Книга: Физиология человека. Общая. Спортивная. Возрастная
Назад: 4. Высшая нервная деятельность
Дальше: 6. Произвольные движения
5. Нервно-мышечный аппарат
У человека существует три вида мышц: поперечно-полосатые скелетные мышцы, особая поперечно-полосатая сердечная мышца и гладкие мышцы внутренних органов.
5.1. Функциональная организация скелетных мышц
Скелетные мышцы человека содержат около 300 млн мышечных волокон и имеют площадь порядка 3 м2. Целая мышца представляет собой отдельный орган, а мышечное волокно – клетку. Мышцы иннервируются двигательными нервами, передающими из центров моторные команды, чувствительными нервами, несущими в центры информацию о напряжении и движении мышц, и симпатическими нервными волокнами, влияющими на обменные процессы в мышце. Функции скелетных мышц заключаются в перемещении частей тела друг относительно друга, перемещении тела в пространстве (локомоция) и поддержании позы тела.
Функциональной единицей мышцы является двигательная единица, состоящая из мотонейрона спинного мозга, его аксона (двигательного нерва) с многочисленными окончаниями и иннервируемых им мышечных волокон. Возбуждение мотонейрона вызывает одновременное сокращение всех входящих в эту единицу мышечных волокон. Двигательные единицы (ДЕ) небольших мышц содержат малое число мышечных волокон (ДЕ мышц глазного яблока 3–6 волокон, мышц пальцев руки 10–25 волокон), а ДЕ крупных мышц туловища и конечностей – до нескольких тысяч (например, ДЕ икроножной мышцы человека – около 2000 мышечных волокон).
Мелкие мышцы иннервируются из одного сегмента спинного мозга, а крупные мышцы – мотонейронами 2–3 спинальных сегментов. Мотонейроны, иннервирующие одну мышцу, составляют общий мотонейронный пул, в котором могут находиться мотонейроны различных размеров. Большие ДЕ образованы крупными мотонейронами, которые имеют толстые аксоны, множество концевых разветвлений и большое число связанных с ними мышечных волокон. Такие ДЕ имеют низкую возбудимость, генерируют высокую частоту нервных импульсов (порядка 20–50 импульсов в 1 с) и характеризуются высокой скоростью проведения возбуждения. Они включаются в работу лишь при высоких нагрузках на мышцу. Мелкие ДЕ имеют мотонейроны небольших размеров, тонкие и медленно проводящие аксоны, малое число мышечных волокон. Они легко возбудимы и включаются в работу при незначительных мышечных усилиях. Нарастание нагрузки вызывает активацию различных ДЕ скелетной мышцы в соответствии с их размерами – от меньших к большим (правило Хеннемана).
Мышечное волокно представляет собой вытянутую клетку (ее диаметр около 10-100 мкм, а длина 10–12 см). В состав волокна входят его оболочка – сарколемма, жидкое содержимое – саркоплазма, ядро, энергетические центры – митохондрии, белковые депо – рибосомы, сократительные элементы – миофибриллы, а также замкнутая система продольных трубочек и цистерн, расположенных вдоль миофибрилл и содержащих ионы Са2+,– саркоплазматический ретикулум. Поверхностная мембрана клетки через равные промежутки образует поперечные трубочки, входящие внутрь мышечного волокна, по которым внутрь клетки проникает потенциал действия при ее возбуждении.
Миофибриллы – это тонкие волокна (диаметр 1–2 мкм, длина 2–2,5 мкм), содержащие два вида сократительных белков (протофибрилл): тонкие нити актина и вдвое более толстые нити миозина. Они расположены таким образом, что вокруг миозиновых нитей находится 6 актиновых нитей, а вокруг каждой актиновой – 3 миозиновых. Миофибриллы разделены Ζ-мембранами на отдельные участки – capкомеры, в средней части которых расположены преимущественно миозиновые нити, а актиновые нити прикреплены к Ζ-мембранам по бокам саркомера. (Разная способность актина и миозина преломлять свет создает в состоянии покоя мышцы ее поперечно-полосатый вид в световом микроскопе.)
Нити актина составляют около 20 % сухого веса миофибрилл. Актин состоит из двух форм белка: 1) глобулярной формы – в виде сферических молекул и 2) палочковидных молекул тропомиозина, скрученных в виде двунитчатых спиралей в длинную цепь. На протяжении этой двойной актиновой нити каждый виток содержит по 14 молекул глобулярного актина (по 7 молекул с обеих сторон), наподобие нитки с бусинками, а также центры связывания ионов Са 2+. В этих центрах содержится особый белок (тропонин), участвующий в образовании связи актина с миозином.
Миозин составлен из уложенных параллельно белковых нитей (эта часть представляет собой так называемый легкий меромиозин). На обоих концах его имеются отходящие в стороны шейки с утолщениями – головками (эта часть – тяжелый меромиозин), благодаря которым образуются поперечные мостики между миозином и актином.
5.2. Механизмы сокращения и расслабления мышечного волокна
При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0,05 с (50 мс). За это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0,5 мс. Медиатором здесь является ацетилхолин, который содержится в синаптических пузырьках в пресинаптической части синапса. Нервный импульс вызывает перемещение синаптических пузырьков к пресинаптической мембране, их опорожнение и выход медиатора в синаптическую щель. Действие ацетилхолина на постсинаптическую мембрану чрезвычайно кратковременно, после чего он разрушается ацетилхолинзетеразой на уксусную кислоту и холин. По мере расходования запасы ацетилхолина постоянно пополняются путем его синтезирования в пресинаптической мембране. Однако при очень частой и длительной импульсации мотонейрона расход ацетилхолина превышает его пополнение, а также снижается чувствительность постсинаптической мембраны к его действию, в результате чего нарушается проведение возбуждения через нервно-мышечный синапс. Эти процессы лежат в основе периферических механизмов утомления при длительной и тяжелой мышечной работе.
Выделившийся в синаптическую щель медиатор прикрепляется к рецепторам постсинаптической мембраны и вызывает в ней явления деполяризации. Небольшое подпороговое раздражение вызывает лишь местное возбуждение небольшой амплитуды – потенциал концевой пластинки (ПКП).
При достаточной частоте нервных импульсов ПКП достигает порогового значения и на мышечной мембране развивается мышечный потенциал действия. Он (со скоростью 5 м/с) распространяется вдоль по поверхности мышечного волокна и заходит в поперечные трубочки внутрь волокна. Повышая проницаемость клеточных мембран, потенциал действия вызывает выход из цистерн и трубочек саркоплазматического ретикулума ионов Са2+, которые проникают в миофибриллы, к центрам связывания этих ионов на молекулах актина.
Под влиянием Са2+ длинные молекулы тропомиозина проворачиваются вдоль оси и скрываются в желобки между сферическими молекулами актина, открывая участки прикрепления головок миозина к актину. Тем самым между актином и миозином образуются так называемые поперечные мостики. При этом головки миозина совершают гребковые движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов саркомера к его центру, т. е. механическую реакцию мышечного волокна (рис. 10).
Энергия гребкового движения одного мостика производит перемещение на 1 % длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са2+. Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФазы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению имеющихся мостиков и образованию в присутствии Са2+ новых мостиков на следующем участке акт и позой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна – через 20 мс.
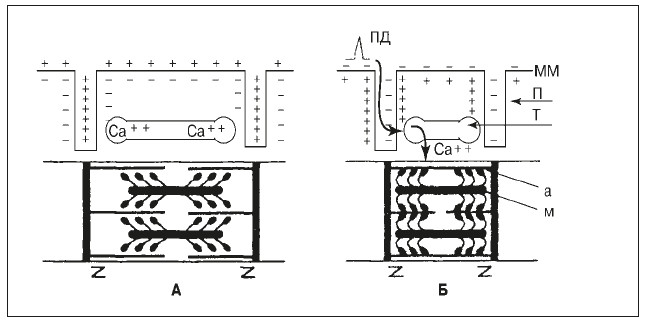
Рис. 10. Схема электромеханической связи в мышечном волокне.
А – состояние покоя; Б – возбуждение и сокращение; ПД – потенциал действия:
ММ – мембрана мышечного волокна; П – поперечные трубочки; I – продольные трубочки и цистерны с ионами Ca2+; а – тонкие нити актина, м – толстые нити миозина с утолщениями (головками) на концах. Зет-мембранами ограничены саркомеры миофибрилл. Толстые стрелки – распространение потенциала действия при возбуждении волокна и перемещение ионов Са2+ из цистерн и продольных трубочек в миофибриллы, где они содействуют образованию мостиков между нитями актина и миозина и скольжение этих нитей (сокращение волокна) за счет гребковых движений головок миозина
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма – «кальциевого насоса», который обеспечивает откачку ионов Са2+ из миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
5.3. Одиночное и тетаническое сокращение. Злектромиограмма
При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается одиночным сокращением. Эта форма механической реакции состоит из трех фаз: латентного, или скрытого, периода; фазы сокращения; фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1,5–2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
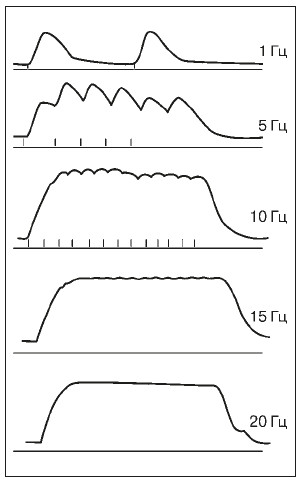
Рис. 11. Одиночное сокращение, зубчатый и сплошной тетанус камбаловидной мышцы человека
(по: Н.В. Зимкин и др., 1984); верхняя кривая – сокращение мышцы, нижняя – отметка раздражения мышцы, справа указана частота раздражения
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции – наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения – тетанус. Различают две формы тетануса – зубчатый тетанус, возникающий при более редком раздражении, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных одиночных сокращений, и сплошной, или гладкий, тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5–8 имп. в 1 с) возникают одиночные сокращения, при увеличении частоты (15–20 имп. в 1 с) – зубчатый тетанус, при дальнейшем нарастании частоты (25–60 имп. в 1 с) – гладкий тетанус. Одиночное сокращение – более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно (рис. 12), поддерживая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверхдлинные дистанции). При этом отдельные ДЕ могут развивать как одиночные, так и тетанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т. е. одновременное возбуждение практически всех ДЕ, что, в свою очередь, требует одновременной активации соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное тетаническое сокращение.
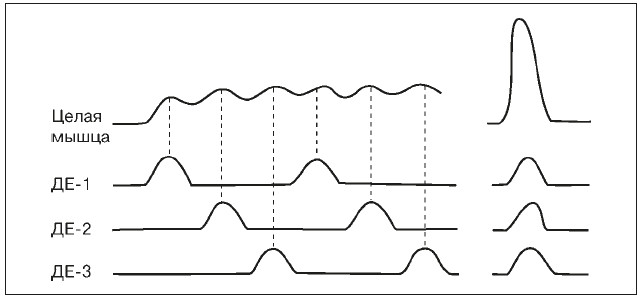
Рис. 12. Различные режимы работы двигательных единиц (ДЕ)
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон «Все или ничего»), В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа или с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных ДЕ. В случае же значительных напряжений потенциалы действия многих ДЕ. алгебраически суммируются, возникает сложная интегрированная кривая записи электрической активности целой мышцы – электромиограмма (ЭМГ).
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе – вид отдельных пачек импульсов, приуроченных в основном к начальному моменту сокращения мышцы и разделенных периодами «электрического молчания». Особенно хорошо ритмичность появления подобных пачек наблюдается у спортсменов при циклической работе (рис. 13). У маленьких детей и неадаптированных к такой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
Чем больше внешняя нагрузка и сила сокращения мышцы, тем выше амплитуда ее ЭМГ. Это связано с увеличением частоты нервных импульсов, вовлечением большего числа ДЕ в мышце и синхронизацией их активности. Современная многоканальная аппаратура позволяет производить одновременную регистрацию ЭМГ многих мышц на разных каналах. При выполнении спортсменом сложных движений можно видеть на полученных ЭМГ кривых не только характер активности отдельных мышц, но и оценить моменты и порядок их включения или выключения в различные фазы двигательных актов. Записи ЭМГ, полученные в естественных условиях двигательной деятельности, можно передавать к регистрирующей аппаратуре по телефону или радиотелеметрически. Анализ частоты, амплитуды и формы ЭМГ (например, с помощью специальных компьютерных программ) позволяет получить важную информацию об особенностях техники выполняемого спортивного упражнения и степени ее освоения обследуемым спортсменом.
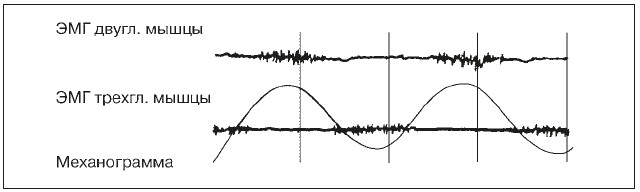
Рис. 13. Электромиограмма мышц-антагонистов при циклической работе
По мере развития утомления при той же величине мышечного усилия амплитуда ЭМГ нарастает. Это связано с тем, что снижение сократительной способности утомленных ДЕ компенсируется нервными центрами вовлечением в работу дополнительных ДЕ, т. е. путем увеличения количества активных мышечных волокон. Кроме того, усиливается синхронизация активности ДЕ, что также повышает амплитуду суммарной ЭМГ.
5.4. монофункциональные основы мышечной силы
Движение является результатом взаимодействия внутренних и внешних сил, развиваемых в опорно-двигательном аппарате, – активных (возникающих при сокращении или напряжении мышцы во время ее возбуждения) и пассивных (упругое напряжение при растяжении мышцы, сопротивление мышцы и ее сухожилия).
Сила мышцы зависит от ряда морфологических и физиологических факторов: количества и свойств мышечных волокон в мышце, исходной длины мышцы, характера нервных импульсов, механических условий действия мышцы на кости скелета.
Сила мышцы является суммой силы отдельных ее мышечных волокон. Подсчитано, что 1 одиночное мышечное волокно икроножной мышцы развивает напряжение 100–200 мг, 1 ДЕ икроножной мышцы человека содержит около 2000 мышечных волокон и развивает напряжение 200–400 г, 1 икроножная мышца содержит около 1000 ДЕ и развивает напряжение 200–400 кг.
Большое значение имеет анатомическое строение мышцы. В параллельно-волокнистых и веретенообразных мышцах (камбаловидная мышца и др.) сила мышц тем больше, чем больше ее анатомический поперечник, т. е. площадь поперечного сечения целой мышцы. В перистых мышцах (двуглавая мышца и др.) физиологический поперечник, т. е. площадь поперечного сечения всех мышечных волокон, гораздо больше, чем ее анатомический поперечник. В такой мышце упаковано значительно больше мышечных волокон и соответственно больше ее сила.
На силу сокращения мышцы влияет ее исходная длина, так как от нее зависит возможное количество поперечных мостиков между актином и миозином. Предполагают, что в каждом цикле присоединения-отсоединения поперечных мостиков расходуется энергия 1 молекулы АТФ на 1 поперечный мостик. Следовательно, чем больше образуется в мышечном волокне актино-миозиновых мостиков, тем выше скорость расщепления АТФ, больше тяга сократительных белков и соответственно больше развиваемая мышцей сила.
Наибольшее количество актино-миозиновых контактов образуется при небольшом растяжении мышцы до некоторой оптимальной длины. При значительном растяжении саркомера нити актина далеко расходятся в стороны и практически не контактируют с расположенным в средней части саркомера миозином. В случае же резкого уменьшения длины саркомера нити актина в центре перекрывают друг друга, препятствуя контактам с миозином и также уменьшая число образуемых мостиков. В связи с этими особенностями взаимодействия сократительных белков наибольшая сила мышцы проявляется при некотором ее предварительном растяжении.
Одной из важнейших характеристик скелетных мышц, влияющих на силу сокращения, является состав (композиция) мышечных волокон. Различают три типа мышечных волокон – медленные неутомляемые (I типа), быстрые неутомляемые или промежуточные (II-а типа) и быстрые утомляемые (II – б типа).
Медленные волокна (I типа), их обозначают также SO – Slow Oxydative (англ. – «медленные окислительные»), – это выносливые (неутомляемые) и легко возбудимые волокна, с богатым кровоснабжением, большим количеством митохондрий, запасов миоглобина и с использованием окислительных процессов энергообразования (аэробные). Их у человека в среднем 50 %. Они легко включаются в работу при малейших напряжениях мышц, очень выносливы, но не обладают достаточной силой. Чаще всего они используются при поддержании ненагрузочной статической работы, например при сохранении позы.
Быстрые утомляемые волокна (II-б типа), или FG – Fast Glicolitic (англ. – «быстрые гликолитические») – используют анаэробные процессы энергообразования (гликолиз). Они менее возбудимы, включаются при больших нагрузках и обеспечивают быстрые и мощные сокращения мышц. Зато эти волокна быстро утомляются. Их примерно 30 %. Волокна промежуточного типа (II-а) – быстрые неутомляемые, окислительные, их около 20 %. В среднем, для разных мышц характерно различное соотношение медленных неутомляемых и быстрых утомляемых волокон. Так, в трехглавой мышце плеча преобладают быстрые волокна (67 %) над медленными (33 %), что обеспечивает скоростно-силовые возможности этой мышцы (рис. 14), а для более медленной и выносливой камбаловидной мышцы характерно наличие 84 % медленных и всего 16 % быстрых волокон (Салтин Б., 1979).
Состав мышечных волокон в одной и той же мышце имеет огромные индивидуальные различия, зависящие от врожденных типологических особенностей человека. К моменту рождения человека его мышцы содержат лишь медленные волокна, но под влиянием нервной регуляции в ходе онтогенеза устанавливается генетически заданное индивидуальное соотношение мышечных волокон разного типа. По мере перехода от зрелого возраста к пожилому число быстрых волокон у человека заметно снижается и соответственно уменьшается мышечная сила. Например, наибольшее количество быстрых волокон в наружной головке четырехглавой мышцы бедра мужчины (около 59–63 %) отмечается в возрасте 20–40 лет, а в возрасте 60–65 лет их число почти на 1/3 меньше (45 %).
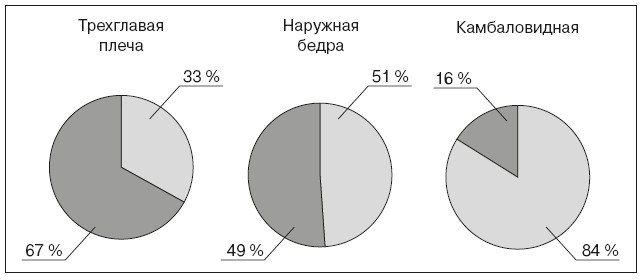
Рис. 14. Состав пышечных волокон в разных мышцах (медленные – светло-серым цветом; быстрые – темно-серым)
Количество тех или других мышечных волокон не изменяется в процессе тренировки. Возможно только нарастание толщины (гипертрофия) отдельных волокон, а также некоторое изменение свойств промежуточных волокон. При направленности тренировочного процесса на развитие силы происходит нарастание объема быстрых волокон, что и обеспечивает повышение силы тренируемых мышц.
Характер нервных импульсов изменяет силу сокращения мышц тремя способами:
1) увеличением числа активных ДЕ – это механизм вовлечения или рекрутирования ДЕ (сначала происходит вовлечение медленных и более возбудимых ДЕ, затем – высокопороговых быстрых ДЕ);
2) увеличением частоты нервных импульсов, в результате чего происходит переход от слабых одиночных сокращений к сильным тетаническим сокращениям мышечных волокон;
3) увеличением синхронизации ДЕ, при этом происходит увеличение силы сокращения целой мышцы за счет одновременной тяги всех активных мышечных волокон.
Существенное значение имеют механические условия работы мышцы – точка приложения ее силы и точка приложения сопротивления (поднимаемого груза). Например, при сгибании в локте вес поднимаемого груза может быть порядка 40 кг и более, при этом сила мышц-сгибателей достигает 250 кг, а тяга сухожилий – 500 кг.
Между силой и скоростью сокращения мышцы существует определенное соотношение, имеющее вид гиперболы (соотношение сила – скорость, по А. Хиллу). Чем выше сила, развиваемая мышцей, тем меньше скорость ее сокращения, и наоборот, с нарастанием скорости сокращения падает величина усилия. Наибольшую скорость развивает мышца, работающая без нагрузки. Скорость мышечного сокращения зависит от скорости передвижения поперечных мостиков, т. е. от частоты гребковых движений в единицу времени. В быстрых ДЕ эта частота выше, чем в медленных ДЕ, и соответственно потребляется больше энергии АТФ. Во время сокращения мышечных волокон в 1 с происходит примерно от 5 до 50 циклов прикрепления-отсоединения поперечных мостиков. При этом никаких колебаний силы в целой мышце не ощущается, так как ДЕ работают асинхронно. Лишь при утомлении возникает синхронная работа ДЕ, и в мышцах появляется дрожь (тремор утомления).
5.5. Режимы работы мышцы
Механическая работа (А), совершаемая мышцей, измеряется произведением поднимаемого веса (Р) на расстояние (h): А = P × h (им), При регистрации работы изолированной мышцы лягушки видно, что чем больше величина груза, тем меньше высота, на которую его поднимает мышца. Различают три режима работы мышцы: изотонический, изометрический и ауксотонический.
Изотонический режим (режим постоянного тонуса мышцы) наблюдается при отсутствии нагрузки на мышцу, когда мышца закреплена с одного конца и свободно сокращается. Напряжение в ней при этом не изменяется. Это происходит при раздражении изолированной мышцы лягушки, закрепленной одним концом на штативе. Так как при этих условиях Р = 0, то механическая работа мышцы также равна нулю (А = 0). В таком режиме работает в организме человека только одна мышца – мышца языка. (В современной литературе также встречается термин изотонический режим по отношению к такому сокращению мышцы с нагрузкой, при котором по мере изменения длины мышцы напряжение ее сохраняется неизменным, но в этом случае механическая работа мышцы не равна нулю, т. е. она совершает внешнюю работу.)
Изометрический режим (режим постоянной длины мышцы) характеризуется напряжением мышцы в условиях, когда она закреплена с обоих концов или когда мышца не может поднять слишком большой груз. При этом h = 0 и соответственно механическая работа тоже равна нулю (А = 0). Этот режим наблюдается при сохранении заданной позы и при выполнении статической работы. В этом случае в мышечном волокне все равно происходят процессы возникновения и разрушения мостиков между актином и миозином, т. е. тратится энергия на эти процессы, но отсутствует механическая реакция перемещения нитей актина вдоль миозина. Физиологическая характеристика такой работы заключается в оценке величины нагрузки и длительности работы.
Ауксотонический режим (смешанный режим) характеризуется изменением длины и тонуса мышцы, при сокращении которой происходит перемещение груза. В этом случае совершается механическая работа мышцы (А = P × h). Такой режим проявляется при выполнении динамической работы мышц даже при отсутствии внешнего груза, так как мышцы преодолевают силу тяжести, действующую на тело человека. Различают две разновидности этого режима работы мышц: преодолевающий (концентрический) и уступающий (эксцентрический) режим.
Изучение работы мышцы с различными нагрузками и в разном темпе позволило вывести закон средних нагрузок и среднего темпа движений: максимальную механическую работу мышца совершает при средних нагрузках и среднем темпе движений. При высоких скоростях сокращения мышцы часть ее энергии тратится на преодоление сопротивления (растущего внутреннего трения и вязкости мышцы), а при низких скоростях – на поддержание изометрического напряжения, которое также присутствует в этом случае для закрепления достигнутой длины мышцы в каждый данный момент времени.
Работу, производимую мышцами человека, изучают, используя различные методики ее регистрации – эргографию, велоэргометрию идр. В эргографии (греч. эр гон – «работа», графо – «писать») регистрируется амплитуда подъема различных грузов, подвешенных через блок. Вычисляя по эргограмме величину работы как произведение груза на амплитуду его подъема (А = P × h), И.М. Сеченов описал в 1905 г. явление активного отдыха. Оказалось, что пассивный отдых правой руки после ее утомления дает меньшее увеличение ее работоспособности, чем после работы (во время ее отдыха) левой руки.
5.6. Энергетика мышечного сокращения
При работе мышц химическая энергия превращается в механическую, т. е. мышца является химическим двигателем, а не тепловым. Для процессов сокращения и расслабления мышц потребляется энергия АТФ. Расщепление АТФ с отсоединением одной молекулы фосфата и образованием аденозиндифосфата (АДФ) сопровождается выделением 10 ккал энергии на 1 моль: АТФ = АДФ + Ф + Эн. Однако запасы АТФ в мышцах невелики (около 5 ммоль/л). Их хватает лишь на 1–2 секунды работы. Количество АТФ в мышцах не может изменяться, так как при отсутствии АТФ в мышцах развивается контрактура (не работает кальциевый насос и мышцы не в состоянии расслабляться), а при избытке – теряется эластичность.
Для продолжения работы требуется постоянное восполнение запасов АТФ. Восстановление АТФ происходит в анаэробных условиях – за счет распада креатинфосфата (КрФ) и глюкозы (реакции гликолиза) – и в аэробных условиях – за счет реакций окисления жиров и углеводов. Энергосистемы, используемые в качестве источников энергии, обозначают как фосфагенная энергетическая система (или система АТФ-КрФ), гликолитическая (или лактацидная) система и окислительная (или кислородная) система.
Быстрое восстановление АТФ происходит в тысячные доли секунды за счет распада КрФ: АДФ + КрФ = АТФ+ + Кр. Наибольшей эффективности этот путь энергообразования достигает к 5-6-й секунде работы, но затем запасы КрФ исчерпываются, так как их также немного (около 30 ммоль/л).
Медленное восстановление АТФ в анаэробных условиях обеспечивается энергией расщепления глюкозы (выделяемой из гликогена) – реакцией гликолиза с образованием в конечном итоге молочной кислоты (лактата) и восстановлением трех молекул АТФ. Эта реакция достигает наибольшей мощности к концу 1 – й минуты работы. Особое значение этот путь энергообразования имеет при высокой мощности работы, которая продолжается от 20 с до 1–2 мин (например, при беге на средние дистанции), а также при резком увеличении мощности более длительной и менее напряженной работы (спурты и финишные ускорения при беге на длинные дистанции) и при недостатке кислорода во время выполнения статической работы. Ограничение использования углеводов связано не с уменьшением запасов гликогена (глюкозы) в мышцах и в печени, а с угнетением реакции гликолиза избытком накопившейся в мышцах молочной кислоты.
Реакции окисления обеспечивают энергией работу мышц в условиях достаточного поступления в организм кислорода, т. е. при аэробной работе длительностью более 2–3 мин. Доставка кислорода достигает необходимого уровня после достаточного развертывания функций кислородтранспортных систем организма (дыхательной, сердечно-сосудистой систем и системы крови). Важным показателем мощности аэробных процессов является предельная величина поступления в организм кислорода за 1 мин – максимальное потребление кислорода (МПК). Эта величина зависит от индивидуальных возможностей каждого человека. У нетренированных лиц в 1 мин поступает к работающим мышцам около 2,5–3 л О, а у высококвалифицированных спортсменов (лыжников, пловцов, бегунов-стайеров и др.) достигает 5–6 л и даже 7 л в 1 мин.
При значительной мощности работы и огромной потребности при этом в кислороде основным субстратом окисления в большинстве спортивных упражнений являются углеводы, так как для их окисления требуется гораздо меньше кислорода, чем при окислении жиров. При использовании одной молекулы глюкозы (CjSL-O Д, полученной из гликогена, образуется 38 молекул АТФ, т. е. аэробный путь энергообразования обеспечивает при том же расходе углеводов во много раз больше продукции АТФ, чем анаэробный путь. Молочная кислота в этих реакциях не накапливается, а промежуточный продукт пировиноградная кислота сразу окисляется до конечных продуктов – СО, и Н,0.
В качестве источника энергии жиры используются в состоянии двигательного покоя, при любой работе сравнительно невысокой мощности (требующей до 50 % МПК) и при очень длительной работе на выносливость (требующей около 70–80 % МПК). Среди всех источников энергии жиры обладают наибольшей энергетической емкостью: при расходовании 1 моля АТФ выделяется около 10 ккал энергии, 1 моля КрФ – около 10,5 ккал, 1 моля глюкозы при анаэробном расщеплении – около 50 ккал, при окислении 1 моля глюкозы – около 700 ккал, при окислении 1 моля жиров – 2400 ккал (Код Я.М., 1982). Однако использование жиров при работе высокой мощности лимитируется трудностью доставки кислорода работающим тканям.
Работа мышц сопровождается выделением тепла. Теплообразование происходит в момент сокращения мышц – начальное теплообразование (оно составляет всего одну тысячную всех энерготрат) и в период восстановления – запаздывающее теплообразование.
В обычных условиях при работе мышц тепловые потери составляют около 80 % всех энерготрат. Для оценки эффективности механической работы мышцы используют вычисление коэффициента полезного действия (КПД). Величина КПД показывает, какая часть затрачиваемой энергии используется на выполнение механической работы мышцы.
Ее вычисляют по формуле:
КПД = [А : (Е – е)] × 100 %,
где А – энергия, затраченная на полезную работу; Е – общий расход энергии; е – расход энергии в состоянии покоя за время, равное длительности работы.
У нетренированного человека КПД примерно 20 %, у спортсмена – 30–35 %. При ходьбе наибольший КПД отмечается при скорости 3,6–4,8 км/ч, при педалировании на велоэргометре – при длительности цикла около 1 с. С увеличением мощности работы и включением «ненужных» мышц КПД уменьшается. При статической работе, поскольку А = 0, эффективность работы оценивается по длительности поддерживаемого напряжения мышц.
Назад: 4. Высшая нервная деятельность
Дальше: 6. Произвольные движения

