Книга: Достающее звено. Книга 2. Люди
Назад: Пролог
Дальше: Глава 2 Человекообезьяны: грацильные австралопитеки
Глава 1
Обезьянолюди: ранние австралопитеки
Ранние австралопитеки населяли Африку между 7 и 3,9 млн лет назад. В конце миоцена леса исчезали, саванны наступали, и вот под барабанную дробь, звонкие литавры и голосистые фанфары на сцену выходит…
Сахелянтроп Sahelanthropus tchadensis! И не беда, что у него голова с кулак, ничего, что больше всего он похож на карликовую гориллу, – это действительно он, Великий Предок!
Череп и нижняя челюсть сахелянтропа были найдены французскими исследователями в Северной Африке, в Республике Чад, в Торос-Меналла, и описаны в 2002 году (Brunet et al., 2002); позже добавились еще две нижние челюсти и новый зуб (Brunet et al., 2005). Это древнейший и примитивнейший представитель нашей родной эволюционной линии. По фауне отложения с останками сахелянтропа датированы 6–7 млн лет назад; к сожалению, фауна не меняется быстро, а вулканов, по пеплу коих можно было сделать точную датировку, поблизости не оказалось, так что поначалу приходилось довольствоваться погрешностью в миллион лет. Однако прогресс идет, и уже скоро по соотношению изотопов бериллия в костях антракотериев была получена на полмиллиона лет более точная дата – 6,8–7,2 млн лет назад (Lebatard et al., 2008).

Рис. 2. Череп Sahelanthropus tchadensis.
Череп получил коллекционный номер TM 266-01-60-1, но этот скучный паровозный код, конечно, не мог в должной мере отразить ликование антропологов, которые по сложившейся традиции окрестили его личным именем Тумай. Нам безмерно повезло, что от древнейшего гоминида сохранился целый череп, хотя бы и сильно перекошенный; деформацию же выправили трудолюбивые реставраторы (Zollikofer et al., 2005).
Кстати, об именах…
Антропологи тоже любят украшать себе жизнь. Так среди австралопитеков и “ранних Homo” появились “Беби из Таунга”, “Люси”, “Сын Люси”, целых два “ребенка Люси” (одна из них – “Дочка Люси”, или “Селам”), “Первое семейство”, “Миссис Плез” и “Мистер Плез”, “Тумай” и “Арди”, “Миллениум Мен”, или “Человек Тысячелетия”, “Кадануумуу”, или “Большой Человек”, “Загадочный череп” и “Черный череп” (он же “череп Дарта Вейдера”), “Абель”, “Джордж” и “малыш Джонни”, “Твигги”, “Синди”, “Орфей” и “Эвридика”, “Щелкунчик” и даже такие почти индейские имена, как “Колено Джохансона” и “Маленькая Стопа” (он же “Синдерелла”).
Череп Тумая совсем маленький, с объемом мозга всего 360–370 см³ – меньше, чем в среднем у обыкновенного шимпанзе, но чуть больше, чем у бонобо. Лоб невероятно плоский, череп сверху почти совсем ровный: по этому признаку сахелянтроп выглядит примитивнее любой человекообразной обезьяны. Получается так потому, что лицо сахелянтропа расположено впереди от мозговой коробки, а не под ней, как это обычно для человекообразных. Рельеф черепа не по размеру мощный: надбровье выступает хотя и не столь резко, как у шимпанзе, но фактически рекордно для австралопитековых; височные линии ближе к затылку сливаются в сагиттальный гребень; выйный гребень так вообще выглядит как широкая пластина, идущая от височных костей поперек затылка, заостренная по краю и загнутая вниз. Рельефом сахелянтроп однозначно превосходит шимпанзе, что загадочно. Возможно несколько объяснений. Во-первых, сахелянтроп мог питаться гораздо более жесткой пищей. Во-вторых, переход к прямохождению мог сопровождаться усилением шейной мускулатуры, так как голова держалась на позвоночнике наискосок, ее надо было уравновешивать сзади, рычагом для чего и служил выдающийся затылочный гребень.
Одно из самых замечательных свойств сахелянтропа – положение большого затылочного отверстия. Как уже говорилось, у четвероногих обезьян оно расположено сзади и ориентировано назад, а у человека – сдвинуто на середину основания черепа и смотрит вниз. У сахелянтропа же его положение и ориентация строго промежуточны. Видимо, это существо проводило на двух ногах гораздо больше времени, чем шимпанзе, но и совсем прямоходящим его назвать трудно. Вообще, основание черепа – самое продвинутое место сахелянтропа.
Лицо Тумая относительно мозгового отдела заметно уменьшено. При взгляде на него в душе истинного антрополога распускаются ромашки. Ведь именно таким положено быть лицу настоящего Великого Предка: в меру крупное, не уплощенное в горизонтальной плоскости и выступающее вперед в вертикальной, но без крайностей, характеризующих горилл и шимпанзе. Анфас Тумай весьма напоминает некоторых самок бонобо: глазницы квадратные, нос широкий, альвеолярный отросток умеренно высокий. Однако по ряду признаков сахелянтроп отличается от шимпанзе: так, у него очень широкое межглазничное расстояние. В профиль же он ни на кого не похож, разве что на ардипитека: верхняя половина лица вертикальна, а челюсти равномерно выдвинуты вперед. Даже от грацильных австралопитеков отличия достаточно очевидны: у Тумая намного слабее развит альвеолярный прогнатизм (Guy et al., 2005). Правда, нижняя челюсть выглядит весьма тяжелой, даром что она невелика.
Зубы сахелянтропа довольно крупные, но их относительные пропорции замечательны: тогда как моляры больше, чем у шимпанзе, клыки – намного меньше. И это притом, что Тумай – самец! Тут может быть два объяснения: первым делом так и подмывает сказать, что клыки сахелянтропа редуцировались и это весьма прогрессивно; однако, учитывая данные по проконсулам, приходится признать, что скорее уж правильнее говорить, что это у шимпанзе клыки увеличились. Если взять по модулю уменьшение от проконсула к сахелянтропу и увеличение от него же к шимпанзе, то шимпанзе оказывается самым прогрессивным, коли уж прогрессом мы считаем отличие от исходника.
Судя по составу фауны, местность, где жил сахелянтроп, включала и водоемы, и саванны, и леса (Le Fur et al., 2014), так что у него был широкий выбор, где жить и чего есть.
Конечно, наука на то и наука, что дело не могло обойтись без скептиков, подбросивших ложку дегтя в праздничное варенье первооткрывателей Тумая (Wolpoff et al., 2006). Они указали на целый ворох примитивностей сахелянтропа, роднящих его с миоценовыми человекообразными типа кениапитека, уранопитека и даже гигантопитека. По ряду параметров основания черепа Тумай больше схож с некоторыми самками горилл, нежели австралопитеками, не говоря уж о людях. Однако чего и ждать от Великого Предка? На то он и предок, чтобы быть похожим на других родственников.
В итоге многие ученые склонны отводить сахелянтропу почетную роль “двоюродного прапрапрадедушки”, но не считают его одним из наших непосредственных предков. Для окончательного прояснения вопроса нужны новые находки. Пока их явно недостаточно, но богатство местонахождения Торос-Меналла оставляет надежду на скорые открытия.
К великому сожалению, от сахелянтропа не сохранились кости посткраниального скелета. Зато они найдены в Кении. Из ее земли на эволюционную арену на почти прямых ногах ковыляет…
Оррорин Orrorin tugenensis. Его останки фрагментарнее, но одновременно многочисленнее. Их обнаружили также французские антропологи в местности Туген-Хиллс на исходе 2000 года, отчего новооткрытый Великий Предок получил по-голливудски броское прозвище Миллениум Мен – Человек Тысячелетия. Официальное описание состоялось в 2001 году. (Senut et al., 2001). Кости были найдены в области Баринго, в местности Капчеберек, в формации Лукейно, и были датированы 5,72–5,88 млн лет назад (Deino et al., 2002).
Дюжина окаменелостей включала два куска нижней челюсти, зубы, фрагменты плечевой, фаланги кисти, правой и двух левых бедренных костей. В этом суповом наборе поместилось аж пять индивидов, как самцов, так и самок. Следы зубов кошачьего на бедренной кости повествуют печальную историю о нелегкой судьбе пионеров прямохождения…
Кстати, о дежавю и пользе статистики…
На самом деле, останки орроринов были известны и раньше, только тогда никто не знал, что это и с чем его надо есть. Еще в 1974 г. в Кении, в Лукейно, в местонахождении Чебойт, был найден фрагмент челюсти с нижним моляром KNM LU 335, но долгие годы – почти тридцать лет – он оставался “вещью в себе”, непонятным добавлением к существовавшей тогда картине эволюции австралопитеков, так как его было не с чем сравнить. Было ясно, что он очень древний (5,72–5,88 млн лет назад), похожий и на шимпанзе, и на человека, сравнительно небольшой, но лишь после описания более представительных материалов он обрел смысл.
Зубы орроринов размером примерно как у шимпанзе, более поздних ардипитеков и афарских австралопитеков, но клыки сравнительно небольшие, особенно в сравнении с шимпанзе.
Гораздо больше информации можно извлечь из длинных костей оррорина. Обломок плечевой кости очень мощный, очевидно, руки у оррорина были могучие, судя по специфике прикрепления мышц – отлично приспособленные к лазанию по деревьям. О том же свидетельствует сильный изгиб фаланги кисти.
Главный интерес вызвали бедренные кости. Одной из важнейших особенностей оррорина оказался хорошо выраженный комплекс прямохождения. Сравнение их с костями современных человекообразных обезьян и людей позволило утверждать, что уже почти 6 млн лет назад в Восточной Африке жили практически полностью прямоходящие существа. Самое поразительное – по некоторым признакам оррорины выглядят даже более прогрессивными, чем гораздо более поздние афарские австралопитеки (Pickford et al., 2002). Некоторые журналисты даже переврали это в обычно-сенсационном ключе, объявив о “перевороте в представлениях о человеческой родословной”: дескать, вовсе не известные доныне австралопитеки были нашими действительными предками, а новооткрытые оррорины. Особо обращает на себя внимание прямизна бедренной кости, передне-задняя уплощенность, длина и наклон ее шейки, а также сферичность головки и другие тонкости морфологии. Впрочем, другие исследователи более осторожно подходят к вопросу (Richmond et Jungers, 2008).
Оррорины были совсем некрупными созданиями – около 1,1–1,2 м (максимум 1,44) ростом и 35–50 кг весом (Nakatsukasa et al., 2007). Видимо, это был средний размер для всех ранних и большинства грацильных австралопитеков.
Скачут ножки по дорожке: такие ли уж прыткие были оррорины?
Многое в жизни зависит от точки зрения. В палеоантропологии дело часто обстоит так же. Потому-то одни и те же находки разные исследователи склонны расценивать совершенно по-разному. Кости не меняются, а их интерпретации разнятся до противоположностей. Не избежал этой чехарды мнений и Orrorin tugenensis – один из древнейших известных гоминид.Находка была разрекламирована как сенсация века и даже тысячелетия. Шутка ли – получалось, что именно Человек Тысячелетия был нашим прямым предком, а ставшие уже привычными афарские австралопитеки – просто тупиковая ветвь.Однако в первых описаниях бедренные кости орроринов сравнивались только с современными видами. А ведь нынешние человекообразные обезьяны – вовсе не те звери, что скакали по ветвям миоценовых лесов миллионы лет назад. И гориллы, и шимпанзе прошли собственный долгий путь эволюции, особенности их бедренных костей, хоть частенько и считаются примитивными, в действительности сформировались едва ли не позже нашего прямохождения и имеют специфику, в немалой степени обусловленную адаптацией к полуназемному образу жизни. Исправить этот пробел сравнительной палеоантропологии взялась международная испано-американо-итальянская команда антропологов (Almecija et al., 2013). Они сравнили бедренные кости орроринов не только с современными обезьянами и людьми, но и миоценовыми испанопитеком, дриопитеком, экваториусом и проконсулом. Из этой плеяды только проконсул является вероятным предком орроринов, но эволюционный уровень у них всех заведомо примитивный. Результат сопоставления оказался полностью закономерным, хотя для кого-то, может быть, и неожиданным.Во-первых, современные орангутаны, гориллы и шимпанзе отличаются от миоценовых гоминоидов, причем по некоторым чертам в ту же сторону, что и современный человек; гиббон, что характерно, сохранил древнее строение. Во-вторых, по тем же признакам оррорин оказывается примитивным, схожим с приматами, жившими 15–20 млн лет назад, но в целом промежуточным между миоценовыми обезьянами и более поздними австралопитеками, от которых те же признаки плавно видоизменяются до состояния “ранних Homo” и далее до современного человека.Самой показательной чертой оказалось расположение ягодичной бугристости: у миоценовых гоминоидов, современных гиббонов и оррорина она смещена на боковую поверхность кости, а у современных крупных человекообразных обезьян, древних гоминид и людей расположена на задней стороне бедра. А ведь на эту бугристость крепится ягодичная мышца, которая у разных существ может выполнять удивительно разнообразные функции. У человека она выпрямляет ногу, тогда как у обезьян скорее отводит ее в сторону. У орроринов она, получается, была совсем дремучей, а нога не могла полностью распрямляться в тазобедренном суставе. Столь же архаично выглядят и другие признаки Человека Тысячелетия: морфология головки бедра, угол шейки к диафизу, выступание вбок основания большого вертела и завернутость внутрь малого вертела.Таким образом, миф о необычайной продвинутости прямохождения оррорина не подтвердился. Конечно, это не отменяет наличия у оррорина действительно очень прогрессивных черт, например очень длинной шейки бедра, уплощенной спереди назад, а также сферической формы головки.Получается, что Orrorin tugenensis закономерно занимает промежуточное положение между древнейшими человекообразными и современными людьми, тогда как нынешние крупные человекообразные вовсе не копии своих ископаемых предков. Только гиббоны, чьи пращуры никогда не спускались с деревьев, сохранили примитивное состояние бедренной кости. Зато как поменялись их руки!В очередной раз можно убедиться, что все течет, все меняется, эволюция – постоянный процесс и никакие биологические объекты не замирают в морфологическом трансе (ну, разве кроме щитней Triops cancriformis, у которых День сурка начался 230 млн лет назад да так и не закончился, но это совсем-совсем другая история…).
Таким образом, оррорины, видимо, вставали на две ноги, но четвероного-древесное прошлое еще явно сквозило в их облике. Время меж тем шло, прогресс тоже не стоял на месте. Появлялись новые виды…
Ардипитек кадабба Ardipithecus kadabba – пока недостаточно изученное, но очень важное звено эволюции человека (Haile-Selassie, 2001). Его разрушенные временем останки были найдены в Эфиопии, в долине реки Средний Аваш, в местонахождениях Алайла и Аса-Кома 3; они имеют возраст 5,2–5,8 млн лет назад. Первые находки были сделаны еще в 1997 году, но продолжались и позже: в 2002 году там же (Haile-Selassie et al., 2004), а после похожие зубы были найдены в Гоне (Simpson et al., 2007).
От кадабб сохранились в основном зубы, но есть обломки нижней челюсти, ключицы, двух плечевых и локтевой, две фаланги кисти и одна – стопы. Не будучи Ж. Кювье, не так легко представить облик и образ жизни существ по столь скудным останкам. Все же ясно, что кадаббы были мелкими созданиями: нижняя челюсть меньше, чем у афарских австралопитеков, хотя и массивнее, чем у них. Клыки кадабб сопоставимы с клыками мелких самок шимпанзе: круглые в сечении, конические по форме, они превосходят клыки орроринов и более поздних гоминид. Первые нижние премоляры, найденные в Среднем Аваше, секториальные: это значит, что их коронка асимметрично скошена, чтобы большому верхнему клыку было куда поместиться. Любопытно, что в Гоне этот же зуб не секториальный – видимо, некоторые популяции были прогрессивнее.
Ключица, плечевые и локтевая имеют весьма небольшие размеры, но выраженный рельеф, свидетельствующий о неплохих способностях к лазанию по деревьям. Фаланги кисти принципиально не отличаются от таковых афарских австралопитеков, то есть сочетают обезьяньи и человеческие признаки примерно поровну.
Особенно интересна проксимальная фаланга стопы: с одной стороны, она сильно изогнута, как у человекообразных обезьян, с другой – суставная поверхность ее основания ориентирована вверх, как то характерно для афаренсисов и современных людей, что косвенно свидетельствует о прямохождении.
Кстати, опять о статистике…
Еще в 1967 г. в Лотагаме в Кении был найден фрагмент нижней челюсти с одним моляром KNM – LT 329, имеющий древность 5,0–5,5 млн лет назад. Он долго оставался “бедным родственником” – безымянным и не находящим себе места в общей родословной, а ныне отнесен к ардипитекам.
Все вышеназванные находки интересны и замечательны, но меркнут на фоне Великого Предка – ардипитека рамидуса.
Ардипитек Рамидус Ardipithecus ramidus – образцовый обезьяночеловек, то самое достающее звено, которое гипотетически реконструировали еще в XIX веке, потом так долго искали и так жаждали увидеть воочию. В нем воплотились все научные предсказания, сделанные антропологами за полторы сотни лет. Более того – он превзошел все ожидания и возложенные на него надежды!
Кстати, еще разок о статистике…
Надо сказать, что останки ардипитеков были известны задолго до описания в качестве самостоятельного вида. Еще в 1980-х годах в кенийском местонахождении Табарин, в формации Чемерон, были найдены обломок нижней челюсти с двумя молярами KNM-TH 13150 (4,43 млн лет назад) и кусок плечевой кости KNM-BC 1745 (5,1 млн лет назад). К кому их только не относили! И только с открытием многочисленных находок ардипитеков мы теперь можем уверенно говорить, что в Табарине жили представители этого вида.
В 1993 году в эфиопской местности Арамис в долине реки Средний Аваш антропологи нашли россыпь обломков. В 1994 году по ним был описан вид Australopithecus ramidus (White et al., 1994), а спустя год он был переименован в Ardipithecus ramidus (White et al., 1995). Их возраст – 4,4 млн лет. После фрагменты челюстей, зубы и фаланги этого же вида обнаружились в эфиопском местонахождении Аз-Дума в местности Гона (4,32–4,51 млн лет назад). Показательно, что там же в более древних слоях встречены останки A. kadabba – наглядная преемственность, напрашивающаяся в учебники.
История изучения скелета ардипитека – ярчайший пример научной добросовестности. Ведь между его обнаружением – 29 декабря 1993 года – и полноценной публикацией – 2 октября 2009 года – прошло почти 16 лет! Об уникальной находке было заявлено сразу же, но подробности все это время оставались неизвестны, так что у некоторых исследователей даже зародилось сомнение: “а был ли мальчик”? Все эти долгие годы международная группа исследователей, в том числе первооткрыватель – Иоганнес Хайле-Селассие, трудились над сохранением рассыпающихся костей, реконструкцией раздавленного в бесформенный комок черепа, описанием морфологических особенностей и поиском функциональной интерпретации мельчайших подробностей строения костей. Ученые не пошли по пути предъявления миру очередной скороспелой сенсации, но действительно глубоко и тщательно исследовали самые разные аспекты находки. Для этого пришлось изучить такие тонкости сравнительной анатомии современных человекообразных обезьян и человека, которые до сих пор оставались неизвестными. Естественно, к сравнению были привлечены и данные по множеству ископаемых приматов и австралопитеков. Мало того, детальнейшим образом были рассмотрены геологические условия захоронения ископаемых останков, древняя флора и фауна, что позволило реконструировать среду обитания ардипитеков достовернее, нежели для многих более поздних австралопитеков.
В итоге все части скелета ардипитека были подробнейшим образом описаны в серии статей: череп (Suwa et al., 2009a), зубы (Suwa et al., 2009b), рука и кисть (Lovejoy et al., 2009b), таз и нога (Lovejoy et al., 2009d), стопа (Lovejoy et al., 2009a), на основании чего были сделаны выводы о его способе передвижения (Lovejoy et al., 2009с) и путях возникновения бипедии в целом (Lovejoy, 2009).
Новоописанный скелет ардипитека являет собой замечательный пример подтверждения научной гипотезы. В своем облике он идеально сочетает признаки обезьяны и человека. Фактически тот образ, который полтора столетия будоражил воображение антропологов и всех, кому небезразлично наше происхождение, стал наконец реальностью. И все равно, сколь тщательно антропологи ни вымысливали и ни высчитывали стати Великого Предка, ардипитек удивил всех.
Находки в Арамисе многочисленны – останки принадлежат не менее чем 21 особи, но наиболее важен скелет взрослой самки ARA-VP-6/500, от которого осталось около 45 % костей (больше, чем от знаменитой Люси – самки афарского австралопитека из Хадара с древностью 3,2 млн лет назад!), в том числе почти весь череп, хотя и в крайне деформированном состоянии. “Арди” – так нежно прозвали Великую Прародительницу – имела рост около 1,2 м и могла весить до 50 кг. Существенно, что половой диморфизм ардипитеков был выражен гораздо слабее, чем у шимпанзе и даже более поздних австралопитеков, то есть самцы были ненамного крупнее самок; это мы знаем благодаря сравнению клыков разных особей. Объем мозга Арди достигал 300–350 см³ – столько же, как у сахелянтропа, но меньше, чем обычно у шимпанзе.
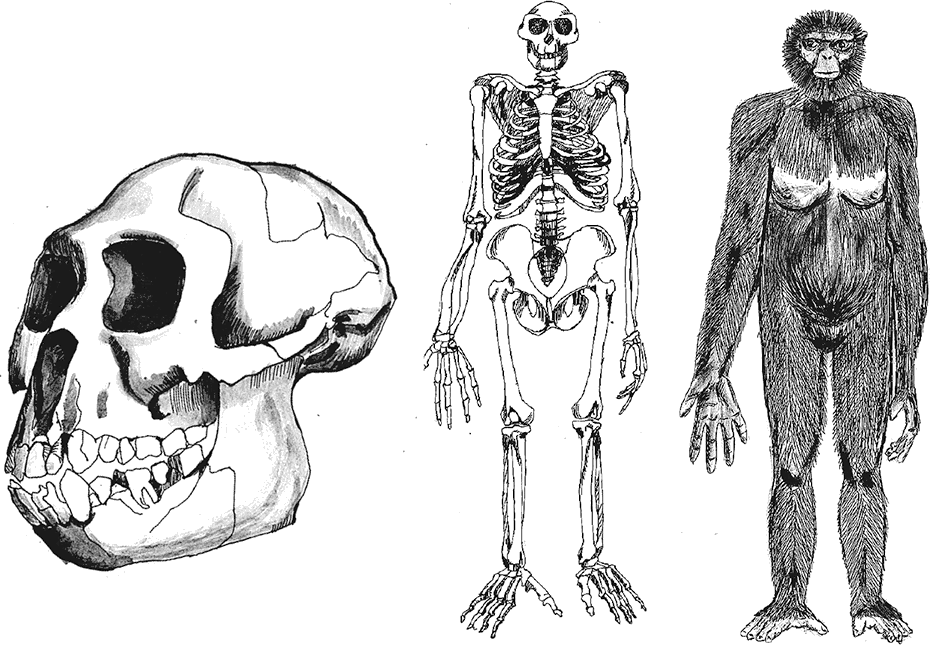
Рис. 3. Череп, скелет и реконструкция Ardipithecus ramidus.
Строение черепа ардипитека довольно примитивно, фактически обезьянье. Арди была бы мало отлична от мелкой самки шимпанзе, если бы не маленькие лицо и клыки и не сдвинутое вперед затылочное отверстие. Череп очень мал, со сравнительно слабым рельефом, без сагиттального гребня. Лоб более выпуклый, чем у сахелянтропа, маленькое слуховое отверстие и нижнечелюстная ямка височной кости – важные диагностические признаки – напрочь обезьяньи. Глазницы большие, межглазничное пространство не слишком широкое, нос можно даже назвать узким. Лицо относительно мозгового отдела довольно маленькое, но челюсти сильно выступают вперед, скуловые дуги широко расставлены, а между верхним вторым резцом и клыком имеется небольшая диастема – промежуток для нижнего клыка. Нижние челюсти ардипитеков не слишком велики, в целом как у шимпанзе и австралопитеков. Зубы имеют строение, промежуточное между шимпанзе и афарскими австралопитеками, причем по некоторым чертам ближе именно к шимпанзе. Уникальной особенностью рамидусов оказалась преимущественная редукция верхних, а не нижних клыков, так что последние оказались крупнее и обезьяноподобнее первых, больше выступающими за ряд прочих зубов.
Кстати, череп Арди весьма похож на Тумая, их различия в принципе можно бы отнести на счет полового диморфизма и индивидуальную изменчивость. Правда, разница в два с половиной миллиона лет – все же не шутка, так что антропологи не торопятся объединять эти виды.
Замечательно, что у ардипитеков лицо и зубная система не имеют специализированных черт, присущих австралопитекам и современным обезьянам; в частности, лицо Арди не было уплощено. На основе этой особенности даже выдвигалось предположение, что ардипитеки могли быть общими предками человека и шимпанзе либо даже только предками шимпанзе, но предками прямоходящими! То есть шимпанзе могли иметь двуногих прародителей. Впрочем, более тщательное исследование показало, что такая вероятность все же минимальна.
Посткраниальный скелет ардипитека – это просто праздник какой-то! В нем чудесным образом соединились черты четвероногих, древолазящих и прямоходящих существ. Обезьянья сущность Арди очевидна: мощные руки до колен, изогнутые фаланги пальцев рук, далеко отставленный в сторону и сохранивший хватательную способность большой палец стопы. Очевидно, немало времени эти существа могли проводить на деревьях и бегать по земле с опорой на ладонь, а не на фаланги согнутых пальцев. Однако большой палец руки намного длиннее, чем у человекообразных обезьян, хотя и короче, чем у человека. Таз Арди по пропорциям оказывается идеально средний между узким и высоким тазом обезьян и широким и низким – людей. У ардипитека он расширен, но и в высоту немало вытянут.
Сколь ни выражены четвероногие и древесные черты ардипитека, прямохождение его тоже достаточно очевидно – по строению таза, бедренной и берцовых костей. А больше всего удивляет стопа Арди: при оттопыренном хватательном большом пальце, шимпанзоидной таранной кости и изогнутых плюсневых имеются прилично развитые продольный и поперечный своды стопы – очевидные признаки прямохождения.
Минутка фантазии
Спуск с деревьев на землю был подготовлен целой серией преадаптаций. Увеличение размеров тела у первых человекообразных привело к уменьшению попрыгучести, утрате хвоста и четвероногому хождению по горизонтальным ветвям; дальнейший рост размеров сделал обезьян вертикально лазающими животными. Вертикальное лазание – отличная преадаптация к спуску на землю и прямохождению. Но вот уже на земле могли случиться неожиданные вещи. Скажем, при двуногости далеко не очевидно – надо ли сохранять руки? Нелетающие птицы запросто избавляются от лишних придатков. Мадагаскарские эпиорнисы и новозеландские моа утеряли крылья до полного отсутствия. Кенгуру и тушканчики находятся на пути к тому же и сохраняют лапки только потому, что иногда все же опираются на них. За несколько миллионов лет привыкания к земле ранние австралопитеки могли распрощаться с уже не столь актуальными руками; ими, конечно, можно балансировать при ходьбе, можно придерживаться за нависающие ветки, но это все не принципиально. Будь руки до спуска на землю чуть более специализированы – хотя бы как у гиббонов, не говоря уж о колобусах, – и по африканским саваннам побежали бы истинно двуногие, но безрукие обезьянки.Нас спасли три вещи. Во-первых, заточенные под ветки стопы были кривенькими шлепалками, на которых быстро не поскачешь. Необходимость обороняться в саванне возросла, ведь к привычным хищникам добавилось много новых, например гиены и шакалы. Руками же можно швырять всякие разности, отгоняя врагов. Во-вторых, перевесила жадность: очень уж хотелось что-то запихивать в рот обеими руками. Вся предыдущая история приматов сделала руки необходимыми манипуляторами, без которых обезьянам крайне трудно управиться с пищей. В-третьих, K-стратегия подразумевает, что детишки должны быть прицеплены к матери, а без ловких лапок это сделать, конечно, можно (летучие мышата, скажем, держатся молочными зубками, которые по этому поводу имеют вид крючочков; еще можно таскать деток зубами за шкирку, как кошки), но все же затруднительно.Были у ардипитеков и другие потенциальные возможности: они могли остаться четвероногими или, наскучив мимолетным увлечением наземной жизни, вернуться в родные кроны. Однако миллионы лет вертикального лазания приспособили их организм к вертикальному положению тела. Даже шимпанзе и гориллы в природе регулярно ходят на двух ногах не потому, что им это позарез нужно, а потому что просто хочется. Так что переход к двуногости был хотя и не предопределенным, но логичным следствием всей предыдущей истории. Обратный же путь на деревья мы еще успеем реализовать, когда кончатся нефть и газ.
Особенно важно, что ардипитеки жили в достаточно закрытых местообитаниях, с большим количеством деревьев и зарослей. По мнению некоторых антропологов, такие биотопы исключают классическую теорию о становлении прямохождения в условиях похолодания климата и сокращения тропических лесов. О. Лавджой на основании слабого полового диморфизма ардипитеков усовершенствовал свою старую гипотезу о развитии двуногости на основе социальных и половых взаимоотношений, вне прямой связи с климато-географическими условиями.
Однако ситуацию можно рассматривать и иначе, ведь для Арамиса первоначально было реконструировано покрытие кронами деревьев примерно в 65 % – ненамного больше половины, а после – вообще менее 60 %, скорее 20–40 %. Местообитание представляло собой подобие парка с достаточно просторно стоящими деревьями. Это значит, что с дерева на дерево далеко не всегда можно было перепрыгнуть, не спускаясь на землю. А ведь как раз примерно такие условия предполагаются сторонниками гипотезы происхождения бипедии в условиях вытеснения лесов саваннами. Ясно, что тропические леса не могли исчезнуть мгновенно, а обезьяны не могли освоить саванну в течение одного-двух поколений. Замечательно, что именно этот этап так подробно изучен теперь на примере ардипитеков из Арамиса. Эти существа прекрасно чувствовали себя на деревьях, но от одного к другому им надо было добираться по земле, поэтому они одинаково ловко лазали по ветвям и ходили на двух ногах, а иногда, видимо, и на четвереньках.
Питались ардипитеки, судя по строению зубов, самыми разными растениями, как побегами с листьями, так и плодами, избегая какой-либо специализации, что стало залогом будущей человеческой всеядности. Все же, по данным изотопного анализа зубов, они потребляли меньше растений открытых местностей, чем более поздние австралопитеки.
Понятно, что о социальной структуре ардипитеков нам ничего не известно, но малые размеры клыков и слабый половой диморфизм свидетельствуют о невысоком уровне агрессии, слабой межсамцовой конкуренции и, видимо, меньшей возбудимости, что вылилось через миллионы лет в способность современного человека сосредотачиваться, обучаться, тщательно, аккуратно и слаженно выполнять трудовую деятельность, кооперироваться, координировать и согласовывать свои поступки с прочими членами группы. Именно эти параметры отличают человека от обезьяны. Приятно, что они сформировались мало того что раньше человеческой морфологии, но и стали в немалой степени причиной этой морфологии.
Важный вывод, сделанный на основе сравнительного изучения ардипитеков, шимпанзе, горилл и современных людей, состоит в том, что многие черты человекообразных обезьян возникли независимо. Лишний раз подтвердилась давняя гипотеза параллельного появления такой специализированной особенности, как передвижение на согнутых фалангах пальцев рук у шимпанзе и горилл. Раньше считалось, что от линии гоминид сначала отделилась единая линия человекообразных обезьян, а потом она разделилась на горилл и шимпанзе; генетики посеяли основательные сомнения в таком раскладе, но палеонтологических подтверждений до открытия ардипитека не хватало. Теперь мы знаем, что шимпанзе по ряду признаков больше похожи на ардипитеков, чем на горилл, так что отделение линии горилл должно было произойти до того момента, когда появилась специализация к передвижению на фалангах пальцах, ведь у ардипитеков ее нет и, судя по всему, никогда не было (правда, эта гипотеза имеет свои слабые стороны, дело при желании можно представить и иначе, но в совокупности с генетическими данными картина вырисовывается именно такая).
Сравнение ардипитеков с сахелянтропом и более поздними австралопитеками лишний раз показало, что эволюция человеческих предков шла некими рывками. Общий уровень развития у сахелянтропа 6–7 млн лет назад и ардипитека 4,4 млн лет назад практически одинаков, тогда как спустя “всего” 200 тыс. лет (4,2 млн лет назад) у анамских австралопитеков появилось множество новых особенностей, которые, в свою очередь, мало менялись вплоть до времени появления “ранних Homo” 2,3–2,6 млн лет назад. Такие “скачки” или “повороты” эволюции были известны и до описания ардипитека, но теперь мы имеем возможность определить точное время еще одного из них; можно и попробовать объяснить их, увязав, например, с климатическими изменениями.
Одно из удивительнейших заключений, которые можно сделать из изучения ардипитеков, – это то, что человек по множеству признаков отличается от общего с шимпанзе предка меньше, чем шимпанзе или горилла. Причем это касается прежде всего размеров челюстей и строения кисти и стопы – частей тела, на особенность строения которых у человека чаще всего обращают внимание. По сути, многие детали у человека примитивнее, чем у современных обезьян, если за прогресс считать уровень отличия от общего предка. Ясно, что такой вывод может сильно не понравиться многим и многим людям (в первую очередь тем, кто склонен мыслить религиозными категориями или вообще идеализировать человечество). Но в том и состоит сила науки, что она оперирует фактами, а не эмоциями.
Рамидусы положили основание новому роду австралопитековых – собственно австралопитекам Australopithecus. Замечательно, что у нас есть достающее звено между этими звеньями: в эфиопском местонахождении Аса-Исие (расположенном все в той же долине Среднего Аваша) с датировкой 4,1–4,2 млн лет назад обнаружены челюсти, зубы и кости, промежуточные между Ardipithecus ramidus и Australopithecus anamensis (White et al., 2006). Челюсть из Аса-Исие отличается большими клыками, выступающими за ряд прочих зубов. Позвонки и фаланга кисти крупнее, чем у афарских австралопитеков. Фаланга, кроме прочего, имеет следы мощных сухожилий мышц-сгибателей – свидетельство лазания по деревьям. Бедренная кость выпрямленная, на ней черты прямохождения (в частности, след прикрепления большой ягодичной мышцы, хотя и слабо выраженный, расположенный к тому же не сзади, а несколько сбоку) сочетаются с признаками четвероногости (шероховатая линия почти отсутствует).
Австралопитек анамский Australopithecus anamensis – самый древний и примитивный в своем роде (сразу же отметим, что частенько в интернете его по неграмотности называют “аннамским”; напомним, что Аннам – старое название Вьетнама). Строго говоря, его бы можно относить уже к грацильным австралопитекам, но в морфологии анаменсисов сохранялось много архаики, а переходы между видами слишком незаметны, чтобы спорить о границах групп.
Вид был описан в 1995 году на основе многочисленных находок в кенийских местонахождениях Канапои и Аллия Бей (Leakey et al., 1995; Ward et al., 2001). Анамские австралопитеки жили 3,9–4,2 млн лет назад.
Кстати, снова о дежавю и статистике…
О существовании неких очень ранних австралопитеков, условно названных Preaustralopithecus, было известно давно, но их останки долгое время оставались слишком редкими и фрагментарными, чтобы по ним можно было сказать нечто определенное. Так, еще в 1939 г. Л. Коль-Ларсен в местонахождении Гаруси в Танзании отыскал небольшой фрагмент верхней челюсти с двумя зубами, который получил много названий (например, Praeanthropus africanus или Meganthropus africanus), в 1965 г. в Канапои в Кении был обнаружен конец плечевой кости, в 1987 г. в Белохдили в Эфиопии – кусок лобной кости австралопитека, жившего 3,8–3,9 млн лет назад, а в 1989 г. в местонахождении Фиджиж FJ4 в Южной Эфиопии – семь зубов с датировкой 4–4,18 млн лет назад. Все эти обломки не находили прочного места в схеме эволюции гоминид вплоть до описания анамского австралопитека, к коему и причисляются ныне.
От анамских австралопитеков сохранилось довольно много частей, хотя большей частью довольно фрагментарных. Целых черепов пока нет, но ясно, что височная кость примитивна: слуховое отверстие маленькое, а вот суставная ямка немножко продвинутее, чем у рамидуса, – у анаменсисов появился суставной бугорок, хотя бы и маленький и уплощенный. Челюстей найдено достаточно, чтобы сказать, что у анамских австралопитеков морда и зубы в среднем были чуть больше, чем у афаренсисов. Форма альвеолярной дуги, как и у предшествующих гоминид, типично обезьянья – вытянутая, с параллельно расположенными рядами заклыковых зубов. Мало отличается от понгидного варианта и нижняя челюсть, имеющая к тому же некоторые намеки на специализацию. Примитивность выражалась в асимметрии первого нижнего премоляра – для удобства расположения верхнего клыка, который, кстати, имел очень мощные корни. Моляры, хотя абсолютно и велики, нельзя назвать мегадонтными. А ведь мегадонтия – когда зубы относительно размеров челюсти сильно увеличены – была типична для грацильных австралопитеков, не говоря уж о парантропах. Очевидно, во времена анаменсисов мегадонтия еще не была в тренде.
Плечевая и кости кисти сочетают черты человекообразных обезьян и людей примерно в равных пропорциях. Судя по большой берцовой кости, анамские австралопитеки были вполне прямоходящими и по способу передвижения уже не отличались от афаренсисов, да и от нас тоже. А вот лучевая кость свидетельствует о другом: изгиб ее нижней части и ориентация лучезапястной суставной поверхности (умно это звучит так: “сильный медиальный изгиб дистального конца и дистальное выступание дорзального края дистального конца” – да, именно на таком странном языке общаются антропологи!) говорят о блокировании разгибания запястья. А ведь аналогичное строение лучевой кости характерно для крупных человекообразных, ходящих с опорой на фаланги согнутых пальцев. Напомним, что у ардипитека такой специализации не было. Выходит, по крайней мере некоторые популяции анаменсисов могли возвращаться на четвереньки? Впрочем, индивидуальную изменчивость никуда не денешь, и пока у нас не будет больше материалов, утверждать что-то уверенно ни один антрополог не станет.

Рис. 4. Череп Australopithecus anamensis.
Кстати, лучевая кость очень длинная, соответствующая росту современного человека в 1,75 м, притом что по другим костям рост получается около полутора метров. Очевидно, пропорции рук анаменсисов были еще не совсем человеческими.
Некоторые специализации анаменсисов выглядят своеобразно, так что не все ученые видят этот вид в числе наших предков. Однако в целом они представляют собой хорошее звено между рамидусами и афаренсисами.
Достающее звено между самими анаменсисами и афаренсисами найдено не так давно в эфиопском местонахождении Ворансо-Милле, в слоях с датировками 3,57–3,8 млн лет назад (Haile-Selassie et al., 2010b). Челюсти и зубы, обнаруженные тут, настолько промежуточны, что их нельзя однозначно отнести ни к Australopithecus anamensis, ни к A. afarensis. Несколько зубов и бедренная кость из Галили также хронологически и морфологически зависают между двумя этими видами.
Пока мы крайне мало знаем про образ жизни ранних австралопитеков, однако с каждым годом количество находок растет, а знания об окружающей среде того далекого времени ширятся. Обитали ранние австралопитеки в лесистых, иногда даже болотистых местах, а также в лесостепях. Именно в это время – у ранних австралопитеков – появилась главная отличительная черта гоминид – прямохождение. Оно сочеталось у них с древолазанием и – по крайней мере, у самых ранних – четвероногостью. Не исключено, например, что они проводили на деревьях полные опасности ночи. Признаки таких древних способов передвижения сохранялись и много позже – вплоть до “ранних Homo”, у коих они больше являются генетическим воспоминанием о древесных пращурах. Вообще, в сумме обезьяньих и человеческих признаков в скелете ранних австралопитеков оказывается примерно поровну, что дает возможность говорить о них как о том самом пресловутом “недостающем звене”, отсутствие которого столь часто ставят в упрек ученым креационисты и разнообразные пара– и псевдоученые. Учитывая, что ранние австралопитеки уже больше двадцати лет как найдены и продолжают изучаться – это самое что ни на есть “достающее” звено.
Обобщенно ранних австралопитеков можно описать как существ ростом чуть выше метра, с головой, почти неотличимой от обезьяньей, имеющих к тому же размеры мозга, как у шимпанзе, однако с довольно маленькими клыками, с руками до колен, приспособленными для хватания предметов, но с несколько изогнутыми пальцами, с почти прямыми ногами, широким, почти человеческим тазом, с примитивной, но имеющей своды стопой.
Некоторые детали строения ранних австралопитеков позволяют предположить, что не все они выстраиваются в одну прямую линию. В разное время разные исследователи предполагали, что, возможно, сахелянтропы были родственниками горилл, ардипитеки – непосредственными предками современных шимпанзе, а анамские австралопитеки вымерли, не оставив потомков. С другой стороны, не так уж запредельно они различаются, а основные черты свидетельствуют о более-менее прямой преемственности. Но как сравнивать двух австралопитеков, если от одного сохранился череп, а от другого – лучевая кость? Кроме того, играет роль и “человеческий фактор”: каждый исследователь любит свою находку, и зачастую в появлении нового названия немалую роль играет желание прославить если не себя, то новонайденную окаменелость. С этим же связано появление в печати сенсационных заголовков, как правило провозглашающих с открытием очередного черепа “полный переворот в воззрениях на происхождение человека”. Профессиональные антропологи относятся к таким “революциям в науке” тоже с большим энтузиазмом, но спокойно, поскольку они не столько “переворачивают” наши представления, сколько дополняют все еще далеко не полную картину.
Пока ученые не пришли к общепризнанному взгляду на родство разных видов этих существ, что вполне понятно, учитывая фрагментарность большинства находок. Кто из них ближе к нам, а кого эволюционные вихри стерли с лика Земли без следа? Пока этот вопрос остается открытым. Из описанных видов вроде бы оррорины и ардипитеки наиболее подходят на роль наших предков, но споры среди антропологов не утихают. Может, некоторые известные виды ранних австралопитеков не имеют к нам прямого отношения, а в африканской земле еще нетронутыми покоятся останки наших пращуров? Вполне возможно, учитывая, что все виды ранних австралопитеков описаны в последние пятнадцать – двадцать лет и знаем мы о них крайне недостаточно.
Древний гоминоид Ливии – ископаемый шимпанзе или ранний австралопитек?
(поучительная история)
В 1979 г. в журнале Nature вышла статья, посвященная исследованию миоценовой фауны местонахождения Сахаби в Ливии (Boaz et al., 1979). В ней упоминалась ключица гоминоида, найденная в слоях с датировкой 4–7 млн лет назад. В следующем, 1980 г. в American Journal of Physical Anthropology было опубликовано уже подробное описание этой ключицы, с уточнением датировки до 5–7 млн лет назад (Boaz, 1980). Автор – антрополог Н. Боаз – подробнейшим образом описал места прикрепления мышц, форму, степень сагиттального и коронального изгиба, размеры и пропорции кости. Интерпретации мешала лишь острая нехватка сравнительного материала по ископаемым гоминидам: на тот момент была известна всего одна древняя ключица – OH 48 из Олдувая, одна, на тот момент не описанная, – из Лаэтоли, да еще три гораздо более древних, принадлежавших плиопитекам.Большой размер ключицы, превосходящий самые крупные величины мандрилов, свидетельствовал о том, что кость, скорее всего, не принадлежит мартышковому примату. Сильный грудной изгиб – гоминидный признак – говорил о том, что кость, скорее всего, принадлежит не шимпанзе (у детенышей, самок и мелких самцов изгиб может быть большим, но он расположен в другой плоскости); с другой стороны, сильный изгиб в вертикальной плоскости как раз типичен для шимпанзе. Смещение на середину кости места прикрепления дельтовидной мышцы и мощная выраженность этого прикрепления оказались нетипичны ни для шимпанзе, ни для человека. Тип локомоции должен был отличаться и от обезьян, и от человека, и, скорее всего, от дриопитеков, хотя по ключице об этом судить трудно. Н. Боаз сделал вывод, что кость принадлежит гоминоиду, но уточнить его принадлежность помогут лишь новые находки.Напомним, это был 1980 год. В свете находок 1990-х и 2000-х годов кость из Сахаби могла бы оказаться ключицей некоего самого северного раннего австралопитека… Могла бы оказаться останками самого древнего и северного шимпанзе… Могла бы… Но! Она оказалась ребром дельфина (White et al., 1983). В Сахаби, наряду с останками наземной фауны (включая, кстати, и древних мартышкообразных приматов), были найдены зубы акул и фрагменты костей китообразных. Судя по всему, именно в этот водоплавающий ряд попадает и “ключица гоминида”. Такой вот неожиданный финал несостоявшейся сенсации.Занятно, что Н. Боаз в статье упомянул несколько случаев неверной идентификации “гоминидных ключиц”: MLD 20 и MLD 36 из Макапансгата в Южной Африке оказались при перепроверке боковыми метакарпалиями древней лошади-гиппариона, одна с острова Русинга и одна из Мабоко в Кении – бедренными костями рептилий. Однако ни знание этих прецедентов, ни знание анатомии самых разнообразных животных не спасло Н. Боаза от такой же ошибки. Попробуйте пересмотреть больше трех тысяч фрагментов костей десятков видов животных (а именно столько было найдено в Сахаби) и ни разу не ошибиться! Слишком уж много костей, и удивительно сильно иногда похожи даже неодинаковые части неодинаковых животных. Некоторые несознательные личности на этом месте воскликнут: “Ну вот! Бестолковые ученые! Опять все перепутали! То зуб свиньи посчитают предком человека, то ногу лошади, то ребро дельфина! Как можно им верить!” Однако ж стоит напомнить, что именно ученые разобрались, чем зуб свиньи отличается от человеческого, а метакарпалия гиппариона и ребро дельфина – от ключицы гоминида. Ошибки были найдены и распознаны. Без грандиозного багажа знаний это было бы невозможно, и именно такой багаж, несмотря на случающиеся недоразумения, служит надежным фундаментом для целостной и непротиворечивой картины мира. А ошибки… Что ж, на то и ошибки, чтобы на них учиться.
Назад: Пролог
Дальше: Глава 2 Человекообезьяны: грацильные австралопитеки

