Книга: Мусорная ДНК. Путешествие в темную материю генома
Назад: Иногда все кончается хорошо
Дальше: Примечания
Глава 20. Луч света во тьме
Мы близимся к концу блужданий по темным закоулкам нашего генома. Внимательный читатель может заметить, что мы пока не рассмотрели тайну одного из заболеваний человека, впервые упомянутых еще в начале книги. Эта болезнь носит довольно неуклюжее название — плече-лопаточно-лицевая мышечная дистрофия (ПЛЛМД). При этом недуге постепенно тают мышцы лица, плеч и предплечий.
Болезнь возникает, когда будущий пациент наследует меньшее, чем нужно, количество повторов определенной нуклеотидной последовательности водной из копий хромосомы 4. Мутацию выявили еще несколько лет назад, но до самого последнего времени оставалось совершенно непонятным, почему она вызывает заболевание. Ведь рядом с этим генетическим дефектом, похоже, попросту нет никакого гена, кодирующего белки.
Но мы наконец-то обрели понимание того, каким образом возникают симптомы болезни. История впечатляющая. Она сводит воедино многие темы, с которыми мы уже встречались. Сейчас вы увидите, как ДНК, эпигенетика, генетические реликты и аномальный процессинг РНК осуществляют неслыханный патологический заговор1.
Небольшое напоминание. На каждой из нормальных копий хромосомы 4 указанная последовательность повторяется от 11 до 100 раз. Длина последовательности — чуть больше 3 тысяч пар нуклеотидных оснований. У больных ПЛЛМД число повторов меньше: на одной из копий хромосомы 4 у них встречается от одной до 10 таких последовательностей, расположенных подряд.
Здесь-то мы и сталкиваемся с первой сложностью. Есть люди, у которых число копий данного элемента ДНК составляет 10 или меньше, но при этом у них нет ПЛЛМД. Мышцы у них пребывают в совершенно здоровом состоянии. Малое количество повторов вызывает проблемы, лишь если оно наблюдается на копии хромосомы 4 и лишь если эта копия обладает особым свойством.
Чтобы осознать важность этого особого свойства, не помешает подробнее взглянуть на состав повторяющихся элементов. Оказывается, все они содержат некий ретроген. Ретроген — одна из форм мусорной ДНК. Он возникает, когда информационная РНК нормального гена клетки копируется обратно в ДНК и снова встраивается в геном. Во многом это напоминает процесс, показанный на рис. 4.1. Он происходил еще давным-давно, в нашем эволюционном прошлом.
Поскольку ретрогены изначально создаются на основе матриц информационной РНК, в них часто отсутствуют регуляторные последовательности, свойственные нормальным генам. Ретрогены не содержат сплайсинговых сигналов (поскольку матрица информационной РНК уже прошла сплайсинг перед копированием в ДНК). У них нет областей-промоторов и областей-энхансеров. Но некоторые из ретрогенов все-таки могут использоваться для создания информационной РНК. Именно так обстоит дело с ретрогеном ПЛЛМД. Однако обычно это не имеет значения, ибо такая РНК не функционирует в клетке должным образом. Она не содержит в себе нужных сигналов, побуждающих клетку добавлять цепочку оснований А к концу информационной РНК (этот процесс показан на рис. 16.5). Поэтому такая информационная РНК нестабильна и не может использоваться как матрица для производства белка.
Но если человек обладает лишь небольшим количеством повторов ПЛЛМД-элемента и если на хромосоме 4 имеются все другие свойственные ей последовательности, «окончательная» копия ПЛЛМД-ретрогена может подвергаться сплайсингу, порождая еще одну — дополнительную — последовательность. В результате на конце информационной РНК появляется сигнал, который позволяет клеточной аппаратуре пристраивать череду оснований А к этому концу. Такое добавление оснований А, в свою очередь, стабилизирует данную информационную РНК. Она преспокойно доставляется в рибосомы, где служит матрицей для производства белка — того самого белка, который никогда не должен экспрессироваться в зрелых мышечных клетках.
ПЛЛМД-белок регулирует экспрессию других генов, соединяясь с определенными ДНК-последовательностями. Сам он обычно экспрессируется только в генеративной, зародышевой линии — в половых клетках, которые производят яйцеклетки и сперматозоиды. Пока не существует четкого и убедительного объяснения, почему экспрессия данного белка вызывает увядание мышц. Возможно, тут задействован целый ряд механизмов. Может быть, этот белок активирует гены, провоцирующие отмирание мышечных клеток. Или приводит к потере стволовых клеток мышечной ткани — вероятно, посредством активирования других ретрогенов и захватчиков генома, которых следует держать в пассивном состоянии. Есть интригующая возможность: не исключено, что мышечные клетки, экспрессирующие ПЛЛМД-белок, разрушаются собственной иммунной системой пациента.
Генеративная линия — ткань, которая считается иммунологически привилегированной, поскольку обычно она изолирована от клеток нашей иммунной системы. А значит, нашей иммунной системе так никогда и не удается проведать о том, что клетки иммунологически привилегированных зон являются нормальной составной частью нашего организма. Если белки генеративной линии экспрессируются в зрелых мышечных клетках, иммунная система может отреагировать на них как на чужеродные организмы и атаковать клетки, экспрессирующие эти компоненты, с которыми иммунная система прежде никогда не сталкивалась.
Таким образом, ПЛЛМД показывает, какую важную роль играет мусорная ДНК при заболеваниях. Генетический дефект приводит к изменению некоторого количества мусорной ДНК. Вследствие этого некий мусорный элемент экспрессируется, а затем и модифицируется путем добавления к нему какой-то мусорной последовательности. Мало того: ПЛЛМД-ретроген стабильно экспрессируется лишь при определенной картине эпигенетических модификаций.
В нормальных клетках ПЛЛМД-повторы обычно экспрессируются, когда клетки находятся в плюрипотентном состоянии (в таком состоянии находятся, к примеру, эмбриональные стволовые клетки). На данной стадии ПЛЛМД-повторы покрыты активирующими эпигенетическими модификациями. Однако при дифференциации клеток активирующие модификации заменяются репрессивными и данная область отключается. Но если плюрипотентные клетки создаются в организме больного ПЛЛМД, активирующие модификации не заменяются при дифференциации клеток и область повторов остается включенной.
Еще один аспект — общий контроль генетической зоны, содержащей ПЛЛМД-ген. Между областью повторов и остальной частью хромосомы 4 находится область-изолятор (инсулятор). Белок 11-FINGER (мы с ним уже знакомы) связывается с инсулятором, обеспечивая иную картину эпигенетических модификаций в зоне ПЛЛМД по сравнению с соседними частями хромосомы.
Мало того, трехмерная структура этих областей хромосомы 4 также оказывает влияние на экспрессию ПЛЛМД-ретрогена. Почти наверняка именно сочетание всех перечисленных факторов приводит к той картине неполной атрофии мышц, которую наблюдают у больных ПЛЛМД.
Механизм, посредством которого при ПЛЛМД изменение в одной из областей мусорной ДНК приводит к болезни, являет собой ошеломляющий пример комплексных и многослойных взаимодействий различных элементов нашего генома. Он показывает, что нам не следует, размышляя о происходящем в наших клетках, представлять себе какие-то линейные пути и связи. Ведь речь идет о сложной системе взаимозависимых процессов. Рисунок 20.1 наглядно это демонстрирует. Теперь мы видим, почему так бессмысленны споры о том, какой компонент нашего генома является самым важным. Нарушение хотя бы какого-то его аспекта приводит к неким последствиям, одни из них окажутся серьезнее, чем другие, но все они действуют совместно.
Разумеется, это не значит, что среди миллиардов наших нуклеотидных пар решительно каждая обладает какой-то функцией. Возможно, некоторые действительно представляют собой бесполезный геномный мусор, тогда как другие области являются мусорными лишь в том смысле, что организм мог бы давно их выбросить, однако нашел им применение2.
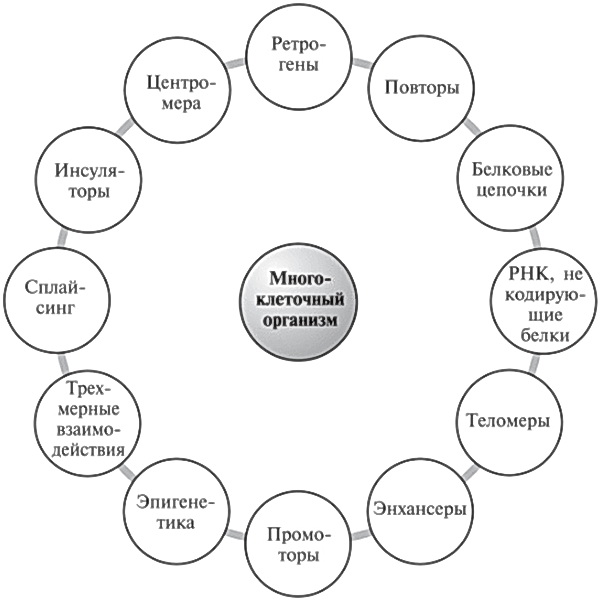
Рис. 20.1. Лишь некоторые из взаимодействующих факторов, которые должны действовать совместно, чтобы создать сложнейший организм, то есть нас с вами.
Мы еще многого не знаем. В частности, мы пока не получили ответов на целый ряд вопросов, которые могут показаться очень простыми и прямолинейными. К примеру, мы не знаем, сколько же функциональных областей мусорной ДНК существует в клетке. Думаете, ответ дать легко? Взгляните-ка на рис. 20.2 и побыстрее ответьте: сколько квадратов на шахматной доске?

Рис. 20.2. Скорее отвечайте: сколько квадратов на шахматной доске?
Все неизменно выдают мгновенное инстинктивное «64». На самом деле квадратов 204, поскольку на доске можно выстраивать более крупные квадраты всевозможных размеров, основываясь на более очевидных для нас элементах — черных и белых клетках. С нашим геномом такая же история. Один и тот же отрезок ДНК может включать в себя кодирующий белки ген, длинные некодирующие РНК, малые РНК, антисмысловые РНК, сайты сплайсингового сигнала, нетранслируемые области, промоторы и энхансеры. Прибавьте к этому дополнительные слои — те воздействия, которые оказывает различие в ДНК-последовательностях разных людей; направленные и случайные эпигенетические модификации; изменчивые трехмерные взаимодействия. Плюс связывание с другими РНК и белками. И прибавьте еще те влияния, причиной которых является постоянно меняющаяся среда, где мы обитаем.
Устройство нашего генома невероятно сложно, а потому вряд ли стоит удивляться, что мы пока не разобрались во всех его особенностях. Невероятным триумфом можно считать уже то, что мы понимаем хотя бы что-то. Там, в темной геномной материи, всегда найдется нечто еще не изученное.
Назад: Иногда все кончается хорошо
Дальше: Примечания

