Книга: Мусорная ДНК. Путешествие в темную материю генома
Назад: Эпигенетика и энхансеры: интенсивное общение
Дальше: От кустарного промысла до фабричного конвейера
Преодолевая расстояния
В большинстве событий, которые мы описывали в этой главе, участвуют энхансеры, которые находятся сравнительно близко от таргетируемых ими генов: обычно в пределах 50 тысяч нуклеотидных оснований. Довольно легко представить себе, как это происходит: длинная некодирующая РНК и комплекс-медиатор действуют как своего рода якорь для фермента, копирующего ДНК в информационную РНК. Однако во многих ситуациях энхансер и регулируемый им ген, кодирующий белок, на хромосоме отстоят друг от друга довольно далеко: их может разделять до нескольких миллионов пар нуклеотидных оснований. Такая же разница, как между попыткой передать солонку кому-то на противоположном конце стола и попыткой передать ее кому-то, кто располагается на другом конце футбольного поля. Нелегко представить себе, каким образом может осуществляться такого рода дистанционное взаимодействие между геном и энхансером. Размеров длинной некодирующей РНК и комплекса-медиатора не хватит, чтобы перекрыть такое огромное расстояние.
Чтобы разобраться в этом процессе, придется усложнить обычный образ генома. Как правило, весьма полезно и удобно описывать ДНК как веревочную лестницу или железнодорожные пути, ибо это помогает вообразить две ее нити и то, как они удерживаются вместе посредством пар нуклеотидных оснований. Однако проблема в том, что при этом мы думаем о ДНК как о чем-то линейном, к тому же достаточно жестком, поскольку подсознательно мы сравниваем ее с твердыми предметами из более привычной нам среды.
Но мы уже знаем, что ДНК — не жесткая молекула, ведь ее можно очень сильно сплющивать, сминать, комкать, чтобы она уместилась в ядре. Так что давайте углубим наши исследования. Если все-таки принять двунитевую природу ДНК как данность (чтобы не усложнять картину), можно представить себе фрагмент нашего генома как длинную макаронину — возможно, самый длинный в мире кусок лапши тальятелле. В двух местах она отмечена пищевым красителем: эти места изображают энхансер и ген, кодирующий белок. Глядя на рис. 12.3, мы видим два возможных сценария. Пока макароны не сварили, они сохраняют жесткость, и энхансер находится вдали от гена. Но после варки макароны становятся гибкими. Теперь они могут сгибаться во всевозможных направлениях, в результате чего окрашенные участки, изображающие энхансер и ген, могут сближаться.
Некоторые части наших хромосом в определенных клетках подавляются и выключаются почти навсегда — чтобы отключить гены, которые никогда не понадобится экспрессировать в тканях данного типа. Скажем, клеткам нашей кожи незачем экспрессировать белки, которые используются кровью для переноса кислорода. В клетках кожи соответствующие области генома совершенно недоступны: они плотно свернуты и напоминают чересчур сжатую пружину. Однако в этих же клетках есть громадные области, не находящиеся в таком сверхсжатом состоянии. Гены в них вполне доступны и могут включаться. ДНК в таких зонах напоминает самую длинную вареную макаронину в мире, целиком заполняющую кастрюлю. Макаронина сгибается и извивается в кипящей воде, образуя всевозможные петли и дуги.
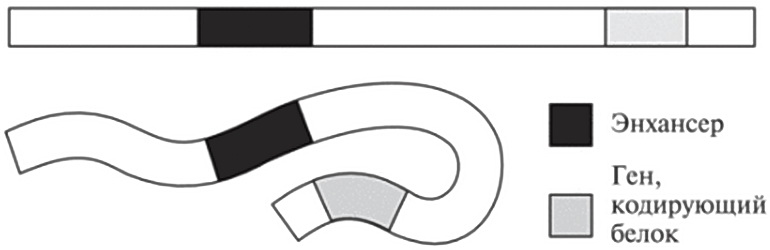
Рис. 12.3. Упрощенная схема показывает, как складывание гибкой молекулы ДНК может сблизить два отдаленных ее участка — скажем, энхансер и ген, кодирующий белок.
В таких случаях ген, кодирующий белок, и расположенный вдали от него энхансер могут сильно сближаться друг с другом. Затем длинная некодирующая РНК и комплекс-медиатор удерживают две петли вместе, обеспечивая усиление экспрессии гена. В этом медиатору должен помогать еще один комплекс. Этот дополнительный комплекс требуется также для разделения хромосом, прошедших дупликацию в процессе деления клетки, так что он хорошо оснащен для работы с масштабными перемещениями ДНК. Мутации в некоторых генах, кодирующих белки этого дополнительного комплекса, вызывают две болезни развития — синдром Робертса и синдром Корнелии де Ланге17. Их проявления могут быть довольно разными и, вероятно, определяются тем, какой именно ген мутирует и какова эта мутация. Обычно дети с этими заболеваниями рождаются слишком маленькими и в дальнейшем сохраняют это отставание в росте. Они испытывают трудности с обучением. Зачастую у них деформированы конечности18.
Подобный «петлевой» механизм распространен довольно широко. Возможно, он применим не только к энхансерам. Не исключено, что с его помощью и другие регуляторные элементы сближаются с теми или иными генами. В рамках исследования трех клеточных типов, затронувшего лишь 1% человеческого генома, удалось выявить более 1000 таких дистанционных взаимодействий в каждой линии клеток. Эти сложные взаимодействия чаше всего связывали области, разделенные примерно 120 тысячами пар нуклеотидных оснований. Среди более чем 90% таких петель ближайший ген просто игнорировался. Представьте, что вам понадобилось одолжить немного сахара, и вы отправляетесь не к ближайшему соседу, а к кому-нибудь, кто живет в полумиле от вас.
Продолжим тему соседей. Описываемые внутриклеточные взаимодействия ужасающе легкомысленны. Представьте себе свингерскую вечеринку 1970-х, только во много раз более разнузданную. Исследователи обнаружили, что у некоторых генов количество различных регуляторных областей, с которыми они взаимодействуют, доходит до 20. А у некоторых регуляторных областей количество генов, с которыми они взаимодействуют, доходит до десятка. Вероятно, не все эти взаимодействия происходят в одной и той же клетке в одно и то же время. Ученые показали главное: между генами и регуляторными регионами нет добропорядочных однозначных отношений «А и В». Нет, мы имеем дело со сложным набором взаимодействий, дающим клетке (или организму в целом) необычайно гибкие возможности регулирования всего этого пестрого узора генетической экспрессии19. Нам еще многое предстоит выяснить об этих сетях и о том, как они действуют. Сейчас дело выглядит так: мусорная ДНК, формирующая промоторы, заводит наши генетические моторы, но есть и мусорная ДНК, формирующая длинные некодирующие РНК и энхансеры. Она-то и превращает двигатель из слабенького сандеровского в такой, который вполне может разогнать «вейрон» на автостраде жизни.
Назад: Эпигенетика и энхансеры: интенсивное общение
Дальше: От кустарного промысла до фабричного конвейера

