Книга: Лекции о работе больших полушарий головного мозга
Назад: Лекция девятая
Дальше: Лекция одиннадцатая
Лекция десятая
Иррадиирование и концентрирование нервных процессов в коре больших полушарий: б) иррадиирование и концентрирование тормозного процесса по большим полушариям; в) иррадиирование и концентрирование раздражительного процесса
Мм. гг.! В прошлой лекции мы приступили к изучению движения нервных процессов в коре больших полушарий. Начав с внутреннего торможения, мы из опытов убедились в том, что оно, будучи произведено соответствующим раздражением в определенном пункте, сперва быстро иррадиирует по своему анализатору, а затем медленно концентрируется в направлении к исходному пункту, и что даже можно наблюдать это двустороннее движение шаг за шагом. В настоящей лекции мы будем продолжать следить за этим движением по всему полушарию, т. е. за переходом торможения из анализатора в анализатор. Опыты, сюда относящиеся, проделаны со всеми нашими случаями внутреннего торможения.
При описании дифференцировочного торможения было уже сообщено, что торможение, произведенное в одном анализаторе, обнаруживается и в других. В опытах над этим торможением (Белякова) оказалось, что при слабой степени его оно не проявляется в другом анализаторе, при сильном же его напряжении оно выступает и тут, но в гораздо меньшей мере, чем в исходном анализаторе.
Вот несколько из этого ряда опытов.
У собаки между прочими выработаны отдельные пищевые условные рефлексы: на звук гальтоновского свистка в 4000 колебаний в секунду и на бесшумное движение вертящегося перед глазами собаки предмета. Звуковой рефлекс обыкновенно составляют 11–12 капель за 30 секунд, зрительный – 7–8 капель. Звук на полтона ниже – дифференцировка.
Опыт 8.VI.1911

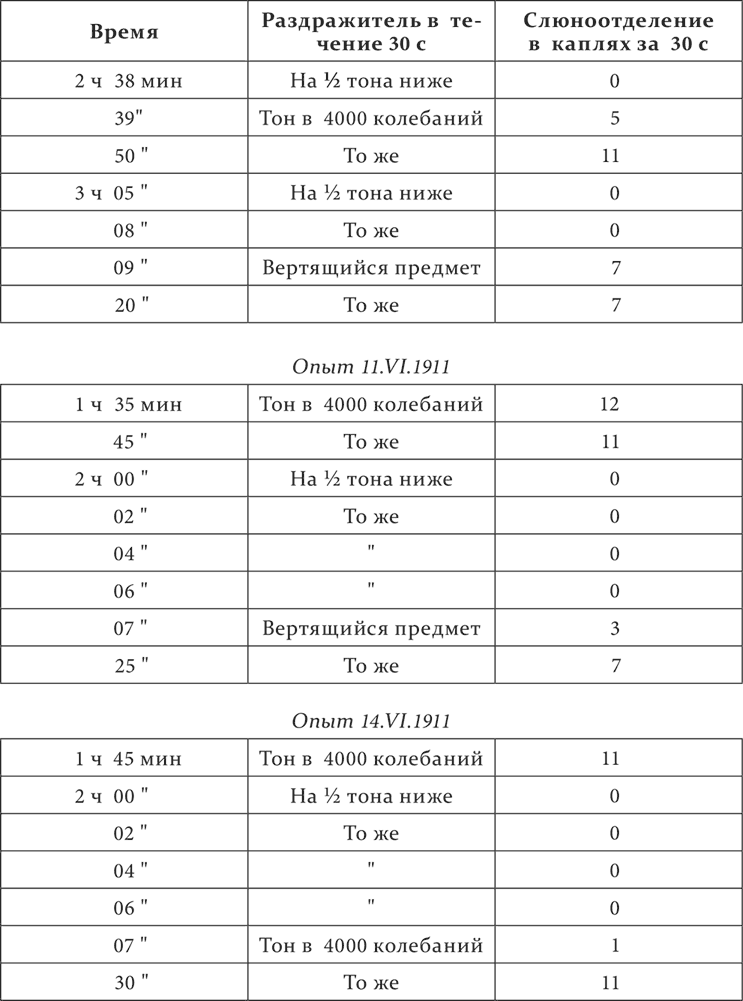
Смысл опытов ясный. После двукратного применения подряд дифференцированного тона и минуту спустя после этого положительный тон теряет в его действии больше половины, зрительный же раздражитель не терпит ни малейшего ущерба. После большего суммирования торможения, после четырехкратного применения (да еще при меньшей паузе) дифференцированного тона, и также минуту спустя, оказывается заторможенным и зрительный рефлекс, но только немного больше, чем в два раза, между тем как в тех же условиях звуковой рефлекс задерживается почти полностью.
Совершенно то же самое получается при угасательном торможении, как показывают опыты Э. Л. Горна. Здесь благодаря некоторой вариации опыта еще убедительнее выступает факт движения тормозного процесса, что касается специально более раннего ухода его из другого анализатора. Вот некоторые из этих опытов.
У собаки образованы между прочими отдельные пищевые условные рефлексы на тон cis духового камертона и на вспыхивание перед глазами собаки трех шестнадцатисвечных лампочек.
Опыт 15.XII.1911
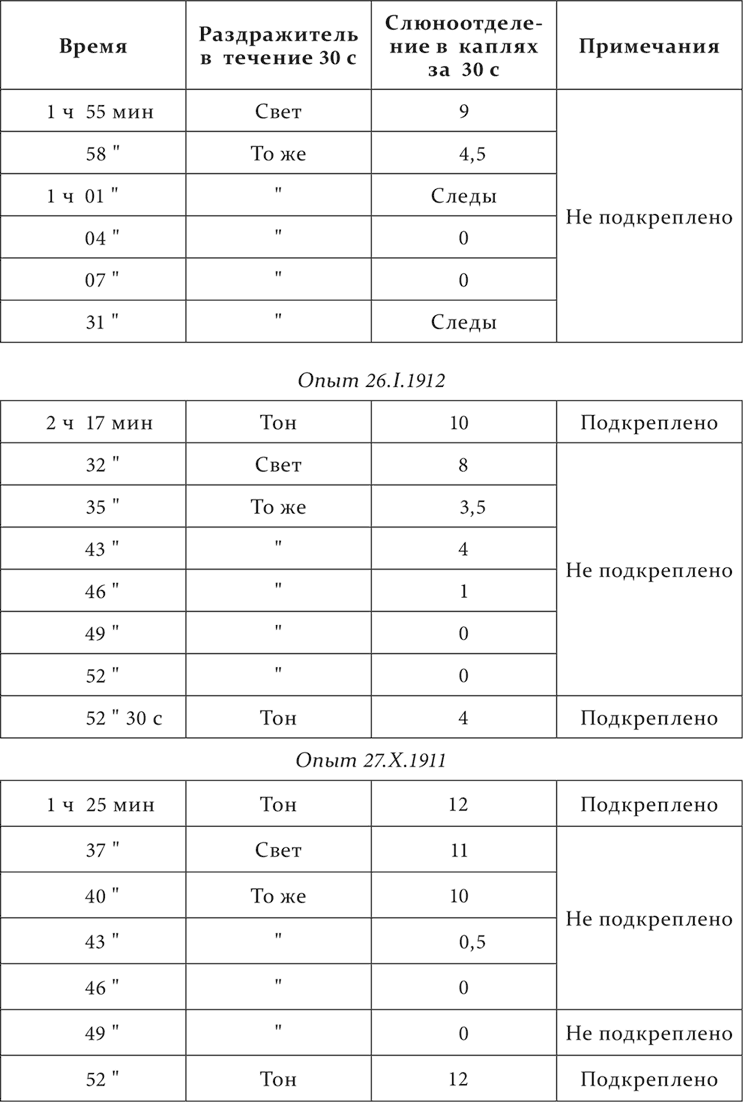
Угашался рефлекс на свет и испытывалось вторичное действие на тоновой рефлекс. Световой рефлекс, угашенный до степени двух нулей и предоставленный себе, начал восстанавливаться лишь через 24 минуты. Тоновой рефлекс, после такой же степени угашения светового рефлекса примененный непосредственно без всякого промежутка, потерял в своем эффекте только около половины, а испытанный в другом опыте, через 3 минуты оказался уже совершенно свободным от торможения. Следовательно, тормозной процесс из светового анализатора распространился на звуковой только в слабой мере и скорее ушел оттуда совсем. Что он ушел оттуда скорее не потому, что был менее значительным, доказывается дальнейшими подробностями опытов. В световом анализаторе промежуток между повторениями (3 минуты) на всех степенях угасания постоянно влек за собой только падение рефлекса, а в звуковом тот же промежуток повел к полному восстановлению рефлекса. Кроме того, во втором из приведенных опытов при угашении светового рефлекса, когда этот рефлекс представлял уже небольшую величину, даже втрое экстренно увеличенный промежуток только остановил дальнейшее уменьшение рефлекса, но не обусловил сколько-нибудь значительного восстановления его. Что, наконец, разница в восстановлении первично и вторично угашаемых рефлексов не зависела в данном случае от особенностей световых и звуковых клеток, свидетельствуют другие наши опыты, где первично угашался тоновой рефлекс, а вторичное угашение наблюдалось на световом, и где, однако, отношения остались совершенно теми же. Ввиду полного тождества этих и уже приведенных опытов, я их не привожу. Таким образом, можно с правом принимать, что во вторично угашаемом анализаторе мы имеем дело с периферией иррадиирующего процесса и что с нее начинается обратное движение процесса, концентрирование его, совершенно так, как это мы наблюдали внутри кожного анализатора.
«Одна и та же степень торможения не сказывается на сильных раздражителях и отчетливо дает себя знать на слабых»
Опыты с движением угасательного торможения по целому полушарию вообще нами очень разнообразились. Мы, имея в первичном угашаемом анализаторе два разных раздражителя (например вспыхивание лампочек и верчение предмета, стоящего перед глазами животного) и угашая один из них, следили за торможением как в другом анализаторе, так и в том же анализаторе на обоих раздражителях. Порядок освобождения от иррадиирующего торможения был таков: скорее всего освобождался рефлекс из другого анализатора; близко к нему по времени, но несколько медленнее – вторично угашаемый рефлекс из первично угашаемого анализатора и, наконец, сильно запаздывая – первично угашенный рефлекс. Последнее, т. е. разница между рефлексами, принадлежащими к одному и тому же анализатору, очевидно, указывает на территориальное разграничение в нем различных рефлексов, с чем мы уже познакомились в предшествующей лекции.
Те же результаты дали опыты и над движением условного торможения. Один и тот же внешний агент был сделан условным тормозом нескольких условных раздражителей, принадлежащих к разным анализаторам, т. е. комбинация его с каждым из них отдельно повторялась без сопровождения безусловным раздражителем и таким образом превращалась в тормозную. Опыт исполнялся таким образом. Испытывался один из этих условных раздражителей и отмечался его слюноотделительный эффект. Затем несколько раз повторялась тормозная комбинация или с этим раздражителем, или с условным раздражителем из другого анализатора. Опыт кончался вторичной пробой первого раздражителя через разные промежутки времени после последнего повторения тормозной комбинации. Если тормозная комбинация включала первый раздражитель, то он теперь последовательно оказывался сильно заторможенным и восстановлялся медленно до его исходной величины. Если же в состав тормозной комбинации входил раздражитель из другого анализатора, то испытанный вначале раздражитель был заторможен только в слабой степени и быстро возвращался к его первоначальной величине. Это значило, что последовательное торможение в анализаторе условно-тормозного раздражителя было очень значительно и долго в нем держалось, в другой же анализатор оно проникло только в малой мере и скоро оттуда ушло.
«Чем тоньше дифференцировка, тем при полной дифференцировке на нее должен развиться более интенсивный тормозной процесс»
Вот опыты из работ В. А. Дегтяревой.
У собаки отдельные пищевые условные рефлексы на удары метронома (М) и вспыхивание электрических лампочек (А). Условный тормоз к тому и другому – бесшумное верчение стоящего перед собакой предмета (В). И условные раздражители, и тормозная комбинация продолжаются каждый раз минуту, также и слюноотделение отмечается в каплях за минуту.
Опыт 13.V.1913
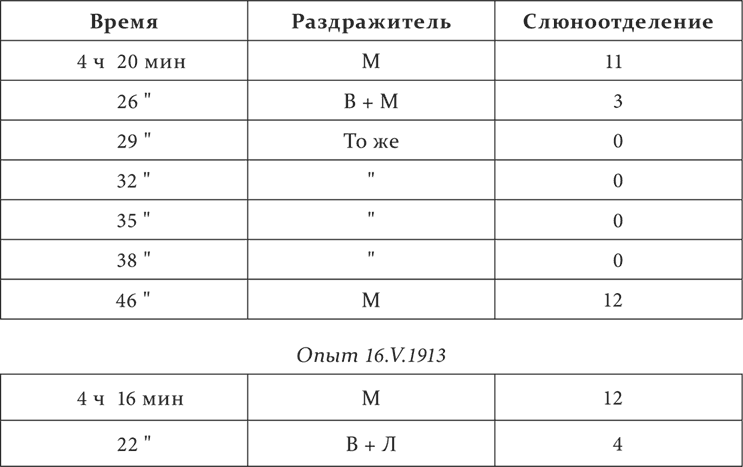

Метрономный рефлекс после пятикратного применения тормозной комбинации, в которой он сам участвовал, восстановился вполне только 7 минут спустя; после же комбинации без него он был почти полный уже только через 15 секунд.
Наконец, нами было исследовано движение торможения, которое развивается при запаздывании положительного эффекта в сильно отставленных рефлексах, предшествуя деятельной фазе. Соответственно особой постановке торможения, как фазовому явлению, надо было ожидать некоторых особенностей в опытах над его движением, что и оказалось. Между разными собаками наблюдалась большая разница в результате наших испытаний. У некоторых животных удавалось легко показать распространение этого торможения в другие анализаторы. Делалось это таким образом. Раздражитель запаздывающего рефлекса прекращался во время недеятельной фазы, и непосредственно или спустя некоторое время пускался в ход условный раздражитель из другого анализатора, предварительно испробованный на его величину. Вот протоколы таких опытов (Горна).
У данной собаки имеются между прочими следующие кислотные условные рефлексы: отставленные на 30 секунд – один на вспыхивание электрических лампочек, другой на кожно-механическое раздражение, и отставленный на 3 минуты на удары метронома.
Опыт 26.I.1912
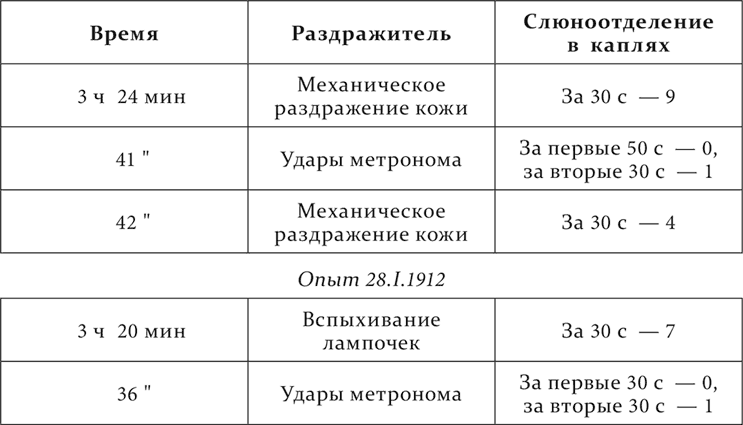

Результат вполне резкий. Процесс торможения, вызываемый метрономом в начале его действия, проник из звукового анализатора как в зрительный, так и в кожный анализаторы и сказался в уменьшении их рефлексов в 2–3 раза.
Но это у одной собаки. У другой результат был скорее обратный. Условные раздражители, примененные непосредственно после недеятельной фазы отставленного рефлекса, даже несколько увеличились в своем размере. Причина этой разницы была ясна. Когда у первой собаки между изолированной недеятельной фазой и пробой другого рефлекса ввели промежуток в 2 минуты, то в течение этих 2 минут деятельная фаза не наступила, т. е. тормозная фаза действительно была точно выделена, и поэтому последовательное торможение на другом рефлексе было налицо. У второй же собаки при тех же условиях в промежутке обнаружилась деятельная фаза, и, естественно, другой рефлекс был не уменьшенным, а увеличенным. Следовательно, у этой собаки ранее и энергичнее выступил раздражительный процесс, т. е. недеятельная фаза не была полностью изолирована. С этим совпадало и то, что у первой собаки тормозной процесс вообще заметно преобладал сравнительно с раздражительным.
В прошлой и настоящей лекциях я привел в изобилии опыты, иллюстрирующие движение тормозного процесса по массе больших полушарий. Вам могло показаться, что это было даже и излишне. Но я сделал это с умыслом, чтобы сделать для вас очевидным, как много раз мы наблюдали это явление в более или менее простом, так сказать, в чистом виде. И все же это был только, если можно так выразиться, основной скелет явления, выступавший перед нами благодаря или особенности экспериментирования, или индивидуальным особенностям животных, скелет, однако, наиболее часто покрываемый добавочными элементами, очень усложняющими, так сказать, полную действительность.
Первое, на что мы уже давно натолкнулись в этих опытах, состояло в следующем.
В описанных в прошлой лекции опытах (Красногорского) об иррадиировании и концентрировании тормозного процесса в кожном анализаторе встретился такой неожиданный тогда для нас факт. При предварительной пробе в опыте условное механическое раздражение места кожи, удаленного от тормозного пункта на 22 см, дало 6 капель слюны за 30 секунд, а раздражение зрительного анализатора дало 5 капель. Производится троекратное раздражение тормозного пункта. Минуту спустя после последнего раздражения испытывается зрительное раздражение – нуль слюноотделения, т. е. полное последовательное торможение. Через 10 минут применяется теперь четырехкратное раздражение тормозного пункта. Через минуту после него раздражение места, удаленного на 22 см, сопровождается полным эффектом – 8 каплями.
Подобный же факт наблюдался в других наших опытах (Чеботаревой). У собаки имелось несколько условных рефлексов на раздражители из разных анализаторов. Условный метрономный раздражитель, примененный вместе с условным тормозом (из зрительного анализатора), не давал ни капли слюны. 1–2 минуты спустя после того метроном один вполне восстановлял свой обычный слюноотделительный эффект и был при этом подкрепляем, а испытанные вскоре после того условные рефлексы на механическое раздражение кожи и на запах камфоры оказывались еще значительно заторможенными.
Таким образом, выходило, что торможение через некоторый срок, после того как оно было произведено в данном анализаторе, уже исчезало из него, между тем в других анализаторах даже через большие сроки оно еще давало себя знать.
Но вместе с тем было обращено внимание на то, что это случалось тогда, когда условные рефлексы из других анализаторов были очень молодые, т. е. только что образованные, или хотя и старые, но давно не практиковавшиеся, или, наконец, вообще слабые, выработанные на агентов малой интенсивности. Ввиду этого можно было себе представить, что когда известный пункт, ранее заторможенный, оказывался свободным от торможения, это еще не значило, что в его районе не было более торможения. В этом пункте раздражительный процесс в силу определенных условий мог брать перевес над тормозным процессом, делаться, так сказать, иммунным по отношению к окружающему его, но уже ослабленному тормозному процессу. И действительно, это постоянный факт, что рефлексы с еще недостаточно концентрированным или вообще со слабым раздражительным процессом чрезвычайно легко подвергаются действию как внешнего, так и внутреннего торможения, между тем как сильные рефлексы остаются вне влияния тех же торможений. Об этой, так сказать, крепости или даже неприкосновенности сильных и хорошо выработанных как раздражительных, так и тормозных пунктов будет речь еще впереди.
«При взаимодействии условных рефлексов, образованных на различные пищевые вещества, торможение одних другими наступает не сразу, а спустя несколько минут»
В свое время, когда впервые наблюдались случаи, как бы противоречащие правилу иррадиирования и концентрирования торможения, были исполнены следующие нарочитые опыты (А. М. Павловой).
У собак были выработаны условные рефлексы на раздражители из разных анализаторов, причем обращалось тщательное внимание на то, чтобы все они повторялись одинаковое число раз. Когда все они достигают максимума эффекта, с одним из них комбинируется новый внешний агент, и эта комбинация обычным методом превращается в тормозную. Тормозная комбинация, а после нее ее условный раздражитель повторяются до тех пор, пока последовательное торможение комбинацией положительного раздражителя не ограничится 2 минутами, т. е. после 2 минут уже незаметно на нем. Конечно, и за этот период соблюдается требование об одинаковой практике всех рефлексов. Затем все остальные рефлексы впервые испытываются совместно с условным тормозом. Оказывается, что и эти рефлексы, за исключением одного, тоже освобождаются от торможения к концу 2 минут после применения тормозной комбинации. Исключение приходится на условный рефлекс из слабого раздражения светом, который обычно отстает по слюноотделительному эффекту от остальных рефлексов. Отсюда следует, что одна и та же степень торможения не сказывается на сильных раздражителях и отчетливо дает себя знать на слабых.
Но это только один элемент, осложняющий факт иррадиирования и концентрирования тормозного процесса. Впоследствии и постепенно мы познакомились и с другим, имеющим гораздо большее значение и еще более нарушающим простой ход иррадиирования и концентрирования. Об этом элементе мы сообщим имеющийся у нас материал в следующей лекции.
«Внешнее раздражение, пришедшее в кору с определенного пункта или района рецептора, не остается в соответствующих им точке или отделе коры, а иррадиирует дальше по коре, и в тем более слабой степени, чем дальше отходит от центра раздражения»
А теперь я перехожу к нашим опытам относительно движения раздражительного процесса. Этот вопрос обработан у нас в гораздо меньшей степени. Наш первый опыт (М. К. Петровой) с иррадиированием раздражительного процесса исполнен совершенно по типу тех опытов, которыми мы начали изучение иррадиирования и концентрирования тормозного процесса. У собак вдоль задней конечности, начиная с плюсны до таза, были расположены пять приборчиков для механического раздражения кожи, приблизительно на равных друг от друга расстояниях. Из действия самого нижнего приборчика (№ 1) у одной собаки сделан условный кислотный рефлекс, у другой – пищевой. Верхние приборчики (№№ 2, 3, 4 и 5) в силу генерализации, конечно, вызывали также соответствующие рефлексы, но они обычным приемом были отдифференцированы и доведены все одинаково до нуля эффекта. Тогда было приступлено к главной задаче опытов. Сперва испытывалось действие самого нижнего приборчика, т. е. положительный раздражитель в течение 30 секунд, причем слюноотделение за каждые 15 секунд отмечалось отдельно, а затем, как обыкновенно, присоединялся безусловный раздражитель. Спустя некоторое время нижний приборчик опять пускался в ход, но только на 15 секунд, а затем сейчас же после него начинали действовать, и тоже в течение только 15 секунд, верхние приборчики, то один, то другой в разных опытах, то более близкий, то более дальний. Получилось следующее. При смене № 1 на № 2, ближайший, слюноотделение в течение действия этого приборчика было обыкновенно много меньше, а иногда и равно слюноотделению, наблюдавшемуся в течение 15 секунд действия приборчика № 1. При смене таким же образом № 1 на № 5, т. е. самого дальнего, слюноотделение при нем резко падало. Приборчики № 3 и № 4, примененные в тождественной обстановке, дали средние цифры, приближаясь № 3 к № 2 и № 4 к № 5. В другом ряде опытов отмечалось слюноотделение не только за время действия тормозных раздражителей, но до конца отделительного периода. Тогда разница в действии близких и дальних приборчиков выступала почти без единого исключения и гораздо рельефнее. Отделительный период продолжается дольше и был гораздо значительнее по величине после действия близкого приборчика, чем дальнего.
Привожу данные ряда опытных дней.
У этой собаки кислотный условный рефлекс.
Опыт 14.XI.1913
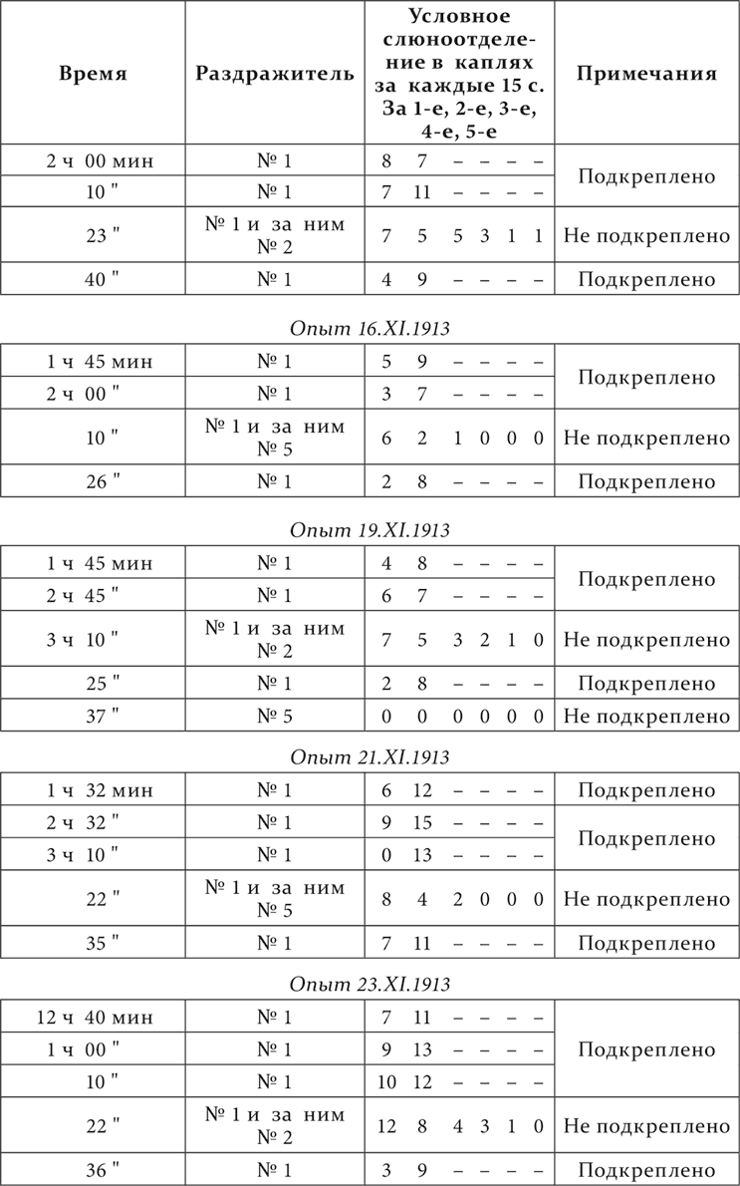
Опыт 28.XI.1913
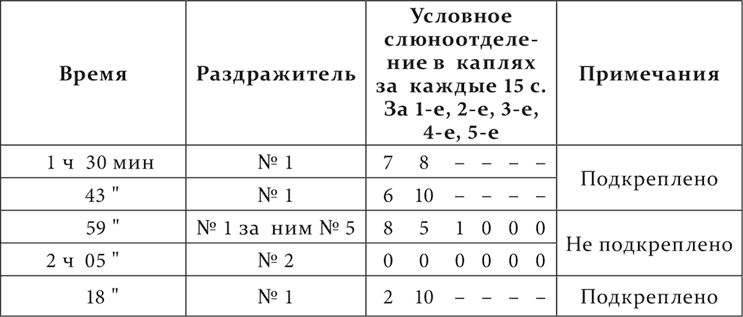
Результат опытов очень резкий. При смене приборчика № 1 на приборчик № 2 отделение слюны во время действия (15 секунд) первого приборчика, соответственно приведенному ряду опытов: 7, 7 и 12 капель, а отделение во время действия (15 секунд) второго вместе с последовательным отделением: 15, 11 и 16; при смене же приборчика № 1 на приборчик № 5 отделение во время действия первого: 6, 8 и 8 капель, а при действии второго вместе с последовательным отделением: 3, 6 и 6 капель. Там резко больше, здесь отчетливо меньше. После действия приборчика № 2 последовательное слюноотделение продолжается 45–60 секунд, после приборчика № 5 – только 15 секунд. А постоянное тормозящее действие приборчика № 2 должно быть и есть больше, чем приборчика № 5, потому что, как мы уже знаем, чем тоньше дифференцировка, тем при полной дифференцировке на нее должен развиться более интенсивный тормозной процесс. Что действительно тормозящее действие от приборчика № 2 было больше, чем действие от приборчика № 5, в данной работе было констатировано нарочитыми опытами с последовательным торможением и от одного и от другого приборчика, притом как на приборчике № 1, так и на условных раздражителях из других анализаторов. Привожу пример последовательного торможения на приборчике № 1 через 5 минут после действия тормозящих приборчиков. Опыт на той же собаке.
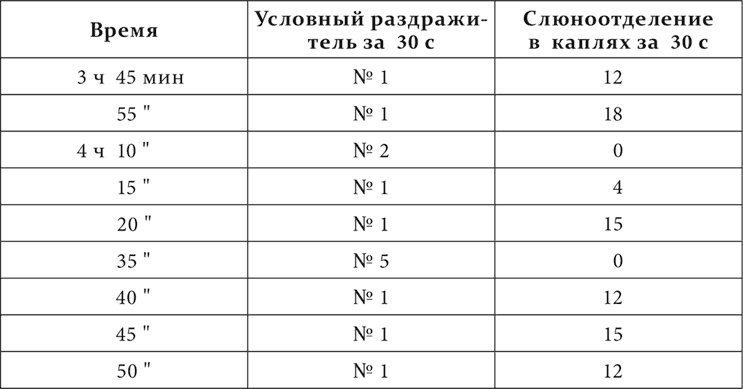
Торможение от № 2 через 5 минут его действия еще очень сильное, от № 5 через тот же срок почти или совсем не заметно.
После этого результат ряда приведенных опытов может быть истолкован только так, что процесс раздражения, вызванный действием приборчика № 1, на близком месте, соответствующем приборчику № 2, дает себя знать больше, чем на месте более дальнем, соответствующем приборчику № 5; другими словами, он распространился, иррадиировал из места его возникновения в ближайшие к нему пункты.
Эти опыты еще раз повторялись в последнее время (Подкопаевым). На одной стороне тела с передней лапы через все туловище до конца задней лапы располагаются приборчики для механического раздражения кожи. Действие прибора на передней лапе при помощи обычной процедуры становится положительным пищевым условным раздражением, действие всех остальных – тормозным. Отделение слюны отмечается каждые 5 секунд. Один раз положительный раздражитель действует до подкрепления все 30 секунд, другой раз только первые 15 секунд, затем следует пауза в 15 секунд, только после которой и производится подкрепление. Таким образом, мы имели перед собой ход отделения слюны при сплошном раздражении и при прерываемом на 15 секунд. При сплошном величина слюноотделения за первые 15 секунд относилась к величине его за вторые 15 секунд как 1:2,4; при прерываемом – как 1:1,25. В дальнейших опытах положительный раздражитель после пятнадцатисекундного действия сменялся на тормозной раздражитель тоже пятнадцатисекундного продолжения то при помощи ближайшего приборчика, то самого дальнего. Теперь отношение величины слюноотделения за первые 15 секунд к величине за вторые в первом случае было как 1:1,35, а во втором как 1:0,53.
«Факт первоначальной генерализации всякого только что выработанного условного рефлекса: как бы самопроизвольное действие в качестве условных раздражителей таких агентов, которые сами по себе ранее не применялись, но составляют родственную группу с тем агентом, на который при помощи определенной процедуры был выработан данный условный рефлекс»
Очевидно, и тут, как в опытах предшествующего сотрудника, через 0 секунд (практически через 0,5–1 секунды) по прекращении положительного раздражения раздражительный процесс обнаруживается на ближнем тормозном пункте, уничтожая его действие, а на дальнем его нет.
Как в том, так и в этом ряде опытов мы не имеем возможности сказать, какая это фаза движения раздражителя – фаза иррадиирования или концентрирования? идет процесс вперед или уже возвращается назад? Сейчас предпринимаются опыты, в которых тормозные раздражители применяются не сейчас же по прекращении положительного раздражителя, а спустя то или другое число секунд.
К сожалению, относительно этого вопроса приведенным исчерпывается наш теперешний нарочитый опытный материал, но у нас имеется случайно наблюденный материал, очевидно или по всей вероятности имеющий ближайшее отношение к тому же предмету.
Вот эти отдельные случаи.
В нашем распоряжении была собака с чрезвычайно резко выраженным сторожевым рефлексом. Постоянно занимающийся с ней экспериментатор (М. Я. Безбокая), находясь с ней в отдельной комнате, с полным удобством снаряжал ее к опыту, ставил в станок и укреплял на ней разные приборы. С условными рефлексами дело шло вполне удовлетворительно. Но появление в экспериментальной комнате постороннего лица вызывало сильнейшую агрессивную реакцию со стороны животного, тем более если постороннее лицо (в таком случае был я) заменяло экспериментатора перед собакой. Когда я при таком положении дела применял выработанный пищевой условный раздражитель, то слюноотделительный эффект его был значительно больше обычной величины при работе с собакой ее постоянного экспериментатора, и собака ела предлагаемую пищу с чрезвычайным мускульным напряжением. Но агрессивная реакция в отношении меня совершенно прекращалась, когда я, намеренно, что называется, не шелохнулся, не делал ни малейшего движения. Из всего рефлекса на меня оставалось только постоянное фиксирование меня глазами. И теперь тот же условный раздражитель давал меньше слюны, чем в норме, или даже совсем оставался без секреторного эффекта. Но стоило мне держаться свободнее и особенно встать, чтобы сейчас же возвращались и агрессивная реакция и увеличенное слюноотделение. Интересно дальше следующее. Если в присутствии постороннего лица, сидевшего неподвижно, собака успокаивалась, то подкармливание собаки вслед за условным раздражителем вызывало на некоторое время после еды агрессивную реакцию на это лицо, хотя оно продолжало сидеть неподвижно. Едва ли эти факты можно принимать иначе, как только так. Сильное раздражение, производимое видом постороннего, делающего движения лица, иррадиировало по мозгу и повысило, между прочим, возбудимость и его района, относящегося к акту еды. Когда же это раздражение при уменьшении внешнего раздражителя ослабло и концентрировалось, то, наоборот, возбудимость других отделов уменьшилась сравнительно с покойным состоянием (наше внешнее торможение). Такое же влияние в свою очередь имело и раздражение едой, повысив на короткое время возбудимость центра агрессивной реакции.
«Внешнее раздражение, пришедшее в кору с определенного пункта или района рецептора, не остается в соответствующих им точке или отделе коры, а иррадиирует дальше по коре, и в тем более слабой степени, чем дальше отходит от центра раздражения»
Подобное же явление мы наблюдали на другой совершенно исключительной собаке (в работе И. Р. Пророкова). У нее механическое раздражение кожи вызывало особенную и в высшей степени резкую двигательную реакцию (может быть, полового характера или что-либо аналогичное эффекту щекотания). Несмотря на это, на механическое раздражение кожи образовался вместе с тем и условный пищевой рефлекс. Этот пищевой рефлекс, в ярком контрасте с обычным отношением, был по слюноотделению самым большим из всех остальных условных рефлексов. В данном случае обыкновенно при начале раздражения кожи появлялась особенная двигательная реакция, а 10–15 секунд спустя она сменялась энергичнейшей пищевой двигательной реакцией, какой не было при других условных раздражителях. Когда мы некоторыми приемами, о чем интересная речь будет впереди, устранили странную реакцию, то кожно-механический условный рефлекс занял по своей величине обычное место, уступая первенство звуковым раздражителям. Тоже очевидный случай иррадиации раздражительного процесса.
Таким же образом мы склонны понимать и следующий факт. При взаимодействии условных рефлексов, образованных на различные пищевые вещества, торможение одних другими наступает не сразу, а спустя несколько минут. Можно думать, что сначала имеет место иррадиация из непосредственно данным веществом раздражаемого пункта химического анализатора на соседние, чем временно поднимается их возбудимость, и лишь потом, когда раздражение в первоначальном пункте сконцентрируется, наступает торможение (опыты Савича).
Генерализация происходит, развивается в течение некоторого времени, потому что более отдаленные рефлексы, как более слабые, обнаруживаются позже»
С той же точки зрения можно бы объяснить и неожиданный факт, встретившийся в работе П. Н. Васильева при образовании разных температурных условных рефлексов. На раздражение кожи холодом около 0 °C был выработан условный кислотный рефлекс, а на тепловое раздражение около 47 °C – пищевой. Потребовалось большое время для образования этих рефлексов. А когда они, наконец, были готовы, они оказались в высшей степени неустойчивыми в следующем отношении. Стоило несколько раз подряд повторить и подкрепить условный кислотный холодовой рефлекс, чтобы примененный после него и хорошо выработанный пищевой тепловой рефлекс превратился как бы в холодовой, и наоборот. Факт превращения был вполне отчетлив и бесспорен. Натура рефлекса определялась, во-первых, двигательной реакцией, конечно, совершенно различной при этих обоих рефлексах: при пищевой собака повертывалась в сторону экспериментатора, пристально смотря то на него, то туда, откуда появлялась еда, и облизывалась; при кислотной отворачивалась от экспериментатора, фыркала, скулила, трясла головой и делала языком выбрасывающие движения. Во-вторых, за это говорил химический характер слюны из подчелюстной слюнной железы: сильно слизистой, густой при пищевом рефлексе и жидкой, водянистой при кислотном. Мы проследили эти разницы особенно тщательно. Факт такого упорного и долгого превращения казался странным. Его было можно бы понять следующим образом.
Как известно, периферические концы холодового и теплового анализаторов распределены на поверхности кожи точечно и смешанно. Можно было бы предполагать, что и центральные концы этих температурных анализаторов также тесно переплетены между собой, и из наших опытов (А. А. Шишло) мы действительно знаем, что корковые концы холодовых и тепловых нервов топографически совпадают. Если принять это во внимание, то будет понятно, почему раздражение так легко иррадиирует из холодового анализатора в тепловой, и обратно, почему так трудно достигается в наших опытах разъединение деятельности обоих анализаторов.
Теперь я должен вернуться к факту, который был уже упомянут в первой лекции об анализаторной деятельности коры и который по его механизму должен быть помещен в настоящем отделе нашего изложения. Это факт первоначальной генерализации всякого только что выработанного условного рефлекса, т. е. факт как бы самопроизвольного действия в качестве условных раздражителей таких агентов, которые сами по себе ранее не применялись, но составляют родственную группу с тем агентом, на который при помощи определенной процедуры был выработан данный условный рефлекс.
Для образца я приведу опыты над кожным анализатором (Анрепа). На одной половине тела были расположены приборчики для механического раздражения кожи. Действие приборчика № 0, находящегося на бедре, было сделано условным пищевым раздражителем. Другие приборчики были распределены таким образом: № 1 – на конце задней конечности, № 2 – на тазу, № 3 – на середине туловища, № 4 – на плече, № 5 – на предплечье и № 6 – на конце передней конечности. Действие всех этих приборчиков само по себе, т. е. без всякой предварительной выработки, также сопровождалось условным слюноотделением, причем слюноотделительный эффект чрезвычайно постепенно падал по мере удаления от № 0. В нижеприводимой табличке содержатся средние цифры, полученные на одной из трех примененных для этих опытов собак. Количества слюны выражены в делениях шкалы, где деление соответствует 0,01 см³. Отделение отмечалось за 30 секунд действия раздражителей.
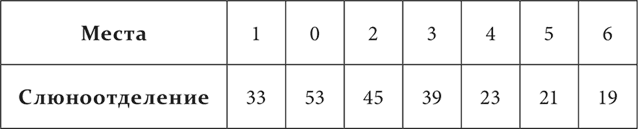
Стойкость генерализации очень различна у разных собак, и удерживание ее при опытах на определенной степени не всегда легко. Если мы пробуем новые места и всякий раз раздражение подкрепляем, мы выравниваем рефлексы. Если же пробные раздражения оставляем без подкрепления, мы даем повод к развитию торможения, к образованию дифференцировок. Поэтому в этих опытах, чтобы иметь более или менее точный результат, гарантированный повторением, при повторных пробах строго поочередно один раз раздражение подкрепляют, другой раз нет, или же делают эти пробы через очень большие промежутки времени.
Как понимать механизм генерализации? Всего соответственнее нашим теперешним знаниям представлять себе это явление следующим образом. Каждому элементу рецепторного аппарата отвечают свое афферентное нервное волокно и своя нервная клетка в коре полушарий, и каждой большей или меньшей группе их – свои группы волокон и клеток. Внешнее раздражение, пришедшее в кору с определенного пункта или района рецептора, не остается в соответствующих им точке или отделе коры, а иррадиирует дальше по коре, и в тем более слабой степени, чем дальше отходит от центра раздражения, что мы воочию видели на другом нервном процессе, на торможении. Таким образом, рядом с первым пунктом раздражения и в силу иррадиации раздражаемые пункты также должны связаться с центром безусловного рефлекса и повести, следовательно, к образованию прибавочных рефлексов, тем сравнительно более слабых, чем дальше эти пункты находятся от пункта приложения внешнего раздражителя, так как величина условного эффекта определенно связывается с силой раздражения. С такой точкой зрения согласуется и наблюденный (П. С. Купаловым) факт, что генерализация происходит, развивается в течение некоторого времени, потому что более отдаленные рефлексы, как более слабые, обнаруживаются позже.
«Каждому элементу рецепторного аппарата отвечают свое афферентное нервное волокно и своя нервная клетка в коре полушарий, и каждой большей или меньшей группе их – свои группы волокон и клеток»
Объяснение, данное нами для факта первоначальной генерализации условных рефлексов в одном анализаторе, можно было бы применить и к факту универсальной генерализации, наступающей при следовых рефлексах, образованных на поздних следах раздражителя, и о которой было сообщено в седьмой лекции. При позднем присоединении безусловного раздражителя к данному внешнему агенту раздражение от него успевает расплыться по всему полушарию, и потому все пункты полушария могут войти в связь с центром безусловного раздражителя. При раннем же присоединении безусловного раздражителя центр его концентрирует раздражение по направлению к себе и тем ограничивает его слишком обширную иррадиацию.

Академик И. П. Павлов в докторской мантии на церемонии присуждения ему ученой степени доктора Кембриджского университета во время празднования 250-летия Лондонского королевского общества (Великобритания, 1912)
Назад: Лекция девятая
Дальше: Лекция одиннадцатая

