23. Концентрация внимания
Знания о пространстве играют ключевую роль в поведении всех живых существ, от моллюсков до людей. Как отмечал Джон О’Киф, “пространство играет заметную роль во всем нашем поведении. Мы в нем живем и передвигаемся, исследуем его и защищаем”. Ощущение пространства – не только важное, но и интересное явление, потому что, в отличие от других ощущений, у него нет собственного органа чувств. Как же наш мозг получает внутреннее представление о пространстве?
Кант, один из тех мыслителей, чьи идеи лежат у истоков когнитивной психологии, доказывал, что способность представлять пространство встроена в наш мозг. Он писал, что в людях от рождения заложены основы восприятия пространства и времени, поэтому, когда у нас возникают какие‑либо ощущения (будь то зрительные образы, звуки или что‑то осязаемое), они автоматически определенным образом соотносятся с нашими представлениями о пространстве и времени. О’Киф применил логику кантовских рассуждений о пространстве к эксплицитной памяти. Он доказывал, что во многих формах эксплицитной памяти (например, в памяти на людей и предметы) используются пространственные координаты, то есть людей и события мы обычно запоминаем в пространственном контексте. Эта идея была не нова. Еще в 55 году до н. э. Цицерон, великий древнеримский поэт и оратор, описывал греческий метод запоминать слова (по сей день используемый некоторыми актерами), представляя себе ряд комнат в большом доме, ассоциируя слова с каждой из них, а затем мысленно проходя по этим комнатам в правильном порядке.
Поскольку у нас нет специального органа чувств для восприятия пространства, представление о пространстве является в полной мере когнитивной способностью, имеющей самое непосредственное отношение к проблеме связывания. Для получения этого представления мозг должен объединять входящие сигналы, получаемые от нескольких разных сенсорных модальностей (ощущений), а затем создавать единое внутреннее представление, которое основано на сигналах, поступающих от всех модальностей вместе, а не от какой‑то одной из них. Информация о пространстве обычно представлена у нас в мозге во многих участках и многими разными способами, причем свойства каждого из этих представлений могут меняться в зависимости от того, какой цели они служат. Для некоторых представлений пространства мозг обычно использует эгоцентрические координаты (сосредоточенные на получателе сигналов), в которых кодируется, например, положение пятна света на сетчатке, или источника запаха, или осязательных ощущений относительно тела. Эгоцентрическое представление служит людям и обезьянам для ориентации на источник внезапного звука, в сторону которого направляется взгляд, дрозофилам – для избегания источника запаха, с которым связаны неприятные ассоциации, и аплизиям – для работы рефлекса втягивания жабр. Для других форм поведения, например связанных с пространственной памятью у мышей или людей, мозг должен кодировать положение организма по отношению к окружающему миру и положение объектов по отношению друг к другу. Для этих целей мозг пользуется аллоцентрическими (сосредоточенными на окружающем) координатами.
Исследования более простых осязательных и зрительных сенсорных карт головного мозга, основанных на эгоцентрических координатах, стали плацдармом для исследований более сложных аллоцентрических представлений пространства. Но пространственная карта, которую открыл О’Киф в 1971 году, принципиально отличается от эгоцентрических осязательных и зрительных сенсорных карт (которые открыли Уэйд Маршалл, Вернон Маунткасл, Дэвид Хьюбел и Торстен Визел), потому что она основана на разных ощущениях, а не на каком‑то одном из них. В самом деле, когда мы с Олденом Спенсером в 1959 году пытались разобраться в том, как сенсорная информация поступает в гиппокамп, мы регистрировали сигналы отдельных нейронов, действуя раздражителями на разные органы чувств, но не смогли получить ярко выраженной реакции. Тогда мы еще не понимали, что гиппокамп задействован в восприятии окружающего, поэтому в нем представлены сложные мультисенсорные (связанные с несколькими органами чувств) ощущения.
Джон О’Киф первым понял, что в гиппокампе крыс содержится мультисенсорное представление окружающего пространства. Он выяснил, что, когда крыса ходит по клетке, в некоторых нейронах места потенциалы действия запускаются лишь тогда, когда она занимает определенное положение, а если она переходит в другое место, потенциалы действия запускаются в других нейронах. Мозг разбивает окружающее на множество небольших перекрывающихся областей, похожих на элементы мозаики, и каждая из них представлена активностью определенных клеток гиппокампа. После того как крыса попадает в новую среду, у нее за минуты вырабатывается эта внутренняя пространственная карта.
Я начал размышлять о пространственных картах в 1992 году и задумался, как они формируются, как поддерживаются и как внимание управляет их формированием и поддержанием. Меня поразил тот установленный О’Кифом и его коллегами факт, что даже пространственная карта очень просто устроенного места формируется не мгновенно, а в течение десяти – пятнадцати минут после того, как крыса попадает в новую среду. Этот факт заставлял предположить, что формирование пространственной карты есть процесс обучения и что повторение – мать учения и в случае с пространством. При оптимальных условиях эта карта не меняется неделями или даже месяцами, совсем как некоторые формы памяти.
В отличие от зрения, осязания и обоняния, обеспечиваемых врожденными системами и основанных на кантовских априорных знаниях, пространственные карты демонстрируют новый тип сенсорных представлений, основанный на сочетании априорных знаний и обучения. Общая способность формировать сенсорные карты встроена в мозг, но частные сенсорные карты в него не встроены. Клетки места, в отличие от нейронов сенсорных систем, включаются не внешними раздражителями. Совместная деятельность этих клеток соответствует месту, в котором животное думает, что находится.
Мне же теперь хотелось узнать, не служат ли для формирования и поддержания пространственной карты те же молекулярные механизмы, которые обеспечивали долговременную потенциацию и работу пространственной памяти в наших экспериментах с гиппокампом. Хотя О’Киф открыл клетки места еще в 1971 году, а Блисс и Лемо обнаружили долговременную потенциацию в гиппокампе в 1973 году, никто еще не пытался связать эти два открытия друг с другом. В 1992 году, когда мы начали изучать пространственные карты, о молекулярных процессах, обеспечивающих их формирование, еще не было известно. Эта ситуация лишний раз доказывает, почему работа на стыке двух дисциплин (в данном случае – клеточной биологии нейронов места и молекулярной биологии межклеточной передачи сигналов) нередко оказывается очень плодотворной. Задачи и результаты научного эксперимента во многом определяются интеллектуальным контекстом, в рамках которого работает экспериментатор. Мало что в науке может сравниться по увлекательности с внедрением нового образа мышления, связанного с одной дисциплиной, в другую. Именно такое перекрестное оплодотворение научных дисциплин и имели в виду мы с Джимми Шварцем и Олденом Спенсером, когда в 1965 году назвали наше новое подразделение Нью-Йоркского университета отделением нейробиологии и поведения.
Наша совместная работа с Робертом Маллером, одним из первых исследователей клеток места, показала, что некоторые молекулярные процессы, обеспечивающие долговременную потенциацию, действительно необходимы и для долговременного сохранения пространственной карты. Мы знали, что протеинкиназа A включает структурные гены, тем самым запуская синтез белков, необходимых для осуществления поздней фазы долговременной потенциации. Аналогичным образом, хотя ни протеинкиназа A, ни синтез белков не требуется для запуска формирования пространственной карты, и то и другое необходимо для того, чтобы надолго закрепить эту карту, давая мыши возможность вспоминать ее каждый раз, когда она попадает в ту же среду.
Открытие того, что протеинкиназа A и синтез белков требуются для закрепления пространственной карты, поднимало еще один вопрос. Дает ли животным эксплицитную пространственную память именно та карта, которую мы наблюдаем в гиппокампе, то есть она ли позволяет им демонстрировать свое знакомство с соответствующей средой? Составляют ли такие карты то самое внутреннее представление пространства – нейронную основу эксплицитной пространственной памяти? О’Киф с самого начала рассматривал когнитивные карты как внутренние представления пространства, которые служат животным для ориентирования. Поэтому он видел в таких картах орудие ориентирования вроде компаса, а не собственно память. Мы исследовали этот вопрос и выяснили, что, если блокировать работу протеинкиназы A или подавлять синтез белков, это приводит к нарушениям не только долговременной устойчивости пространственной карты, но и способности к долговременному сохранению пространственной памяти. Таким образом, мы получили прямые генетические свидетельства того, что данные карты связаны с пространственной памятью. Кроме того, мы установили, что пространственная память, как и простая эксплицитная память, связанная с рефлексом втягивания жабр у аплизии, тоже делится на краткосрочную и долгосрочную составляющие, то есть процессы, задействованные в выработке карты (и сохранении ее в течение нескольких часов) и долговременном поддержании ее в устойчивой форме.
Несмотря на некоторые черты сходства, между нашей эксплицитной пространственной памятью и имплицитной памятью есть принципиальные отличия. В частности, для записи и считывания эксплицитной памяти требуется избирательное внимание. Поэтому теперь, чтобы исследовать связь эксплицитной памяти с нервной деятельностью, нам нужно было обратиться к проблеме внимания.
Считается, что избирательное внимание играет немалую роль в восприятии, действии и памяти, обеспечивая единство сознательного опыта. На каждое животное в любой момент времени действует море разнообразных сенсорных раздражителей, но животное обращает внимание лишь на один или очень немногие из них, игнорируя или подавляя действие остальных. Способность мозга обрабатывать сенсорную информацию намного скромнее, чем способность связанных с ним рецепторов собирать эту информацию из окружающей среды. Внимание служит фильтром, отбирающим некоторую часть поступающих сведений для дальнейшей обработки. Во многом именно благодаря избирательному вниманию внутренние представления не воспроизводят окружающий мир во всех его подробностях, а по одним лишь сенсорным раздражителям нельзя предсказать любую моторную реакцию. В своих сиюминутных ощущениях мы настроены на какие‑то определенные компоненты сенсорной информации, а всеми остальными пренебрегаем (в той или иной степени). Когда вы отрываете взгляд от этой книги, чтобы посмотреть на человека, входящего в комнату, вы уже не обращаете внимания на слова, напечатанные на странице. При этом вы не обращаете внимания и на то, как выглядит комната, и на других людей, которые могут в ней находиться. Если впоследствии вас попросят описать свои ощущения, вы, скорее всего, вспомните, что в комнату вошел человек, а не то, что на стене была небольшая царапина. Эта способность настраивать сенсорный аппарат есть неотъемлемое свойство всего восприятия, как еще в 1890 году отмечал Уильям Джемс в своем эпохальном труде “Принципы психологии”: “Миллионы объектов <…> действуют на мои органы чувств, но никогда не станут частью моего опыта. Почему? Потому что они не представляют для меня никакого интереса. Мой опыт – это то, на что я готов обратить внимание. <…> Все знают, что такое внимание. Это сосредоточенность сознания в ясной и отчетливой форме на каком‑то одном из нескольких кажущихся доступными объектов или направлений мысли. По сути, это настройка, концентрация сознания. Внимание предполагает отказ от одних вещей ради того, чтобы успешнее заниматься другими”.
Внимание также позволяет нам связывать разные компоненты пространственного образа в единое целое. Постдок Клифф Кентрос и я решили исследовать связь внимания с пространственной памятью, задавшись вопросом о том, необходимо ли внимание для создания пространственной карты, а если необходимо, то как именно оно влияет на ее формирование и закрепление. Чтобы это проверить, мы испытывали мышей в четырех опытах, требующих разных степеней концентрации внимания. Первая степень – это базовое или поверхностное внимание, работающее даже в отсутствие дополнительных раздражителей. В соответствующем опыте мыши бегали по клетке, где их не отвлекали никакие раздражители. Во втором опыте они должны были находить еду (для чего требуется немного больше внимания), в третьем – отличать одну среду от другой, а в четвертом – выполнять задание на запоминание пространства. Мы сделали экспериментальную установку, где мыши, которые бегали по клетке, периодически подвергались воздействию неприятных звуков и вспышек света. Единственный способ их выключить состоял в том, чтобы найти небольшой, никак не обозначенный безопасный участок и сидеть там какое‑то время. Мыши умеют находить такой участок без особого труда.
Мы установили, что даже поверхностного внимания достаточно, чтобы у мыши сформировалась и закрепилась на несколько часов пространственная карта, но через три – шесть часов такая карта становится неустойчивой. Долговременное закрепление карты сильно и последовательно зависит от степени, с которой мыши приходится уделять внимание определенным особенностям окружающей среды. Так, если мыши приходится концентрировать внимание на новой среде, выполняя одновременно с исследованием этой среды задание на запоминание пространства, то пространственная карта сохраняет устойчивость на много дней, и мышь без труда вспоминает, как выполнять это задание, пользуясь полученными сведениями об окружающей среде.
Каков механизм концентрации внимания в мозге? Какую роль он играет в ускоренном кодировании информации об окружающем пространстве и облегченном считывании этой информации по прошествии долгого времени? Мне уже было известно, что внимание – это не просто загадочная сила, действующая в мозге, а модуляторный процесс. Майкл Гольдберг и Роберт Вурц из Институтов здоровья установили, что внимание усиливает реакцию нейронов зрительной системы на раздражители. Судя по всему, в явлениях, связанных с вниманием, был задействован модуляторный проводящий путь, медиатором в котором служит дофамин. Клетки, вырабатывающие дофамин, сосредоточены в среднем мозге, а их аксоны доходят до гиппокампа. Более того, мы выяснили, что подавление действия дофамина в гиппокампе отключает закрепление пространственной карты у животных, внимание которых было сконцентрировано на окружающем пространстве, а активация дофаминовых рецепторов гиппокампа, напротив, приводит к закреплению пространственной карты у животных, внимание которых не было сконцентрировано. Аксоны выделяющих дофамин нейронов среднего мозга посылают сигналы в несколько разных областей мозга, в том числе в гиппокамп и префронтальную кору. При этом префронтальная кора, которая участвует в преднамеренных действиях, передает сигналы обратно в средний мозг, корректируя активность этих нейронов. Наше открытие того, что некоторые из областей мозга, участвующих в произвольных действиях, участвуют также в работе внимания, подкрепляло представление о том, что избирательное внимание играет принципиальную роль в единстве сознательного опыта.
Уильям Джемс в книге “Принципы психологии” отмечал, что есть разные формы внимания. Внимание бывает по крайней мере двух типов: непроизвольное и произвольное. Непроизвольное внимание обеспечивается нейронными механизмами, действующими автоматически, и особенно ярко проявляется в формировании имплицитной памяти. Например, в опытах с выработкой классических условных рефлексов животные будут обучаться ассоциировать один раздражитель с другим в том и только в том случае, если условный раздражитель будет заметным или неожиданным. Непроизвольное внимание активируется определенными свойствами окружающего мира (или раздражителя): его привлекает, по Джемсу, “все большое, яркое, движущееся или кровь”. Произвольное внимание, такое как внимание на дороге и на движение при вождении машины, напротив, свойственно именно эксплицитной памяти и возникает из внутренней потребности обрабатывать информацию о раздражителях, которые автоматически не привлекают внимания.
Джемс утверждал, что наше произвольное внимание – это явно сознательный процесс. Поэтому у нас есть все основания полагать, что он запускается в коре головного мозга. Исходя из редукционистского подхода, в обеих формах внимания должны быть задействованы биологические сигналы о заметности раздражителя, например передаваемые модуляторными нейромедиаторами, которые регулируют деятельность или конфигурацию нейронных сетей.
Наши молекулярные работы с аплизией и мышами подтверждали представления Джемса о существовании двух форм внимания, произвольной и непроизвольной. Принципиальное различие между ними состоит не в отсутствии или присутствии заметных раздражителей, а в том, сознательно или бессознательно воспринимается сигнал о заметности. Так, когда мне нужно было научиться находить дорогу от моего дома в Ривердейле до дома моего сына в Вестчестере, я концентрировал свое внимание сознательно. Но когда мою машину, едущую по дороге, внезапно подрезала другая, я нажимал на тормоз машинально. Результаты ряда исследований также заставляют предположить, что, как и утверждал Джемс, фактором, определяющим, какая формируется память, имплицитная или эксплицитная, служит способ получения сигнала о заметности, вызывающего концентрацию внимания.
Как мы убедились, в обоих типах памяти преобразование кратковременной памяти в долговременную требует активации генов и, судя по всему, в обоих случаях модуляторные нейромедиаторы служат для передачи сигнала, обеспечивающего концентрацию внимания, который говорит о значимости раздражителя. В ответ на этот сигнал включаются гены и синтезируются белки, которые поступают во все синапсы. Например, у аплизии протеинкиназу A активирует серотонин, а у мыши – дофамин. Но между механизмами возникновения сигналов о заметности при работе имплицитной памяти, лежащей в основе сенсибилизации у аплизии, и при работе эксплицитной памяти, необходимой для формирования пространственной карты у мыши, есть принципиальная разница.
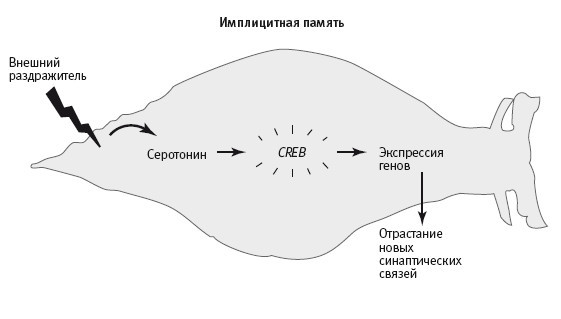
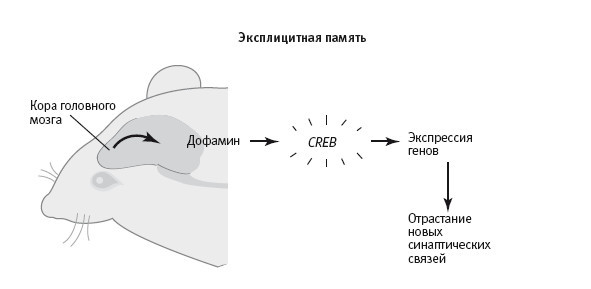
23–1. Сигнал о значимости в долговременной имплицитной и эксплицитной памяти. В случае имплицитной (бессознательной) памяти действие внешнего раздражителя автоматически вызывает сигнал о значимости (выделение серотонина), который вызывает активацию генов и обеспечивает формирование долговременной памяти. В случае эксплицитной (сознательной) памяти кора головного мозга произвольно посылает сигнал о значимости (выделение дофамина), вызывающий концентрацию внимания, которая регулирует активность нейронов гиппокампа и обеспечивает формирование долговременной памяти.
При формировании имплицитной памяти у аплизии сигнал, концентрирующий внимание, возникает рефлекторно и идет снизу вверх: удар током вызывает активацию сенсорных нейронов задней части тела, которые воздействуют непосредственно на клетки, выделяющие серотонин. А при формировании пространственной памяти у мышей выделение дофамина, судя по всему, вызывается произвольно, сверху вниз: кора головного мозга активирует клетки, выделяющие дофамин, а дофамин регулирует активность нейронов гиппокампа (рис. 23–1).
Эта идея о том, что в обеспечивающих концентрацию внимания процессах, запускаемых снизу вверх и сверху вниз, используются сходные молекулярные механизмы, подтверждалась еще одним открытым нами механизмом, который мог быть задействован в закреплении памяти в обоих рассматриваемых случаях. В гиппокампе мыши содержится по крайней мере один прионоподобный белок, похожий на те, что Каусик Си открыл у аплизии. Мартин Тайс, постдок из Германии, и я установили, что дофамин регулирует количество прионоподобного CPEB-белка (CPEB‑3) в гиппокампе мыши примерно так же, как серотонин – количество и состояние CPEB-белка у аплизии. Этот результат открывал интересную возможность (пока еще только возможность), что пространственные карты могут закрепляться за счет того, что концентрация внимания приводит к выделению дофамина в гиппокампе, которое вызывает переход в самоподдерживающееся состояние, тоже обеспечиваемое CPEB-белком.
Открытие важной роли внимания в закреплении пространственной карты поднимает еще один вопрос: похожи ли у всех нас пространственные карты, формируемые в ходе обучения? А именно – одинаковы ли стратегии, которыми пользуются мужчины и женщины для ориентации в окружающем мире? Это интереснейший вопрос, который биологи еще только начинают исследовать.
О’Киф, открывший клетки места в гиппокампе, продолжил исследования пространственной ориентации, обратившись к вопросу о различиях между полами. Он обнаружил отчетливую разницу между способами концентрации внимания при ориентации в окружающем пространстве у мужчин и женщин. Женщины пользуются при этом ближайшими ориентирами. Поэтому, если спросить у женщины дорогу, она, скорее всего, скажет: “Возле аптеки поверните направо, а потом поезжайте прямо, пока не увидите по левую руку белый дом в колониальном стиле с зелеными ставнями”. Мужчины же больше полагаются на внутренние геометрические карты. Мужчина, скорее всего, скажет: “Когда проедете пять миль на север, поверните направо и оттуда еще полмили на восток”. Томографические исследования мозга показывают, что у мужчин и женщин, когда они думают о пространстве, активируются разные участки мозга: гиппокамп левого полушария у мужчин и теменная и префронтальная кора правого полушария у женщин. Результаты этих исследований свидетельствуют о возможных выгодах групповой работы, позволяющей оптимальным образом совмещать обе стратегии.
Различия между полами в способе формирования пространственных карт приобретают дополнительное значение, если рассмотреть их в более широком контексте. В какой степени у мужчин и женщин различаются строение мозга и стиль работы памяти? Врожденные ли это отличия или они развиваются под влиянием обучения и социальных стереотипов? Ответив на эти вопросы, нейробиология поможет нам в принятии многих важных решений, регулирующих жизнь общества.

