дорогу для естественного отбора. Если предположить, что ядро возникло мгновенно, значит, оно появилось в результате резкого эволюционного скачка, а не постепенного приспособления. Но ядро – не просто кладовка для ДНК, а чрезвычайно сложная, тщательно отлаженная структура. Оно содержит различные компоненты (рис. 26), например ядрышко (“фабрику”, где в огромных количествах производятся рибосомальные РНК), двойную мембрану (она усеяна ядерными порами – ошеломительной красоты молекулярными комплексами, сложенными из множества консервативных белков), эластичную ламину (упругую белковую сеточку, выстилающую ядерную мембрану и предохраняющую ДНК от механических повреждений).
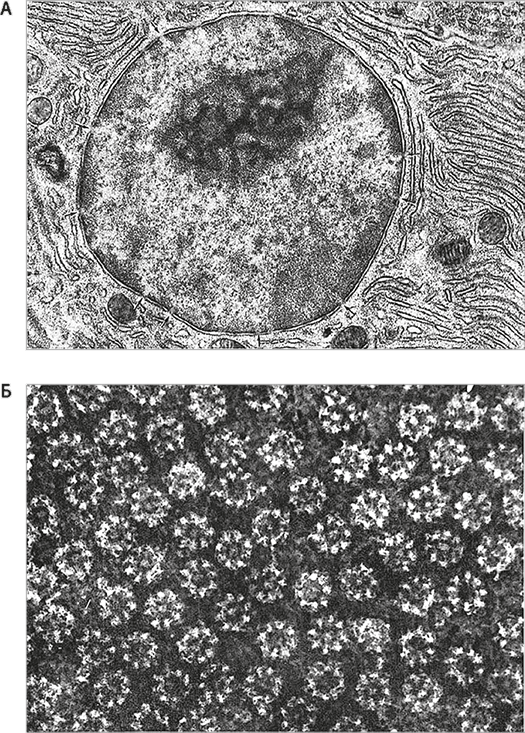
Рис. 26. Ядерные поры.
Изображения, полученные одним из основоположников электронной микроскопии Доном Фоссетом. Вверху (А) хорошо заметна окружающая эукариотическое ядро двойная мембрана и регулярно расположенные поры (отмечены стрелками). Более темные области внутри ядра соответствуют относительно неактивным регионам, где хроматин находится в “конденсированном” состоянии, в то время как в светлых областях активно идет транскрипция. Светлые промежутки между ядерными порами отражают активный транспорт внутрь и наружу ядра. На нижнем снимке (Б) мы видим скопление состоящих из множества белков ядерных поровых комплексов, которые вместе формируют механизм двустороннего транспорта. Ключевые белки поровых комплексов есть у всех эукариот. Следовательно, ядерные поры должны были иметься и у Последнего общего предка эукариот (LECA).
Такие структуры могут возникнуть лишь в результате отбора, действующего на протяжении долгого времени, и для их появления необходимо было усовершенствовать несколько сотен белков и обеспечить их слаженную работу. Этот процесс полностью соответствует дарвиновской теории эволюции и происходит медленно, но не обязательно медленно по геологическим меркам. В геологической летописи мы обычно наблюдаем долгие периоды стазиса, изредка прерываемые периодами быстрых изменений. В геологических масштабах такие изменения происходят быстро, но в масштабах поколений это далеко не всегда так: просто ограничения, которые прежде препятствовали изменениям, вдруг исчезают. Естественный отбор очень редко способствует изменениям. Чаще всего он, напротив, не позволяет популяции спуститься с пика адаптивного ландшафта, отсеивая отклоняющиеся формы. И лишь после значительных изменений адаптивного ландшафта (так сказать, адаптивных землетрясений) отбор может не только способствовать стазису, но и вызывать изменения. И тогда он может действовать с невероятной скоростью. Замечательный пример такого процесса – появление зрения. Глаза возникли во время “кембрийского взрыва”, и на их развитие потребовалось около 2 млн лет: по сравнению с сотнями миллионов лет докембрия это до неприличия быстро. Почему долгий застой вдруг сменился периодом исключительно быстрых изменений? Возможно, из-за повышения уровня кислорода отбор впервые стал способствовать развитию крупных подвижных животных: хищников и их жертв, обладавших глазами и крепким панцирем. Существует математическая модель, которая позволяет рассчитать, сколько времени может занять эволюция глаза из простого светочувствительного пятна какого-нибудь червя. Если предположить, что жизненный цикл червя составляет один год, а в каждом поколении морфологическое строение может измениться не более чем на 1 %, то для возникновения глаза потребуется лишь полмиллиона лет.
Сколько же нужно времени для развития ядра, полового размножения или фагоцитоза? Почему на это может потребоваться больше времени, чем на развитие глаза? Одна из задач, которые нам предстоит решить – расчет минимального времени, которое требуется для развития эукариот из прокариот. Перед тем как приступить, нужно больше узнать о последовательности событий, связанных с появлением эукариот. На первый взгляд, нет оснований думать, что для этого нужны сотни миллионов лет. Может быть, хватит и 2 млн? Если считать, что в день клетка делится один раз, за это время сменится около 1 млрд поколений. Сколько же их нужно? После того, как исчез энергетический барьер, не позволявший эукариотам достичь сложности, я не вижу причин, по которым эукариотические клетки не могли возникнуть за сравнительно короткий промежуток времени. По сравнению с 3 млрд лет прокариотического застоя это действительно выглядит как прорыв, хотя это, по сути, процесс эволюции по Дарвину.
То, что эволюция может идти быстро, еще не означает, что так было на самом деле. Но есть серьезные основания полагать, что эволюция эукариот действительно происходила быстро. Одно из таких оснований – нетерпимость природы к пустоте. Проблема именно в том, что у эукариот есть почти все, а у прокариот – почти ничего. Такое состояние должно быть нестабильным. В гл. 1 мы рассматривали архезоев: относительно просто устроенных одноклеточных эукариот, которых некогда ошибочно считали промежуточным звеном между прокариотами и эукариотами. Это сборная группа организмов, которые произошли от более сложных предков, обладавших полным набором эукариотических признаков. Архезои не являются эволюционной переходной формой. Это экологическая переходная форма: они занимают морфологическую нишу между прокариотами и эукариотами, заполняют эту зияющую пустоту. На первый взгляд, никакого вакуума нет, а есть непрерывный спектр морфологической сложности: от мобильных генетических элементов до гигантских вирусов, от бактерий до простых эукариот, от одноклеточных эукариот до многоклеточных организмов. И лишь недавно, когда выяснилось, что архезои вовсе не то, чем они кажутся, перед глазами предстал провал, отделяющий прокариот от эукариот.
Архезои не были вытеснены другими организмами, значит, в тех же экологических нишах могли выжить и простые переходные формы. Ничто не препятствует тому, чтобы их занимали настоящие переходные формы: клетки без митохондрий, или без ядра, или без пероксисом, или без системы мембран – без комплекса Гольджи и без эндоплазматического ретикулума. Если бы развитие эукариот шло медленно – десятки, сотни миллионов лет, – то должно было остаться множество стабильных промежуточных форм: клеток, у которых отсутствуют те или иные эукариотические признаки. Они должны занимать те же экологические ниши, которые сейчас занимают архезои, и некоторые из этих подлинных эволюционных переходных форм обязательно должны были выжить и стать мостом через пропасть. Но таких организмов нет! Несмотря на долгие поиски, не удалось найти ни одно дожившее до наших дней настоящее промежуточное звено. Если их не вытеснили другие организмы, то почему никто не выжил? Я полагаю, все из-за того, что они были генетически нестабильными. Есть не так уж много путей через пустоту, и большинство промежуточных форм погибло.
Это наводит на мысль о малом размере популяции. Большой размер популяции – признак эволюционного успеха. Если бы первые эукариоты процветали, они должны были активно распространяться, занимать новые экологические ниши и давать новые ветви. В таком случае они должны были быть генетически стабильными и хоть кто-нибудь выжил бы. Но этого не произошло. Вероятнее всего, первые эукариоты были генетически нестабильными, существовали в малых популяциях и очень быстро эволюционировали.
Еще одно доказательство – тот факт, что для всех эукариот характерны одни и те же свойства. До чего это странно! Всех людей объединяет набор одинаковых признаков: прямохождение, отсутствие шерсти, отставленные большие пальцы, крупный мозг, использование языка. Все это следствие нашего общего происхождения и непрерывного скрещивания (иначе говоря, секса). Это самое простое определение вида: популяция особей, скрещивающихся друг с другом. Группы особей, которые не скрещиваются друг с другом, обособляются и приобретают различающиеся признаки: становятся новыми видами. Но с первыми эукариотами такого не произошло – эукариот объединяет один набор основных признаков. Судя по всему, первые эукариоты представляли собой популяцию, в которой все особи скрещивались, то есть занимались сексом.
Можно ли добиться того же результата, пользуясь другими способами размножения? Не думаю. Бесполое размножение – клонирование – быстро приводит к расхождению, так как в разных популяциях накапливаются мутации, которые в разных условиях подвергаются действию отбора. Клонирование ведет к появлению идентичных организмов, но, как ни странно, по мере накопления мутаций это приводит к обособлению популяций. Секс, напротив, создает общий пул признаков в популяции, непрерывно перемешивает их и перебирает сочетания, противодействуя расхождению. То, что эукариотам присущ один набор признаков, наводит на мысль, что они возникли в популяции, где особи скрещивались. Из этого, в свою очередь, следует, что популяция была достаточно мала для того, чтобы все особи в ней скрещивались свободно. В такой популяции любая клетка, не способная к половому размножению, не выживет. Воистину: “Тесны врата и узок путь, ведущие в жизнь, и немногие находят их”.
Но существует и горизонтальный перенос генов (ГПГ), широко распространенный среди бактерий и архей. Как и половой, этот процесс задействует механизмы рекомбинации, создавая “пластичные” хромосомы с новыми комбинациями генов. Но ГПГ, в отличие от секса, – не взаимный процесс, в его рамках не происходит слияния клеток или полногеномной рекомбинации. Горизонтальный перенос – процесс хаотичный и однонаправленный: он не объединяет признаки внутри популяции, а напротив, приводит к усилению различий между особями. Рассмотрим кишечную палочку (E. coli). Ее клетка может содержать около 4 тыс. генов, а “метагеном” (совокупность генов, обнаруженных в разных штаммах E. coli; вид определен по последовательностям рибосомальной РНК) содержит более 18 тыс. генов. Из-за активного горизонтального переноса в разных штаммах может отличаться до половины генов – это куда больший разброс, чем среди всех позвоночных! Если кратко, то ни бесполое размножение, ни ГПГ – главные механизмы наследования у бактерий и архей – не могут объяснить единообразие эукариот.
Еще десять лет назад почти не было доказательств того, что секс – одно из ранних приобретений в эволюции эукариот. Многие виды тогда считались бесполыми, в том числе амебы и архезои, к которым относится лямблия и которых принимали за древнейших эукариот. По сей день никому не удалось застать лямблию в момент совокупления. Но то, чего мы не видели своими глазами, можно воссоздать при помощи технологий: ведь последовательность генома лямблии известна. Ее геном содержит полный набор работоспособных генов, нужных для мейоза (редукционного деления, необходимого для производства половых клеток), а его структура хранит следы регулярной половой рекомбинации. Более или менее то же можно сказать обо всех рассмотренных нами видах. Все известные эукариоты способны к половому размножению, за исключением тех, которые его потеряли. Как правило, такие организмы довольно быстро вымирают. Таким образом, общий предок эукариот был способен к половому размножению. Главные выводы таковы: половое размножение в эволюции эукариот возникло очень рано, и лишь его возникновением в малой генетически нестабильной популяции можно объяснить множество общих признаков эукариот.
О загадке строения наших генов
Гены эукариот имеют фрагментарное строение. После ранних исследований бактериальных геномов считалось, что и в наших хромосомах гены расположены в четком и однозначном порядке, как бусины на нитке. Но в конце XX века выявили неожиданную картину. Генетик Дэвид Пенни высказывался об этом так: “Если бы я участвовал в проектировании генома E. coli, я бы очень гордился этим проектом. Но если бы мне довелось приложить руку к человеческому геному, то было бы очень стыдно признаться, что я имею к этому отношение. Даже университетский комитет не сделал бы хуже”.
В эукариотическом геноме беспорядок. В составе генов короткие последовательности, кодирующие части белков (экзоны), чередуются с длинными некодирующими последовательностями – интронами. Как правило, один ген содержит несколько интронов. Размеры интронов могут довольно сильно различаться, но чаще всего они существенно длиннее кодирующих участков. Их последовательности всегда считываются сначала в РНК – матрицу, определяющую последовательность аминокислот в белке, а затем вырезаются из нее (“сплайсируются”) до того, как РНК достигает рибосом – расположенных в цитоплазме грандиозных фабрик по производству белка. Это непростая задача. Ею занимается другая замечательная белковая наномашина: сплайсосома. Вскоре мы вернемся к тому, насколько она важна, а пока отметим, что процедура вырезания интронов довольно причудлива. Если вырезать интрон не получится, рибосома пройдет по некодирующему участку, и после его считывания получится бесполезный белок. Рибосомы следуют инструкциям столь же неукоснительно и слепо, как бюрократы у Кафки.
Почему гены эукариот фрагментарны? Известно, что такое строение предоставляет некоторые преимущества. Например, на основе одного и того же гена можно получить разные белки, осуществляя сплайсинг (вырезание интронов). Именно так реализуется рекомбинационное многообразие иммунной системы. Кусочки белка комбинируются, формируя миллиарды антител, среди которых должно найтись подходящее почти для любого бактериального или вирусного белка. Связывание антитела с таким белком приводит в движение смертоносную машину иммунной системы. Однако иммунная система – это позднее изобретение, прерогатива крупных животных со сложным строением. А была ли какая-нибудь польза от фрагментарного строения генов на более ранних этапах? В 70-х годах XX века один из первопроходцев эволюционной биологии Форд Дулиттл предположил, что интроны должны были появиться еще на заре возникновения жизни. Суть “гипотезы ранних интронов” в том, что на ранних этапах эволюции отсутствовала подобная современной система исправления ошибок в ДНК, а значит, накопление ошибок в генах должно было происходить очень быстро и опасность генетического вырождения была велика. Учитывая высокую скорость появления мутаций, их общее число зависело от суммарной длины ДНК. Только малые геномы могли избежать вырождения. Приобретение интронов решает эту проблему. Как закодировать много белков при помощи короткой нити ДНК? Комбинируя кусочки! Это красивая идея, у которой до сих пор есть приверженцы, и Дулиттл не единственный. Эта гипотеза, как и другие хорошие гипотезы, позволяет довольно многое предсказать. К сожалению, ее предсказания ошибочны.
Главное предположение, вытекающее из этой гипотезы, таково: первыми возникли эукариоты. Интроны есть лишь у эукариот, а если интроны характерны для изначального, предкового состояния, то эукариоты должны быть самыми первыми клетками (прародителями бактерий и архей), которые впоследствии потеряли свои интроны в результате отбора на оптимизацию генома. С точки зрения филогенетики это абсурд. Современные данные полногеномного секвенирования показывают, что эукариоты, вне всяких сомнений, произошли от архейной клетки-хозяина и бактериального эндосимбионта. Самое первое разветвление дерева жизни – расхождение бактерий и архей. Эукариоты возникли гораздо позднее. Это подтверждают и палеонтологическая летопись, и энергетические расчеты.
Но если наличие интронов не было исходным состоянием, почему и как они появились? Похоже, дело в эндосимбионте. Да, у бактерий нет “настоящих интронов”, однако то, что интроны имеют бактериальное происхождение, почти не вызывает сомнений. Если точнее, они произошли от бактериальных генетических паразитов – мобильных элементов, которые получили название самосплайсирующиеся интроны II типа. Мобильные интроны – это фрагменты эгоистичной ДНК, прыгающие с места на место гены, которые копируют сами себя, встраиваясь в геном и снова покидая его. Это очень хорошо приспособленные для выполнения своей задачи машины. Они считываются в РНК как обычные гены, но затем эта РНК оживает, складываясь в сложную структуру, которая функционирует как молекулярные “ножницы”. При помощи этих “ножниц” паразиты вырезаются из длинных РНК-транскриптов, минимизируя урон для клетки-хозяина, и формируют активные комплексы, кодирующие обратную транскриптазу – фермент, способный синтезировать ДНК на матрице РНК. За счет этих комплексов копии интронов снова оказываются в геноме. Интроны, которые самостоятельно встраиваются и вырезаются из бактериального генома, считаются паразитическими генами.
“…На блохе живет блоху кусающая блошка”. Кто бы мог подумать, что геном – это настоящее змеиное логово, что он кишит паразитами. Но так и есть. Вероятно, мобильные интроны довольно древние. Они встречаются во всех трех доменах жизни, и у них, в отличие от вирусов, никогда не возникает необходимости покидать безопасное укрытие – клетку-хозяина: интроны копируются при ее делении. Живые организмы уже научились с ними жить.
У бактерий довольно хорошо получается с ними справляться: правда, неизвестно, как именно. Возможно, просто за счет силы отбора, который действует в крупных популяциях. Бактерии с неудачным расположением интронов, нарушающим правильную работу генов, проигрывают клеткам, у которых интронов нет. Или, может быть, сами интроны, приспосабливаясь, начинают вторгаться лишь в периферические области ДНК, что не должно сильно мешать клетке-хозяину. В отличие от вирусов, которые могут самостоятельно существовать вне клетки-хозяина и поэтому не слишком заботятся о том, чтобы ее не убить, мобильные интроны прекращают свое существование вместе с хозяином и не получают никакой выгоды, причиняя ему вред. Эту область биологии лучше всего рассматривать в терминах экономики (затраты и прибыли, “дилемма заключенного”, теория игр и т. д.). У бактерий и архей мобильные интроны встречаются нечасто, и их никогда не находят внутри самих генов – поэтому технически они вообще не являются интронами. Они накапливаются с низкой плотностью в промежутках между генами. Бактериальный геном обычно содержит не более 30 интронов на 4 тыс. генов. В эукариотических же геномах их число может достигать десятков тысяч. Малое количество интронов у бактерий – отражение установившегося за долгое время баланса, результат отбора, который в течение многих поколений действовал на обе стороны.
Вот такая бактерия вступила в эндосимбиоз с архейной клеткой-хозяином 1,5–2 млрд лет назад. Из современных форм к предку митохондрий наиболее близок один из видов альфа-протеобактерий, а нам известно, что у нынешних альфа-протеобактерий интронов довольно мало. Но как связаны древние генетические паразиты и структура эукариотических генов? Разве только механизмом РНК-ножниц, который обеспечивает сплайсинг мобильных бактериальных интронов, и отчасти логикой. Выше я упоминал сплайсосомы: белковые наномашины, вырезающие интроны из РНК-транскриптов. Они есть и у нас. Сплайсосома состоит не только из белков: ее центральная часть представляет собой все те же РНК-ножницы. Они осуществляют сплайсинг эукариотических интронов при помощи характерного механизма, который выдает происхождение этих “ножниц” от бактериальных самосплайсирующихся интронов (рис. 27).
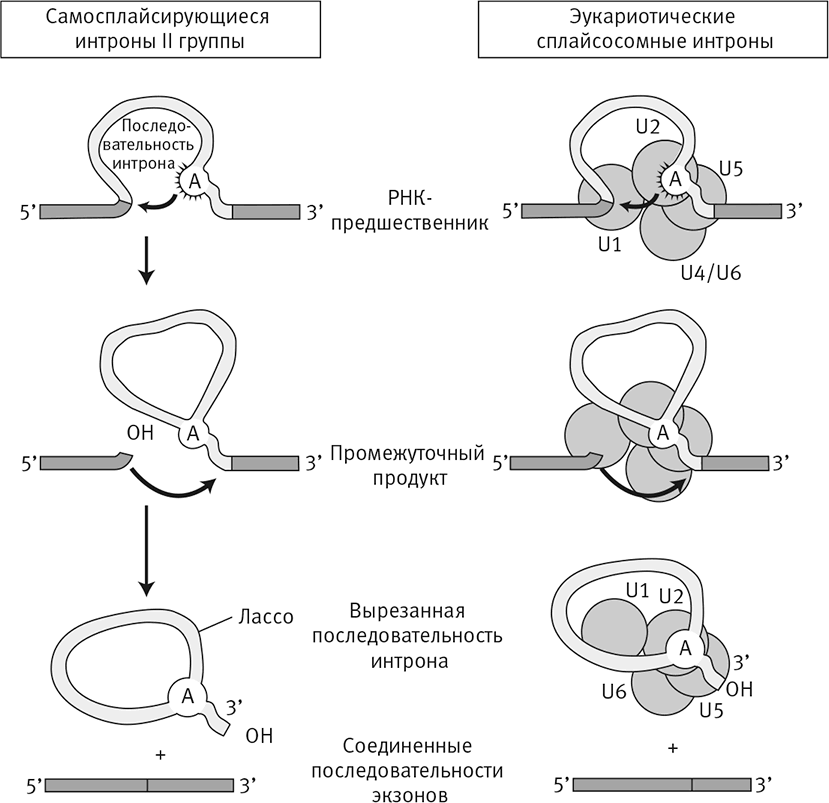
Рис. 27. Мобильные самосплайсирующиеся интроны и сплайсосома.
Эукариотические гены состоят из экзонов (последовательностей, кодирующих белки) и интронов (встроенных в гены длинных некодирующих последовательностей, которые вырезаются из информационной РНК перед синтезом белка). Интроны, вероятнее всего, произошли от паразитических ДНК-элементов из бактериальных геномов (слева), в результате накопления мутаций выродившихся в инертные последовательности внутри эукариотического генома. Они должны удаляться сплайсосомой (справа). В основе этого предположения лежит механизм сплайсинга. Бактериальный паразит (слева) вырезает себя сам, формируя отдельную интронную последовательность, в которой закодирована обратная транскриптаза: фермент, способный синтезировать ДНК на базе паразитических генов и вставлять их многочисленные копии в бактериальный геном. Эукариотическая сплайсосома (справа) – крупный белковый комплекс. Его работа зависит от каталитической РНК (рибозима), которая осуществляет сплайсинг при помощи того же самого механизма. Это позволяет предположить, что сплайсосома (и эукариотические интроны) произошла от мобильных самосплайсирующихся интронов II типа, занесенных бактериальным эндосимбионтом на ранних этапах эукариотической эволюции.
Так и происходит. Сама по себе нуклеотидная последовательность интронов не содержит ничего, что позволяло бы предположить их бактериальное происхождение. Они не кодируют никаких белков вроде обратной транскриптазы. Они самостоятельно не встраивают и не вырезают себя из ДНК. Они не являются мобильными генетическими паразитами. Они лишь “деклассированные элементы” генома, которые не делают абсолютно ничего. Однако мертвые интроны, опустившиеся на дно под грузом многочисленных мутаций, исказились до неузнаваемости и стали гораздо опаснее живых паразитов. Они уже не могут вырезать себя сами. Их должна удалять клетка-хозяин. И она делает это, орудуя “ножницами”, позаимствованными у ныне живущих родственников мертвых интронов. Сплайсосома – это эукариотическая машина, построенная на основе бактериального паразита.
Эту гипотезу выдвинули в статье 2006 года Билл Мартин и Евгений Кунин. Ученые предположили, что на заре возникновения эукариот произошло следующее: эндосимбионт, оказавшись внутри клетки-хозяина, обрушил на архею настоящий дождь генетических паразитов. Те стали заселять геном. Вторжение интронов сыграло большую роль в формировании эукариотического генома и послужило толчком для развития таких важнейших признаков, как наличие ядра. (Я бы прибавил – и полового размножения.) Да, идея может показаться похожей на байку, шатким основанием для которой послужили РНК-ножницы. Но в пользу этой гипотезы говорит и структура самих генов. Огромное число интронов (а оно измеряется десятками тысяч) в сочетании с их расположением внутри эукариотических генов указывает на их древнее происхождение. Более того, это позволяет судить не только о древности самих интронов, но и в целом об отношениях эндосимбионта и хозяина, очень тесных и полных жестокости. Даже если эти идеи и не совсем соответствуют действительности, мне кажется, они относятся к той разновидности ответов, которая нам нужна.
Интроны и происхождение ядра
Еще одна любопытная деталь: консервативность позиций многих интронов у эукариот. Рассмотрим ген цитратсинтазы, который кодирует белок, вовлеченный в базовые метаболические процессы клетки, и который присутствует у всех эукариот. Этот ген есть у людей, водорослей, деревьев, грибов и амеб. Поскольку с момента отделения этих групп от общего предка сменилось множество поколений, этот ген у них несколько отличается, но естественный отбор сохранил его функцию, и последовательность этого гена узнаваема у всех организмов. Это прекрасная иллюстрация общего происхождения эукариот и молекулярных основ естественного отбора. Но неожиданностью стало вот что: такие гены обычно содержат два или три интрона, позиции которых точно совпадают у далеко отстоящих друг от друга организмов, например людей и деревьев. Как это вышло? Есть лишь два правдоподобных объяснения: либо интроны независимо заняли одни и те же позиции потому, что те по каким-либо причинам оказались наиболее подходящими – либо интроны попали туда еще у общего предка эукариот и были унаследованы всеми его потомками. Впрочем, кто-нибудь из потомков мог их потерять.
Если бы нам было известно лишь несколько таких примеров, то подошло бы и первое объяснение. Но известно несколько тысяч интронов, встроенных в одни и те же позиции в сотнях генов, общих у всех эукариот, и поэтому такое объяснение кажется маловероятным. Гораздо более правдоподобнее версия о наличии этих интронов у общего предка эукариот. Отсюда следует, что вскоре после появления эукариотической клетки пришла волна инвазии интронов. В дальнейшем эти интроны должны были претерпеть ряд мутаций, которые нарушили их подвижность и зафиксировали их так же, как меловой контур указывает на положение тела на месте преступления.
Есть куда более убедительное доказательство того, что имела место ранняя инвазия интронов. Гомологичные гены (то есть произошедшие от одного предкового гена) делятся на два типа: ортологи и паралоги. Ортологи – это гомологичные гены, выполняющие у разных организмов одни и те же функции. Пример ортологичного гена – ген цитратсинтазы. Его ортологи есть у всех эукариот. Паралоги – это гомологичные гены, произошедшие от копий предкового гена внутри генома одной клетки. Паралогичные гены объединяются в генные семейства. Такие семейства могут содержать 20–30 генов, и каждый из них в конце концов оказывается приспособленным для выполнения конкретной задачи. Например, семейство генов гемоглобина содержит около 10 генов. Все они кодируют очень схожие белки, но функции этих белков немного различаются. Итак, ортологи – это гомологичные гены у разных видов, а паралоги – гомологичные гены в геноме одного организма. Конечно, целые семейства паралогичных генов могут быть и у разных видов, унаследовавших их от общего предка. Так, у всех млекопитающих в геноме закодированы семейства паралогичных генов гемоглобинов.
Мы можем поделить семейства паралогичных генов на древние и поздние паралоги, как это сделал Евгений Кунин. По Кунину, древние паралоги – это семейства паралогичных генов, которые есть у всех эукариот. У прокариот эти гены не дуплицированы. Поэтому мы предполагаем, что дупликация генов, в результате которой возникло семейство древних паралогов, произошла в эволюции эукариот еще до появления Последнего общего предка эукариот. Поздние паралоги – генные семейства, которые есть лишь у определенных групп эукариот, например, только у животных или у растений. В этом случае делаем вывод, что дупликация этих генов произошла позднее, уже в ходе эволюции группы.
Кунин предположил, что если на ранних этапах эволюции эукариот действительно происходила активная инвазия интронов, мобильные интроны должны были встраиваться в случайные позиции генов. Древние паралоги в тот период как раз активно дуплицировались, и, если волна ранней инвазии интронов тогда еще не утихла, мобильные интроны должны были встраиваться в новые гены – члены разрастающегося семейства паралогичных генов. Дупликации генов семейств поздних паралогов произошли уже после окончания ранней инвазии интронов, поэтому в новых копиях этих генов интроны должны были остаться в тех же позициях. Иными словами, позиции интронов в древних паралогах должны быть существенно менее консервативными. И это действительно так. Позиции почти всех интронов в поздних паралогах консервативны, в отличие от позиций интронов в древних паралогах – именно так, как было предсказано.
Все это наводит на мысль, что предки эукариот действительно пострадали от инвазии мобильных интронов, причем их заразили интронами собственные эндосимбионты. Но почему интроны так размножились у предков эукариот, если и бактерии, и археи, как правило, держат их в ежовых рукавицах? Этому есть два объяснения, и, вполне возможно, верны они оба. Согласно первому, предки эукариот – по сути еще прокариоты, точнее археи, – подверглись бомбардировке бактериальными интронами, оказавшимися прямо в их цитоплазме. Здесь работает принцип храповика. Эндосимбиоз – это эксперимент природы, который, как и всякий эксперимент, может провалиться. Если клетка-хозяин погибает, эксперимент проваливается. Но если погибает один из эндосимбионтов, еще не все потеряно – в клетке их много, и эксперимент продолжается. Но ДНК погибшего эндосимбионта попадает в цитоплазму клетки-хозяина и там может рекомбинировать с хозяйской ДНК, как и при обычном горизонтальном переносе.
Этот процесс нелегко остановить, он продолжается и по сей день: наши собственные ядерные геномы содержат тысячи обрывков митохондриальной ДНК – ядерно-митохондриальные псевдогены (Nuclear Mitochondrial Sequences, NUMTS), которые произошли именно таким путем. Псевдогены возникают время от времени и дают о себе знать лишь тогда, когда они повреждают гены, вызывая генетические заболевания. На ранних этапах эволюции эукариот, еще до того, как они приобрели ядро и стали эукариотами, подобный перенос ДНК должен был происходить куда чаще. Случайный перенос митохондриальной ДНК в геном – это плохо. Но еще хуже, если есть специальные механизмы, встраивающие мобильные интроны в определенные участки генома, избегая других. Как правило, интроны бактерий приспособлены к существованию внутри бактерий, а интроны архей – к существованию внутри архей. Но на заре возникновения эукариот все пошло наперекосяк: бактериальные интроны стали встраиваться в архейный геном, который сильно отличается от бактериального. Никаких механизмов борьбы с чужими интронами тогда не существовало, а иначе что могло удержать интроны от неконтролируемого размножения? Правильно: ничто! Появилась перспектива вымирания. Лучшее, на что можно было надеяться – влачить жалкое существование в виде маленькой популяции генетически нестабильных, слабых клеток.
Вторая причина активного размножения интронов на ранних этапах эволюции эукариот – слабое давление отбора. Отчасти это следствие того, что в малой популяции ослабленных клеток конкуренция между особями слабее, чем в огромных популяциях здоровых клеток. Но первые эукариоты обладали поистине небывалой устойчивостью к инвазии интронов. В конце концов, источником этих интронов были эндосимбионты – будущие митохондрии: энергетический рог изобилия и вместе с тем генетическая ноша. Такая генетическая и энергетическая обуза, как интроны, обходится бактериям недешево: маленькие клетки, у которых ДНК немного, воспроизводятся быстрее, чем крупные клетки с избыточным количеством ДНК. Бактерии стремятся оптимизировать свои геномы до минимально возможного размера. У эукариот же сильная асимметрия размеров геномов: их ядерные геномы свободно разрастаются за счет уменьшения геномов эндосимбионтов. Разрастание генома клетки-хозяина не преследует никакой цели – просто отбор не препятствует увеличению размеров генома, как происходит у бактерий. Снижение действия отбора на размер генома – палка о двух концах. Это позволяет эукариотам приобретать тысячи новых генов путем дупликации и рекомбинации – и дает возможность выжить с огромным количеством генетических паразитов. Эти два процесса неразрывно связаны. Эукариотические геномы оказались нашпигованы интронами просто потому, что это стало возможно с энергетической точки зрения.
Судя по всему, первые эукариоты были атакованы генетическими паразитами собственных эндосимбионтов. Как ни странно, сами по себе эти паразиты не представляют большой опасности. Настоящие проблемы начинаются, когда они погибают, оставляя “трупы” – засоряющие геном интроны. Клетке-хозяину приходится их вырезать, иначе происходит синтез бессмысленных белков. Это делают сплайсосомы, произошедшие от молекулярных РНК-ножниц мобильных интронов. Но сплайсосома, несмотря на всю свою внушительность, лишь отчасти решает проблему интронов. Ее недостаток в том, что она очень медленно работает. Даже современным сплайсосомам, за 2 млрд лет достигшим совершенства, требуется несколько минут, чтобы вырезать один интрон. Рибосомы же работают с бешеной скоростью – до 10 аминокислотных остатков в секунду. Им требуется менее 30 секунд, чтобы синтезировать типичный для бактериальных клеток белок длиной около 250 аминокислотных остатков. Если даже сплайсосоме удастся добраться до РНК (что нелегко, поскольку РНК нередко облепляет множество рибосом), это не сможет предотвратить синтез большого количества бесполезных белков, содержащих транслированные последовательности интронов.
Как избежать “катастрофы ошибок”? По мнению Мартина и Кунина – возвести барьер между незрелыми РНК и рибосомами. Именно таким барьером является ядерная мембрана, разграничивающая транскрипцию и трансляцию: внутри ядра с генов считываются кодирующие РНК, а снаружи ядра эти РНК транслируются в белки на рибосомах. Важнее всего вот что: медленный процесс сплайсинга происходит внутри ядра – до взаимодействия с рибосомами. В этом назначение ядра: служить преградой для рибосом. Это объясняет, почему эукариотам необходимо ядро, а прокариоты обходятся без ядра: у них нет проблем с интронами.
Но полностью сформировавшаяся ядерная мембрана не может появиться ниоткуда! Ее развитие должно было занять множество поколений. Тогда почему предки эукариот не вымерли в тот период, когда ядерная мембрана еще не появилась? Да, многие из них действительно вымерли, но загадка может оказаться не такой уж трудной. Ключ – в другом странном явлении, связанном с мембранами. Хотя генетический анализ указывает, что клетка-хозяин была настоящей археей и должна была иметь специфические для архей мембранные липиды, мембраны эукариот состоят из бактериальных липидов. Это известный факт. На ранних этапах эволюции эукариот архейные мембраны по каким-то причинам сменились бактериальными. Почему это произошло?
У этого вопроса есть два аспекта. Первый – практический: может ли это в принципе произойти? Ответ: да, может. Как ни странно, мозаичные мембраны, состоящие из смеси архейных и бактериальных липидов, действительно устойчивы – это известно из экспериментов. Следовательно, возможен постепенный переход от архейных мембран к бактериальным. Теоретически таким заменам ничто не препятствует, но на практике они происходят чрезвычайно редко. И мы подходим ко второй части вопроса: что могло вызвать эту замену? Ответ: эндосимбионт.
В ходе беспорядочного переноса ДНК от эндосимбионтов в геном клетки-хозяина должны были попасть бактериальные гены синтеза липидов. Можно предположить, что эти гены стали экспрессироваться и производить активные ферменты. Впоследствии именно эти ферменты синтезировали липиды бактериального типа для клеточных мембран, но на первых порах этот синтез, вероятно, был неконтролируемым. К чему ведет бесконтрольный синтез липидов? Если он происходит в водной среде, то они собираются в липидные пузырьки. Джефф Эррингтон из Ньюкасла продемонстрировал, что в живых клетках происходит то же самое: у бактерий мутации, увеличивающие синтез липидов, приводят к накоплению внутри клетки мембранных пузырьков, которые окружают геном. Подобно тому, как в холодную погоду бродяги иногда обматываются полиэтиленовыми пакетами, чтобы согреться, так и геномная ДНК обкладывается липидными мешочками, которые создают несовершенный барьер для рибосом, отчасти решающий проблему интронов. Этот барьер и должен быть несовершенным – ведь сквозь сплошную мембрану невозможен транспорт РНК к рибосомам. Дырявый барьер лишь замедлял этот процесс, задерживал РНК, давая сплайсосомам чуть больше времени на вырезание интронов, прежде чем рибосомы примутся за дело. Иными словами, случайное (но предсказуемое) событие предоставило естественному отбору основу для решения проблемы. Этой основой послужила кучка липидных мешочков, собравшихся вокруг генома. В результате же появилась ядерная мембрана, усеянная удивительно сложно устроенными ядерными порами.
Морфология ядерной мембраны хорошо согласуется с этой гипотезой. Липидные мешочки, подобно полиэтиленовым пакетам, можно расплющить. В поперечном сечении сплющенный мешочек состоит из двух близко отстоящих друг от друга параллельных слоев, то есть представляет собой двойную мембрану. Ядерная мембрана устроена так же: множество слитых вместе сплющенных пузырьков с ядерными поровыми комплексами в промежутке. В ходе клеточного деления ядерная мембрана распадается обратно на мелкие пузырьки. Затем они увеличиваются, снова сливаются друг с другом и образуют мембраны ядер, принадлежащих уже двум дочерним клеткам.
Набор генов, кодирующих ядерные структуры, вписывается в эту картину. Если бы ядро появилось до митохондрий, то компоненты его частей – ядерных пор, ламины и ядрышка – должны были бы кодироваться генами клетки-хозяина. Но это не так. Ядерные структуры состоят из химерической смеси белков. Некоторые кодируются генами бактериального происхождения, небольшая часть – архейного, остальные – генами, которые обнаружены лишь у эукариот. Такая мозаичность может объясняться лишь тем, что ядро появилось после приобретения митохондрий, в результате беспорядочного горизонтального переноса генов. Часто говорят, что в ходе эволюции эукариотической клетки эндосимбионты, превратившись в митохондрии, преобразились до неузнаваемости (впрочем, это преувеличение). Но мало кто понимает, что клетка-хозяин претерпела куда более серьезные изменения. Когда-то она была простой археей, которая приютила внутри себя эндосимбионтов. Они забросали наивную хозяйку своей ДНК и интронами, из-за чего ей пришлось приобрести ядро. И не только его: появилась перспектива полового размножения.
Начало секса
Выше упоминалось, что половое размножение у эукариот появилось очень рано. Еще, если помните, я намекал, что появление секса связано с инвазией интронов. Как именно? Для начала попробуем разобраться с самим понятием.
Настоящий секс, который практикуют эукариоты, предполагает слияние двух гамет (в нашем случае сперматозоида и яйцеклетки) с половиной набора хромосом у каждой. Мы, как и многие другие многоклеточные эукариоты, диплоидны: в наших клетках каждый ген присутствует в двух экземплярах – от отца и от матери. Если точнее, у нас по два экземпляра каждой хромосомы. На картинках в учебниках хромосомы выглядят так, будто это стабильные физические структуры, но это далеко не так. В ходе образования гамет хромосомы рекомбинируют – обмениваются друг с другом участками. Благодаря этому появляются новые комбинации генов, которых, скорее всего, прежде не бывало (рис. 28). Если вы пройдетесь по только что рекомбинировавшей хромосоме, перебирая ген за геном, то обнаружите, что она включает в себя гены обоих родителей. В ходе мейоза (редукционного деления) две хромосомы разделяются и попадают в гаплоидные гаметы, в которых каждая хромосома должна быть представлена одной копией. В итоге две гаметы (каждая содержит рекомбинантные хромосомы) сливаются – и получается оплодотворенная яйцеклетка, индивид с уникальной комбинацией генов, будущий ребенок.
Проблема с возникновением полового процесса состоит не в том, что для этого требовалось выработать множество новых механизмов. В ходе рекомбинации две гомологичные хромосомы конъюгируют, плотно прикрепляясь друг к другу. Затем хромосомы крест-накрест обмениваются некоторыми участками. Конъюгация хромосом и генетическая рекомбинация также осуществляются бактериями и археями в ходе горизонтального переноса генов, но она не всегда взаимна и применяется для того, чтобы починить поврежденные хромосомы или вновь обрести утерянные гены. Применяемые здесь молекулярные механизмы в основном те же. Рекомбинация при половом процессе отличается более широкими масштабами, а также тем, что обмен всегда взаимен. Половое размножение – это взаимная рекомбинация, происходящая по всей длине генома. Если такое и встречается у прокариот, то очень редко.
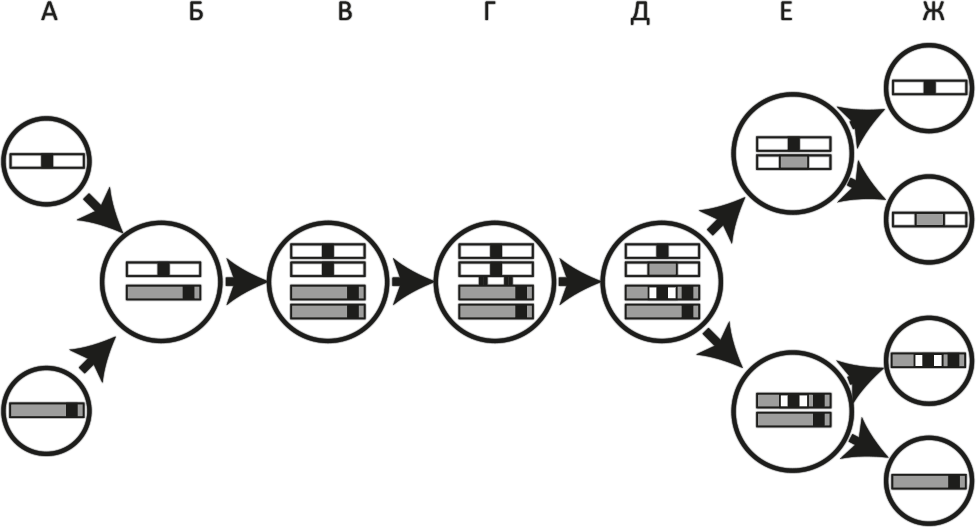
Рис. 28. Половой процесс и рекомбинация у эукариот.
Приведена упрощенная схема полового цикла: слияние двух гамет, за которым следует двухстадийный мейоз. В ходе мейоза осуществляется рекомбинация, что позволяет получить новые, генетически различные гаметы. Две гаметы, содержащие по одной хромосоме из гомологичной пары (хромосомы различаются генетически), сливаются (А). Гаметы образуют зиготу с двойным набором хромосом (Б). (Черные полоски могут обозначать и вредную мутацию, и полезный вариант какого-либо гена.) Перед мейозом хромосомы удваиваются (В). После этого в клетке образуется по две эквивалентных копии каждой хромосомы (каждая хромосома теперь состоит из двух сестринских хроматид). Затем две или более хромосомы попарно рекомбинируют (Г). Хромосомы совершают взаимообмен участками ДНК, и на выходе мы получаем обновленные хромосомы, каждая из которых включает фрагменты обеих исходных: и материнской, и отцовской (Д). При первом делении мейоза хромосомы разделяются, попадая в две разные клетки (Е), а в ходе второго деления образуются гаметы (Ж). Две из получившихся гамет эквивалентны исходным, две другие – нет. Если черные полоски обозначают вредные мутации, то в результате полового процесса образовалась одна гамета без мутаций и одна с двумя мутациями. Последняя будет элиминирована в ходе естественного отбора. Если черные полосы обозначают полезные мутации, то половой процесс объединил их в одной гамете, давая возможность естественному отбору воздействовать на них одновременно. Короче, половое размножение приводит к повышению многообразия гамет, делая их “заметнее” для отбора, и способствует элиминации вредных мутаций и распространению полезных.
В XX веке проблема смысла полового размножения была настоящей “царицей” биологических проблем, но сейчас мы уже понимаем, чем оно полезно: хотя бы тем, что пользоваться половым процессом лучше, чем размножаться исключительно бесполым путем (клонированием). Половой процесс разбивает устойчивые сочетания генов, позволяя естественному отбору “увидеть” гены по отдельности, перебрать по одному все наши признаки. Так можно избавиться от паразитов, приспособиться к изменяющимся условиям и поддерживать необходимый уровень разнообразия в популяции. Подобно тому, как средневековые каменотесы тщательно, со всех сторон, обрабатывали скульптуры для ниш в стенах соборов (ничто не укроется от ока Господа!), так и половой процесс тщательно обрабатывает геномы, позволяя естественному отбору детально, ген за геном, оценить собственные творения. Половое размножение делает наши хромосомы “пластичными”, позволяя разным вариантам генов – аллелям – комбинироваться, и благодаря этому естественный отбор выявляет даже незначительные различия между организмами.
Представьте себе сто генов, собранных в хромосоме, которая никогда не рекомбинирует. Отбор будет действовать лишь на целую хромосому, на весь набор генов сразу. Предположим, среди этих генов есть несколько критически важных: любая мутация в одном из них почти неизбежно приведет к гибели организма. Однако мутации в менее важных генах становятся почти невидимыми для отбора. Слабовредные мутации могут накапливаться в таких генах, а негативные эффекты будут компенсироваться наличием хороших аллелей нескольких важных генов. В результате качество хромосомы и приспособленность организма будут постепенно снижаться. Примерно это и происходит с Y-хромосомой у мужчин: если рекомбинации нет, значит, большая часть генов постепенно деградирует. Естественный отбор может обеспечить сохранение качества лишь критически важных генов. Целая половая хромосома может быть утеряна – так и случилось с Ellobius lutescens – одним из видов слепушонок.
Когда действует положительный отбор, ситуация становится еще хуже. Представим, что редкая полезная мутация предоставляет настолько значительные преимущества, что очень широко распространяется в популяции. Организмы, унаследовавшие новую мутацию, начинают доминировать, и в итоге она фиксируется во всей популяции: все организмы остаются с одним и тем же вариантом гена. Но естественный отбор может “видеть” только целую хромосому, а это значит, что остальные 99 генов, входящие в нее, также зафиксируются в популяции. Это явление называют “генетическим автостопом”: вместе с полезным аллелем одного гена в популяции закрепляются и другие – он как бы везет на себе аллели других генов. Это-то и плохо. Вообразите, что в популяции присутствует два или три варианта (аллеля) каждого гена и из них можно составить от 10 тыс. до 1 млн комбинаций. После фиксации все это многообразие пропадает, и популяция остается с одной-единственной комбинацией 100 генов – тех, которым выпало делить одну хромосому с полезным геном. Это катастрофическая потеря разнообразия. И, конечно, жалкие 100 генов – это грубое упрощение: у бесполых организмов тысячи генов остались лишь в одном варианте, а остальные были отброшены в ходе отбора. Такую форму отбора называют “селективным выметанием”. “Эффективный” размер популяции заметно сокращается, и это ставит бесполых животных под угрозу вымирания. И действительно: почти все бесполые растения и животные исчезли за несколько миллионов лет.
Эти два процесса – накопление слабовредных мутаций и потеря многообразия в ходе “селективного выметания” – в совокупности составляют селективную интерференцию. Если рекомбинации нет, то отбор, действующий на определенные гены, препятствует отбору других сцепленных с ними генов. Путем создания хромосом с комбинациями аллелей – “пластичных хромосом” – половой процесс позволяет отбору воздействовать на все гены по отдельности. Благодаря этому отбор видит все наши “пороки” и “добродетели” вплоть до нуклеотидов. В этом и заключается огромное преимущество полового размножения.
Но у секса есть свои серьезные недостатки: недаром он долго считался самой значительной из эволюционных проблем. Половое размножение разрушает заведомо удачные комбинации аллелей, которые хорошо подходят для определенных условий, и случайным образом перемешивает гены, которые позволяли преуспеть нашим предкам. В каждом поколении набор генов вновь перетасовывается, не оставляя никаких шансов на рождение полных двойников гениальных личностей: например, второго Моцарта. Что хуже, есть еще так называемая “двойная цена секса”. Когда делится бесполая клетка, она порождает две дочерние клетки. Каждая из них, в свою очередь, делится на две дочерние, и так далее. Численность популяции растет экспоненциально. Когда делится половая клетка, ее дочерние клетки должны слиться с аналогичными, чтобы сформировать новую клетку, которая породит еще две дочерних. Выходит, размер бесполой популяции в каждом поколении удваивается, а численность популяции организмов с половым размножением остается прежней. Вместо того чтобы просто получить собственную копию, приятную во всех отношениях, в рамках полового размножения вы сталкиваетесь с проблемой поиска партнера, что сопряжено с эмоциональными (и финансовыми) затратами. Самцы не достаются просто так. Клонируйте себя – и не будет нужды в поединках на рогах, павлиньих хвостах и заполненных мужчинами залах заседаний. Можно будет навсегда забыть об ужасных заболеваниях, передающихся половым путем, например СПИДе и сифилисе, и о вирусах и “прыгающих генах”, которые угрожают засорить наш геном.
Почему половое размножение у эукариот распространено так широко – загадка. Можно предположить, что его преимущества компенсируют затраты лишь при определенных обстоятельствах. Это подтверждается тем, что микробы могут делиться бесполым путем в течение жизни примерно 30 поколений перед тем, как побаловать себя спонтанным сексом – обычно это происходит в условиях стресса. Но я бы сказал, что секс распространен шире, чем это оправданно. Возможно, это связано с тем, что у Последнего общего предка эукариот уже имелось половое размножение, и все его потомки унаследовали от него это свойство. Хотя многие микроорганизмы перестали регулярно пользоваться половым процессом, мало кому удалось отказаться от полового размножения и не вымереть. Как видите, цена абсолютного отказа от секса также очень высока. Это относится и к первым эукариотам: те из них, у которых никогда не было секса – так сказать, не сумевшие до него “додуматься”, – были обречены на вымирание.
Но здесь мы возвращаемся к вопросу о горизонтальном переносе генов. Он аналогичен половому процессу в том отношении, что происходит генетическая рекомбинация и хромосомы становятся “пластичными”. До недавних пор считалось, что бактерии – непревзойденные мастера клонирования. Их численность растет экспоненциально. Одна-единственная бактерия E. coli, делясь каждые полчаса, за три дня способна образовать колонию, суммарная масса особей которой будет равна массе Земли (если, конечно, рост популяции не будет ничем ограничен). Кроме того, эти создания еще могут обмениваться генами при помощи горизонтального переноса, встраивая в свои хромосомы новые гены и утрачивая ставшие ненужными. Бактерии, устроившие вам кишечный грипп, могут отличаться от бактерий того же вида, живущих у вас в носу, на 30 % своего генома. Получается, бактерии наслаждаются преимуществами полового размножения (пластичностью хромосом) наряду со скоростью и простотой бесполого. Но их клетки не сливаются целиком и двух полов у них нет, поэтому многие недостатки полового размножения бактерий не затрагивают. Похоже, они взяли все лучшее от обоих миров. Так почему же ранние эукариоты перешли от горизонтального переноса генов к половому процессу?
Популяционные генетики-математики Салли Отто и Ник Бэртон выделили три условия, при которых наиболее выражены преимущества полового размножения: высокая скорость мутаций, сильное давление отбора и значительное разнообразие внутри популяции. Видимо, именно с этим пришлось столкнуться древним эукариотам.
Сначала рассмотрим частоту мутаций. При бесполом размножении высокая скорость мутагенеза увеличивает темпы накопления слабовредных мутаций. Кроме того, происходит снижение многообразия за счет “селективного выметания”, и селективная интерференция становится более жесткой. Скорость мутагенеза у древних эукариот, вероятно, была высокой, учитывая, что тогда происходила инвазия интронов. Насколько высокой, можно попытаться определить с помощью моделирования. Я занимаюсь этим вопросом с Эндрю Помянковски и Джезом Оуэном, который разбирается в физике и в то же время интересуется большими вопросами биологии. В данный момент Джез разрабатывает численную модель, которая позволила бы определить, в каких условиях половое размножение становится выгодным. Еще один фактор, который стоит принять во внимание – размер генома. Даже если скорость мутагенеза не изменяется (скажем, одна летальная мутация на 10 млрд “букв” ДНК), нельзя бесконечно увеличивать геном: по той или иной причине обязательно случится мутационная катастрофа. При такой скорости мутагенеза с клетками, геном которых включает менее 10 млрд нуклеотидов, все будет в порядке, а вот клетки с более длинным геномом погибнут, заполучив летальную мутацию. Приобретение митохондрий на ранних этапах эволюции эукариот должно было усугубить обе проблемы: они почти наверняка вызвали возрастание скорости мутагенеза и сделали возможным значительное (на несколько порядков) увеличение генома.
Вероятный ответ: половое размножение – единственный способ решения проблемы. Горизонтальный перенос генов теоретически может помочь избежать селективной интерференции при помощи рекомбинации, но работа Джеза показывает, что горизонтального переноса недостаточно. Чем больше геном, тем сложнее заполучить “правильный” ген путем горизонтального переноса. Это становится похоже на игру в рулетку. Единственный способ обеспечить полный набор работоспособных генов в геноме – регулярно производить рекомбинацию всего генома. Этого нельзя достичь путем горизонтального переноса. Здесь нужен “тотальный половой процесс”: лишь так можно осуществить полногеномную рекомбинацию.
Что насчет силы отбора? Интроны могут оказаться важны и здесь. Классический отбор, который действует на современные организмы, благоприятствует половому размножению: оно позволяет спасаться от паразитических инфекций и приспосабливаться к изменяющимся условиям. Но и с учетом этого отбор должен быть сильным, чтобы половое размножение стало выгоднее бесполого. Например, чтобы половое размножение получило селективное преимущество, паразиты должны быть очень широко распространены и сильно вредить. Несомненно, это верно и в отношении ранних эукариот, но им к тому же приходилось противостоять вторжению интронов – паразитических генов. Каким образом мобильные интроны могли способствовать появлению полового размножения? Дело в том, что рекомбинация по всему геному повышает многообразие: появляются клетки с неудачно расположенными интронами и другие, у которых интроны стоят на менее опасных местах. Затем естественный отбор оставляет лучшие варианты. Рекомбинация же при горизонтальном переносе лишь частична и не может обеспечить все многообразие вариантов, когда одна часть клеток очищает свои гены, а другая собирает большую долю мутаций. В великолепной книге “Демон Менделя” Марк Ридли проводит параллель между половым размножением и путем избавления от грехов, описанным в Новом завете: Христос умер, приняв на себя грехи человечества. Половое размножение также позволяет повесить все мутации, накопившиеся в популяции, на козла отпущения, а затем распнуть его.
Степень вариации между клетками также может быть связана с интронами. Археи и бактерии, как правило, имеют единственную кольцевую хромосому, а эукариоты – многочисленные линейные хромосомы. Почему? Из-за интронов. Когда они встраиваются и вырезаются из генома, могут возникать ошибки. Если у них не получается соединить обратно два конца хромосомы после автосплайсинга, в хромосоме остается разрыв. Одиночный разрыв в кольцевой хромосоме дает линейную хромосому, а несколько разрывов дают несколько линейных хромосом. Таким образом, из-за ошибок рекомбинации, вызванных мобильными интронами, у ранних эукариот могли появиться линейные хромосомы.
Из-за этого у эукариот должны были возникнуть серьезные проблемы с клеточным циклом. У разных клеток могло быть разное количество хромосом, и каждая имела собственный набор мутаций и делеций. К тому же ранние эукариоты могли приобретать новые гены и ДНК от своих митохондрий. Ошибки репликации, несомненно, могли приводить к дупликации хромосом. Трудно себе представить, чем в таких условиях мог помочь горизонтальный перенос генов. Но характерный для бактерий стандартный механизм рекомбинации – выстраивание хромосом друг относительно друга и включение в геном недостающих генов – должен был обеспечить накопление клетками новых генов и признаков. И лишь половое размножение позволяло обзаводиться работающими генами, при этом избегая прочих. Приобретая новые гены и ДНК в ходе полового процесса и рекомбинации, геномы ранних эукариот со временем расширялись. Накопление генов должно было отчасти решить проблемы генетической нестабильности, а наличие митохондрий со всеми его энергетическими выгодами спасало ранних эукариот от “энергетического штрафа” – в то время как бактериям приходилось его платить. Конечно, все это предположения, но математическое моделирование позволяет выбрать наиболее правдоподобные.
Как клетки физически разделяли свои хромосомы? Ответ можно найти в механизмах, которые бактерии применяют для разделения крупных плазмид – мобильных “кассет” с генами, отвечающими за определенный признак – например устойчивость к антибиотикам. При делении бактерий большие плазмиды обычно отделяются друг от друга при помощи структуры из микротрубочек, которая напоминает веретено деления эукариот. Очень вероятно, что ранние эукариоты позаимствовали механизмы разделения плазмид, чтобы отделять друг от друга свои многочисленные и разнообразные хромосомы. Таким способом делятся не только плазмиды: некоторые виды бактерий, похоже, разделяют свои хромосомы при помощи относительно динамичных веретен, а не используют, как обычно, клеточную мембрану. Может быть, детальное изучение мира прокариот позволит найти корни физического механизма разделения хромосом в митозе и мейозе.
Слияние клеток почти не встречается у бактерий с клеточной стенкой, хотя известно несколько способных сливаться друг с другом архей. Утрата клеточной стенки, несомненно, сильно облегчила слияние клеток: например, L-формы бактерий, потерявшие клеточную стенку, легко сливаются друг с другом. Многочисленные механизмы контроля слияния клеток у современных эукариот наводят на мысль, что их предков было трудно от этого удержать. В то время митохондрии даже могли способствовать слиянию клеток (эту идею выдвинул эволюционный биолог Нил Блэкстоун). Поразмыслим о проблемах, с которыми должны были столкнуться митохондрии. Будучи эндосимбионтами, они не могли просто покинуть клетку-хозяина и поселиться в другой, и поэтому их собственный эволюционный успех зависел от роста и развития хозяев. Если клетка-хозяин была повреждена мутациями и не способна расти, митохондрии были ограничены в возможностях и не могли размножаться. Но что, если они могли каким-либо образом индуцировать слияние хозяина с другой клеткой? От этого выигрывают оба. Клетка-хозяин приобретает комплементарный геном – и возможность осуществить рекомбинацию или же просто компенсировать мутации в своих генах за счет обладания чужими “чистыми” копиями тех же самых генов (словом, воспользоваться преимуществами аутбридинга). Благодаря тому, что после слияния получившаяся клетка снова могла расти, митохондрии также получали возможность размножаться. То есть ранние митохондрии должны были склонять эукариот к сексу друг с другом! Таким путем митохондрии смогли решить свою проблему, но, по иронии судьбы, это решение привело к другой, даже более широкой, проблеме: к конкуренции митохондрий. Разрешить ее можно было лишь при помощи двух полов – еще одной загадки полового размножения.
Два пола
“Ни один биолог, изучающий половое размножение, не станет исследовать, что может происходить с организмами, у которых существует три пола или больше. Но как иначе можно узнать, почему полов всегда два?” – говорил Рональд Фишер, один из отцов-основателей эволюционной генетики. Этот вопрос все еще ждет ответа.
Если поразмыслить, то два пола – это свойство худшего из миров. Только представьте: если бы все люди принадлежали к одному полу, мы смогли бы спариваться друг с другом и выбор партнера стал бы вдвое проще. А если по каким-либо причинам оказалось бы невозможно иметь лишь один пол, три или четыре пола все равно лучше, чем два. Даже если нельзя спариваться с партнерами своего пола, в нашем распоряжении были бы 2/3 или 3/4 популяции, а не ее половина. Конечно, в спаривании участвуют двое, но нет очевидных причин, почему партнеры не могут быть одного пола, или принадлежать к разным полам, или вообще быть гермафродитами. Трудности, с которыми сталкиваются гермафродиты, приоткрывают суть проблемы: ни один из партнеров не желает становиться “женщиной”, поскольку это очень накладно. Гермафродитные плоские черви вырастают до чудовищных размеров, чтобы их сложнее было оплодотворить, устраивают яростные поединки, фехтуя на пенисах, а сперма прожигает дыры в телах поверженных противников (то есть половых партнеров). Эта история, конечно, оживляет повествование, но в качестве аргумента она не годится: здесь ситуация развивается уже исходя из того, что женская особь тратит больше ресурсов (вообще-то это не очевидно). Почему так должно быть? В чем различие между мужским и женским полами? Это разделение произошло чрезвычайно давно и не имеет ничего общего ни с половыми хромосомами, ни даже с яйцеклетками и сперматозоидами. Два пола, или, по крайней мере, два типа спаривания, обнаружены даже у одноклеточных эукариот – некоторых водорослей и грибов. Их гаметы имеют микроскопические размеры и внешне неразличимы, но, несмотря на это, при выборе партнера для оплодотворения они столь же разборчивы, как и мы.
Одно из главных различий между двумя полами относится к наследованию митохондрий: при спаривании одна половая клетка передает свои митохондрии потомству, а другая – нет. Это относится и к людям (все наши митохондрии унаследованы от матери – в яйцеклетке их около 100 тыс.), и к водорослям, например Chlamydomonas. Хотя эти водоросли производят одинаковые гаметы (это явление называется изогамия), лишь одна из гамет передает свои митохондрии потомкам. Митохондрии второй гаметы уничтожаются, перевариваясь изнутри. Точнее, происходит избирательное переваривание митохондриальной ДНК. Видимо, проблема заключается в митохондриальных генах, а не в морфологической структуре. Странная ситуация: митохондрии склоняют клетки к половому процессу, но в результате не передаются от клетки к клетке, поскольку половина их переваривается. В чем тут дело?
Наиболее убедительное объяснение – эгоистический конфликт. Между генетически идентичными клетками не может быть настоящей конкуренции. По этой причине наши клетки ведут себя мирно и, взаимодействуя, формируют наши тела. Все наши клетки генетически идентичны – мы представляем собой гигантские клоны. Но генетически разнородные клетки конкурируют друг с другом – так происходит при мутациях, вызывающих рак. Сходная ситуация возникает, если внутри одной клетки оказываются генетически разнородные митохондрии. Те клетки или митохондрии, которые размножаются быстрее, будут вытеснять остальных, даже если это разрушительно для организма – может возникнуть нечто вроде митохондриального рака. Ведь клетки представляют собой автономные самовоспроизводящиеся системы, всегда готовые расти и делиться, если есть возможность. Французский нобелевский лауреат Франсуа Жакоб однажды сказал, что мечта каждой клетки – стать двумя клетками. Удивляет не то, что клетки часто делятся, а то, что они могут долго воздерживаться от деления, и благодаря этому наше тело может нормально функционировать. По этим причинам смешивать внутри одной клетки две популяции митохондрий – это искать проблем на свою голову.
Эту идею, предложенную несколько десятилетий назад, отстаивал такой крупный эволюционный биолог, как Билл Гамильтон. Но она себя не оправдала. Во-первых, известны исключения, когда митохондрии свободно смешиваются и это не приводит к неприятностям. Во-вторых, представим себе митохондриальную мутацию, дающую преимущество в скорости размножения. Митохондрии с такой мутацией будут вытеснять все остальные. Если эта мутация летальна, то мутанты вымрут вместе с клетками-хозяевами. Если же нет, то мутанты будут распространяться в популяции. Любые генетические механизмы, ограничивающие их распространение (изменения в ядерных генах, предотвращающие смешение митохондрий и т. д.), должны возникать быстро, чтобы успеть остановить распространение мутантных митохондрий. Малейшее промедление – и приобретение такого гена уже не поможет: это бесполезно, если мутантные митохондрии закрепились в популяции. Эволюция слепа. Она не способна предугадать возникновение следующего митохондриального мутанта. И третье: у митохондрий осталось мало генов, что наводит на мысль, что быстрое размножение митохондрий вовсе не так опасно. Причин тому может быть множество, но одна из них, несомненно, – действующий на митохондрии отбор в сторону увеличения скорости размножения. Это, в свою очередь, означает, что существовали многочисленные мутации, увеличивающие скорость размножения митохондрий, и что возникновение двух полов не устранило эти мутации.
Я предположил: проблема в том, что гены митохондрий должны быть адаптированы к генам ядра. (Подробнее расскажу в следующей главе.) Для эффективных дыхательных процессов гены в митохондриях и в ядре должны работать сообща, и мутации в обоих геномах могут вредить приспособленности. Я предположил, что однородительское наследование, при котором лишь одна гамета передает потомкам митохондрии, может способствовать коадаптации двух геномов. Эта идея мне очень нравится, но результаты работы Зены Хадживасилиу, весьма способного математика с интересом к биологии, которая работает над диссертацией вместе со мной и Эндрю Помянковски, заставили меня взглянуть на нее под другим углом.
Хадживасилиу продемонстрировала, что однородительское наследование действительно улучшает коадаптацию митохондриального и ядерного геномов. Причина довольно проста и сводится к эффектам выборки. Представьте себе клетку с сотней генетически разнородных митохондрий. Вы берете одну из них, помещаете в другую клетку и заставляете размножаться, пока не получите сто митохондрий. За исключением небольшого числа мутаций, все эти митохондрии будут клонами. Теперь вы делаете то же самое со следующей митохондрией и продолжаете, пока не клонируете все сто. В каждой из сотни клеток будет своя популяция митохондрий – одни популяции будут хорошими, другие – плохими. Таким образом, вы увеличили разнообразие. Если бы вы сто раз скопировали клетку целиком, то каждая дочерняя клетка получила бы примерно такую же смесь митохондрий, как у родительской. Эти клетки для естественного отбора неразличимы, поскольку слишком похожи одна на другую. Но клонирование в сочетании с эффектом выборки позволяет получить набор клеток с разной степенью приспособленности, среди которых найдутся клетки, приспособленные лучше исходной.
Этот пример иллюстрирует суть однородительского наследования. За счет выбора небольшого числа митохондрий лишь от одного из двух родителей однородительское наследование увеличивает разнообразие митохондриального состава среди оплодотворенных яйцеклеток. Это разнообразие становится заметным для естественного отбора, отсеивающего плохо приспособленные клетки. Таким образом, приспособленность целой популяции со временем возрастает. Любопытно, что это дает почти те же преимущества, что и половое размножение, однако последнее увеличивает разнообразие ядерных генов, а наличие двух полов увеличивает разнообразие митохондрий между разными клетками. Все очень просто. Ну, нам так казалось.
Мы оценивали приспособленность в условиях наличия и отсутствия однородительского наследования митохондрий, но не рассматривали, что может произойти, если ген однородительского наследования возникнет в популяции клеток, в которой обе гаметы передают потомкам свои митохондрии. Будет ли этот ген распространяться и закрепляться в популяции? Если да, то мы будем наблюдать возникновение двух полов, один из которых передает митохондрии, а второй – нет. Чтобы проверить, возможно ли это, мы разработали модель, в рамках которой сравнили гипотезы коадаптации, эгоистического конфликта и простого накопления мутаций. Результаты получились неожиданными и, на первый взгляд, разочаровывающими. Модель показала: ген однородительского наследования не стал бы распространяться в популяции и, уж конечно, не закрепился бы в ней.
Проблема в том, что выгода однородительского наследования зависит от числа мутантных митохондрий в популяции: чем меньше мутантов, тем меньше пользы оно приносит. Иными словами, затраты и преимущества такого наследования не фиксированы, а зависят от числа мутантных митохондрий в популяции, которое может быть снижено за несколько поколений с однородительским наследованием (рис. 29). Мы показали, что наследование этого типа действительно улучшает приспособленность популяции во всех трех моделях, но по мере того, как в популяции распространяется ген однородительского наследования, преимущества обладания этим геном падают. В конце концов он даже становится вредным: основной его недостаток в том, что клетки с этим геном могут скрещиваться с меньшей частью популяции. Популяция приходит к равновесию, когда в ней содержится всего 20 % клеток с однородительским наследованием митохондрий. При высоком уровне мутаций их доля может достигать 50 %, но особи второй половины популяции могут продолжать спариваться между собой, что дает три пола. Суть в том, что наследование митохондрий не приводит к возникновению двух полов. Однородительское наследование увеличивает разнообразие гамет, тем самым увеличивая приспособленность популяции, но этого явно недостаточно для возникновения типов спаривания.
Мне не очень-то понравилось прямое опровержение собственной идеи. В конце концов я был вынужден признать, что нет реальных условий, при которых однородительское наследование митохондрий могло бы привести к возникновению двух типов спаривания. Их появление должно быть обусловлено иными причинами. Но если и так, однородительское наследование все равно существует, и если бы модель не позволяла объяснить его существование, это означало бы, что она просто неверна. Фактически удалось показать, что если по каким-либо причинам два типа спаривания уже появились, то при определенных условиях (если митохондрий много и они часто мутируют) однородительское наследование может зафиксироваться. Наше заключение казалось неопровержимым, и выводы вполне уживались с известными в природе исключениями из однородительского наследования митохондрий. К тому же оно объясняло, почему однородительское наследование митохондрий присуще почти всем многоклеточным организмам, например людям, у которых, как правило, много митохондрий и они часто мутируют.
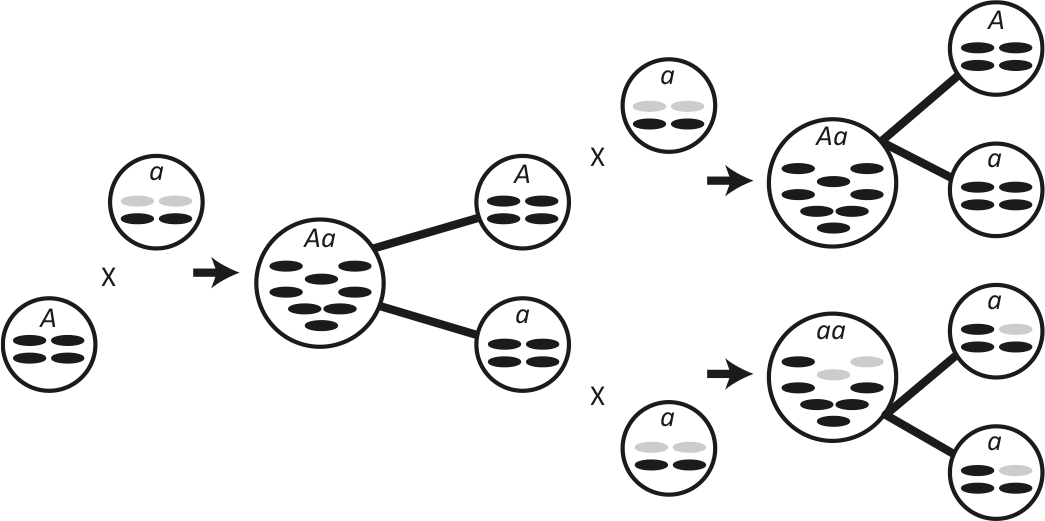
Рис. 29. Повышающие приспособленность мутации “просачиваются” в популяцию при однородительском наследовании митохондрий.
А и а – гаметы, несущие в ядре аллели определенного гена. Гаметы а передают митохондрии потомкам, если сливаются с другой гаметой а. Гаметы А – мутанты с однородительским наследованием: если гамета А сливается с гаметой а, передаются лишь митохондрии гаметы А. На первом скрещивании изображено слияние с А и а с образованием зиготы, которая несет в ядре оба аллеля (Аа), но унаследованы лишь митохондрии гаметы А. Если а содержит дефектные митохондрии (показаны серым цветом), они будут вычищены однородительским наследованием. Затем зигота дает гаметы двух типов: А и а. Они сливаются с гаметами, содержащими дефектные митохондрии. Вверху гаметы А и а дают зиготу Аа с унаследованными от гаметы А митохондриями, не содержащую дефектных митохондрий. Внизу изображено слияние двух гамет а, и дефектные митохондрии передаются зиготе аа. Каждая зигота, Аа и аа, образует гаметы. Митохондрии гамет а теперь “очищены” двумя раундами скрещивания с однородительским наследованием. Это улучшает приспособленность гамет с двуродительским наследованием митохондрий, поэтому улучшение приспособленности, которую дает аллель А, как бы “просачивается” в популяцию и в конечном счете прекращает распространение других вариантов этого гена.
Это прекрасный пример, иллюстрирующий важность математических моделей в популяционной генетике: гипотезы должны проверяться любыми доступными методами. В нашем случае формальная модель показала, что однородительское наследование митохондрий не может фиксироваться в популяции, если прежде в ней уже не было двух типов спаривания. Мы доказали это, насколько смогли, строго. Но еще не все потеряно. Разница между типами спаривания и “истинными” полами (когда мужские и женские особи явно различаются) довольно туманна. У многих растений и водорослей есть и пол, и типы спаривания. Быть может, стоило рассматривать понятие “пол” в другом ключе и сосредоточиться на возникновении “истинных” полов, а не типов спаривания, которые внешне могут выглядеть одинаково? Возможно, наши определения того, что такое пол, были неверны и нам следовало рассматривать эволюцию истинных полов, а не двух якобы идентичных типов спаривания. Может ли однородительское наследование быть причиной различий между истинными полами у животных и растений? Если да, то типы спаривания могли возникнуть по каким-либо иным причинам, а развитие настоящих полов могло быть вызвано как раз наследованием митохондрий. Честно признаться, эта идея выглядела малообещающей, но к ней стоило присмотреться. Мы совсем не ожидали, что придем к поразительному ответу, причем именно благодаря тому, что отправились мы не от стандартной гипотезы об универсальности однородительского наследования, а от неутешительных выводов нашей предыдущей работы.
Бессмертная зародышевая линия и бренное тело
В клетках животных содержится огромное число митохондрий, и мы заставляем их безостановочно работать, чтобы хватило энергии для активной жизни. Это должно приводить к высокому уровню мутагенеза в митохондриях, верно? Более или менее да. В каждой нашей клетке от нескольких сотен до нескольких тысяч митохондрий. Мы не знаем точную частоту их мутаций (ее очень трудно измерить непосредственно), но известно, что наши митохондриальные гены эволюционируют в 10–50 раз быстрее, чем ядерные. Отсюда следует, что однородительское наследование митохондрий у животных должно зафиксироваться особенно быстро. Наша модель показала, что оно действительно будет гораздо легче закрепляться у многоклеточных организмов, чем у одноклеточных. И в этом нет ничего удивительного.
Но легко впасть в заблуждение, если рассматривать эти процессы на собственном примере. Первые животные были совсем не похожи на нас и представляли собой кого-то вроде губок или кораллов – сидячих фильтраторов, которые не перемещались в пространстве (по крайней мере во взрослых стадиях). Неудивительно, что у них было не очень много митохондрий и их митохондриальные гены мутировали реже (во всяком случае реже ядерных). Из этого исходил Арунас Радзвилавичюс, когда работал над диссертацией. Радзвилавичюс – талантливый физик, которого привлекли большие биологические задачи. Вообще забавно, что самые интересные физические проблемы возникают сейчас именно в биологии.
Арунас понял, что обычные деления клеток в многоклеточных организмах и однородительское наследование митохондрий приводят к похожему результату: они увеличивают вариабельность клеток. Почему? Во время каждого клеточного деления популяция митохондрий случайным образом распределяется между дочерними клетками. Если в клетке небольшое число мутантных митохондрий, вероятность того, что эти мутанты будут распределены точно поровну, низка – куда вероятнее, что одна из дочерних клеток получит больше мутантных митохондрий, чем другая. Если повторить деление много раз, мы получим большое разнообразие. Некоторые прапрапраправнуки исходной клетки унаследуют больше мутантных митохондрий, чем другие. Хорошо это или плохо, зависит от того, какие именно клетки и в каком количестве унаследовали мутантные митохондрии.
Представьте себе организм вроде губки, все клетки которого очень похожи друг на друга. У такого организма нет дифференциации клеток на множество специализированных тканей и органов, например мозг и кишечник. Если вы разрежете живую губку на кусочки (не пытайтесь повторить это дома!), эти кусочки регенерируют. Губка может себе это позволить, потому что стволовые клетки у нее почти повсюду и из них могут образовываться и соматические (образующие тело) клетки, и клетки зародышевой линии. В этом отношении губки похожи на растения: ни у тех, ни у других нет специализированных зародышевых клеток, которые отделялись бы на ранних этапах развития. Их гаметы способны развиваться из стволовых клеток во многих тканях. Это различие имеет решающее значение. У нас есть особая линия зародышевых клеток, которые отделяются на ранних стадиях эмбрионального развития. У млекопитающих в норме не образуется гамет, например, из стволовых клеток печени. А вот для губок, кораллов и растений это нормально: половые органы у них могут вырастать в самых разных местах. Эти различия обычно объясняют конкуренцией клеток, но это не очень убедительно. Арунас же обнаружил, что у всех организмов без зародышевой линии есть общая черта: в их клетках мало митохондрий, и эти митохондрии редко мутируют. А редкие мутации могут уничтожаться путем сегрегации.
Вот как это работает. Многочисленные деления увеличивают разнообразие митохондриального состава между клетками. То же самое касается клеток зародышевой линии. Если зародышевые клетки отделяются на ранних стадиях развития, между ними не может быть большого разнообразия, поскольку они претерпели малое число делений. Но если зародышевые клетки развиваются из случайно взятых клеток зрелых тканей, то различий между ними будет гораздо больше (рис. 30). Большое число актов деления клеток означает, что в некоторых зародышевых клетках накопилось больше мутаций, чем в других. Какие-то будут почти идеальны, а в некоторых бардак: словом, будет наблюдаться большое разнообразие. Как раз то, что нужно для естественного отбора: он вычищает плохие клетки, оставляя в живых хорошие. По мере смены поколений качество зародышевых клеток растет, и формировать зародышевые клетки из случайных клеток зрелых тканей оказывается уместнее, чем отделять их на ранних стадиях развития и погружать в спячку.
Хотя повышенное разнообразие – благо для зародышевых клеток, оно может обернуться катастрофой для зрелого организма. Плохие зародышевые клетки вычищаются отбором – остаются лучшие, которые и дадут начало новому поколению. Но как быть с дефектными стволовыми клетками, образующими новые ткани взрослого организма? Скорее всего, из них сформируются ткани с нарушенными функциями, которые могут оказаться не в состоянии поддерживать работу организма. Здоровье организма в целом зависит от состояния самого больного из его органов. Если вдруг я получу инфаркт, прекрасное состояние моих почек уже не будет иметь значения: я умру, и все мои здоровые органы умрут со мной. Таким образом, высокое разнообразие митохондриального состава в организме имеет и преимущества, и недостатки, и ущерб для целого организма может вполне перекрыть те преимущества, которые получают зародышевые клетки. Уровень митохондриального разнообразия, при котором оно становится скорее вредным, чем полезным, зависит от числа тканей в организме и от скорости мутаций.
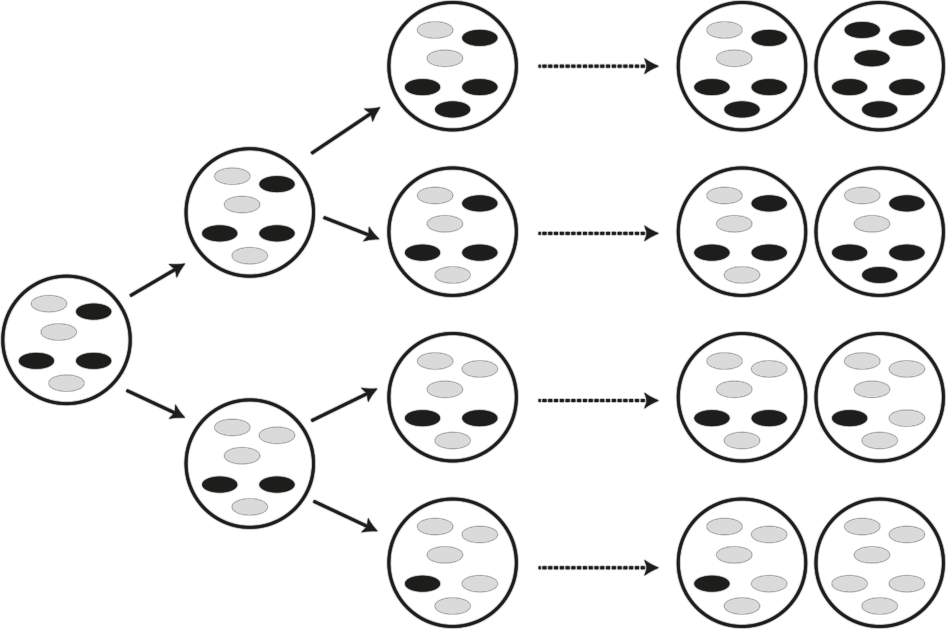
Рис. 30. Случайное распределение митохондрий увеличивает разнообразие клеток.
При делении клетки, содержащей разнотипные митохондрии, они удваиваются, а затем примерно поровну делятся между двумя дочерними клетками. Соотношение разных типов митохондрий в дочерних клетках после каждого деления будет немного иным. Со временем различия усиливаются, так как с каждым делением популяции митохондрий все дальше расходятся. Предположим, что получившиеся дочерние клетки (справа) – это гаметы. В результате предшествующей серии делений образовались гаметы, сильно отличающиеся друг от друга: некоторые хорошо приспособлены, другие – нет. Это готовит почву для естественного отбора, и однородительское наследование оказывает такой же эффект. Это хорошо. Теперь представим, что клетки на схеме справа – это прогениторные клетки, из которых будут формироваться новые ткани и органы. В этом случае избыточное разнообразие приведет к катастрофе. Некоторые ткани будут функционировать нормально, а жизнедеятельность остальных будет нарушена, что снизит общую приспособленность организма. Уменьшить разнообразие прогениторных клеток можно путем увеличения количества митохондрий в зиготе. Это повышает вероятность того, что полученные дочерними клетками наборы митохондрий окажутся примерно равнозначны (эффект выборки). Это может быть достигнуто увеличением размера яйцеклетки, что приводит к возникновению анизогамии (появляются крупные яйцеклетки и маленькие сперматозоиды).
Чем больше различных тканей во взрослом организме, тем выше вероятность, что одна из жизненно важных тканей соберет в себе наихудшие митохондрии. Но если организм состоит из ткани лишь одного типа, это уже не проблема: нет никаких отдельных органов, нарушение работы которых подорвало бы здоровье всего организма. Для простого организма с тканями одного типа увеличение разнообразия митохондриального состава клеток – несомненное благо: при этом растет качество гамет и нет особенного вреда для организма. Поэтому мы предположили, что у первых животных, у которых (вероятно) был низкий уровень митохондриальных мутаций и небольшое число типов тканей, митохондрии наследовались от обоих родителей, а обособления зародышевых клеток не было. Но когда устройство этих древних животных чуть усложнилось и тканей у них стало больше, повышенное разнообразие митохондрий в клетках тела стало снижать приспособленность зрелого организма (из-за него неизбежно возникают дефектные ткани – как в сценарии с инфарктом). Теперь для увеличения приспособленности взрослых особей потребовалось снижать митохондриальное разнообразие, чтобы ткани при образовании получали схожий набор митохондрий, большинство которых хорошо функционирует.
Простейший способ уменьшить митохондриальное разнообразие в тканях зрелого организма – это увеличить число митохондрий в яйцеклетке. По законам статистики, если большая популяция разделится на много частей, разнообразие окажется меньше, чем если малая популяция будет попеременно удваиваться и делиться до образования такого же количества частей. В итоге увеличение размеров яйцеклетки и все большее увеличение числа митохондрий в ней становится выгодным. Согласно нашим расчетам, ген, обусловливающий увеличение размеров яйцеклетки, будет распространяться в популяции простых многоклеточных организмов как раз потому, что он уменьшает митохондриальное разнообразие клеток тканей зрелого организма, устраняя возможные вредоносные различия. С другой стороны, низкое митохондриальное разнообразие наносит ущерб гаметам, которые становятся похожими друг на друга и неразличимыми для естественного отбора. Как совместить две противоположные тенденции? Очень просто! Если одна гамета – яйцеклетка – увеличивается, а вторая гамета уменьшается, превращаясь в сперматозоид, то решаются обе эти проблемы. За счет больших размеров яйцеклетки снижается митохондриальное разнообразие тканей, в результате чего увеличивается приспособленность взрослых особей. В то же время вклад со стороны сперматозоидов в наследование митохондрий снижается, и все заканчивается однородительским наследованием, при котором потомкам передаются митохондрии лишь одной гаметы. А однородительское наследование митохондрий, как мы убедились, увеличивает разнообразие между гаметами и повышает их приспособленность. Иными словами, такие простейшие свойства, как анизогамия (когда гаметы – сперматозоид и яйцеклетка – отличаются друг от друга) и однородительское наследование служат отправными точками для развития организмов с большим числом тканей.
Все это подразумевает низкую частоту митохондриальных мутаций. Это справедливо для губок, кораллов и растений, но не “высших” животных. Что происходит, когда частота митохондриальных мутаций возрастает? Исчезает выгода от отсроченного образования зародышевых клеток. Наша модель показывает, что в таком случае мутации накапливаются очень быстро и митохондрии поздно образовавшихся зародышевых клеток просто кишат мутациями. Как выразился генетик Джеймс Кроу, самый опасный источник мутаций в популяции – это плодовитые старички. К счастью, благодаря однородительскому наследованию, старички никак не могут передать потомству свои митохондрии. Мы учли повышенную частоту мутаций и обнаружили, что ген, вызывающий раннее обособление клеток зародышевой линии, в этом случае будет распространяться в популяции: раннее отделение зародышевой линии и погружение женских гамет в спячку спасают от накопления митохондриальных мутаций. Еще должны поддерживаться адаптации, снижающие уровень мутаций в клетках зародышевой линии. На самом деле, как показал мой коллега Джон Аллен, митохондрии в зародышевой линии самок, судя по всему, неактивны: они “выключаются” уже в момент отделения первичных половых клеток во время эмбрионального развития яичников. Джон Аллен давно утверждает, что митохондрии в яйцеклетках служат генетическими “шаблонами”, которые редко мутируют по той причине, что они неактивны. Наша модель это подтверждает, если рассматривать современных животных с коротким жизненным циклом и большим количеством митохондрий, которые часто мутируют, но не их предков – животных с длительным жизненным циклом, или более обширные группы, например растения, водоросли и протисты.
А это означает, что самого по себе разнообразия митохондриального состава достаточно, чтобы объяснить появление многоклеточных организмов с анизогамией (со сперматозоидами и яйцеклетками), однородительским наследованием и наличием зародышевой линии с обособлением женских гоноцитов на ранних стадиях развития – что в совокупности является основой всех различий между женским и мужским полом. Иными словами, наследование митохондрий позволяет объяснить большинство реальных физических различий между полами. Может быть, эгоистический конфликт между клетками также сыграл свою роль, хотя этого могло и не быть: возникновение разделения клеток на соматические клетки и клетки зародышевой линии можно объяснить и без эгоистического конфликта. Очень важно, что наша модель выявила порядок событий, который отличается от того, что я предположил в самом начале. Я думал, что однородительское наследование митохондрий было предковым состоянием, затем появились клетки зародышевой линии, а возникновение сперматозоидов и яйцеклеток связано с расхождением настоящих полов. Вместо этого наша модель показала, что предковым состоянием было двуродительское наследование, затем возникла анизогамия (наличие сперматозоидов и яйцеклеток), затем появилось однородительское наследование, и, наконец, возникли клетки зародышевой линии. Но верна ли эта пересмотренная хронология? В любом случае, мы слишком мало знаем, чтобы сказать наверняка. Однако это в явном виде сформулированное предсказание, которое можно проверить, и мы надеемся это сделать. В первую очередь мы займемся губками и кораллами. У обеих групп есть сперматозоиды и яйцеклетки, но отсутствует обособление клеток зародышевой линии. Разовьется ли оно у них, если мы будем производить отбор на повышение частоты митохондриальных мутаций?
Перед тем как подойти к завершению, рассмотрим еще несколько следствий. Что может заставить возрасти частоту митохондриальных мутаций? Ускоренный оборот клеток и белков вследствие увеличения физической активности. Накопление кислорода в океанах незадолго до “кембрийского взрыва” сделало возможным появление активно перемещающихся билатерально-симметричных животных. Их возросшая физическая активность могла увеличить частоту митохондриальных мутаций (сравнительная геномика позволяет это измерить), что, в свою очередь, должно было привести у таких животных к обособлению клеток зародышевой линии. Так появилась бессмертная зародышевая линия и бренное тело, то есть появилась смерть – запланированный и предопределенный финал. Клетки зародышевой линии бессмертны в том смысле, что они продолжают делиться вечно. Они не стареют и не умирают. В каждом поколении на ранних стадиях развития происходит обособление зародышевых клеток, которые дадут начало следующему поколению. Хотя отдельные гаметы могут быть повреждены, тот факт, что дети рождаются молодыми, означает, что зародышевые клетки сохранили свой потенциал бессмертия, как клетки губок, которые могут регенерировать из мелких кусочков. Когда специализированные зародышевые клетки обособились, все остальное тело можно использовать для других целей: оно уже не ограничено необходимостью нести бессмертные половые клетки. Мы можем наблюдать это на примере тканей, которые уже не могут регенерировать – например тканей мозга. Это одноразовая, смертная плоть. Продолжительность жизни таких тканей ограничена и зависит от того, сколько времени нужно организму для размножения. Это, в свою очередь, зависит от того, насколько быстро животное достигает половой зрелости, от скорости его развития и предполагаемой продолжительности его жизни. Здесь мы сталкиваемся с источником смерти: компромиссом между половым размножением и старением. Мы обсудим это в следующей главе.
Выше мы рассмотрели, как митохондрии повлияли на эукариотические клетки. Вернемся к главному вопросу: почему у эукариот появился целый набор общих признаков, которых нет ни у бактерий, ни у архей? Как мы убедились, прокариоты ограничены структурой собственных клеток, особенно потребностью в генах, регулирующих клеточное дыхание. Приобретение митохондрий преобразило адаптивный ландшафт эукариот, сделало возможным увеличение объема клеток и размера генома на 4–5 порядков. Первым импульсом к этому явился маловероятный, почти случайный эндосимбиоз двух прокариот, который, однако, повлек тяжелые и закономерные последствия. Тяжелые – потому, что клетка без ядра очень уязвима для атак чужеродных молекул ДНК и генетических паразитов (интронов) ее собственных эндосимбионтов. Закономерные – потому, что все ответные изменения клетки-хозяина (возникновение ядра, полового размножения, двух полов и зародышевой линии) можно объяснить в рамках классической эволюционной генетики, хотя и с нестандартной позиции. Некоторые из этих идей могут быть ошибочными, как моя гипотеза о происхождении двух полов. Но дальнейший разбор выявил то, что я и не предполагал – причину разделения соматических клеток и клеток зародышевой линии, истоки секса и смерти. Внутренняя логика этих процессов, выявленная путем точного моделирования, красива и предсказуема. Если где-то еще может возникнуть жизнь, она, вероятно, придет к сложности сходным путем.
Этот взгляд на всю четырехмиллиардную историю жизни на Земле отводит митохондриям центральную роль в эволюции эукариотических клеток. В последние годы в медицинских исследованиях общепринятой стала сходная точка зрения: сейчас известно, что митохондрии играют очень важную роль в программируемой клеточной смерти (апоптозе), развитии опухолей, дегенеративных заболеваний, бесплодия и много чего еще. Но некоторых ученых-медиков раздражает, когда я доказываю, что митохондрии – это краеугольный камень физиологии клетки. Меня упрекают в однобокости мышления. Ведь если рассмотреть человеческую клетку под микроскопом, вы увидите, что ее работу поддерживает множество подогнанных друг к другу частей, среди которых митохондрия лишь один из винтиков, хотя и важный. Но с эволюционной точки зрения это не так: митохондрия – один из двух равноправных участников, породивших сложно устроенную жизнь. Эукариотические черты – вся физиология клетки – возникли в результате “перетягивания каната” двумя этими участниками. Оно продолжается по сей день. В заключительной главе вы увидите, что это взаимодействие лежит в основе нашего здоровья, плодовитости и долголетия.
Назад: Глава 5 Появление сложных клеток
Дальше: Часть IV Предсказания

