Глава 9. Молекулы и карлики

К началу 1980-х годов мне стало очевидно, что без доступа на Мадагаскар моя карьера лемуролога долго не протянет. Я собрал все, что знал о предмете своего изучения, в одну книгу и опубликовал ее в 1982 году, а затем вернулся к палеоантропологии. Первым делом мне нужно было найти потенциальное место для раскопок, хотя очевидных вариантов у меня было совсем немного. Планируя свое возвращение в Нью-Йорк с острова Маврикий в Индийском океане, где я изучал обезьян, я обратил внимание на Джибути. Эта крошечная территория, когда-то принадлежавшая Франции, вдавалась в эфиопскую часть Афанского треугольника менее чем в 100 милях к востоку от Хадара. Насколько я мог судить по весьма неточным геологическим картам, находившимся в моем распоряжении, гряда осадочных пород, в которой были обнаружены великолепные афарские окаменелости, проходила как раз по территории недавно образованной Республики Джибути. На нее очевидно стоило взглянуть. Кроме того, мне все равно нужно было сделать там пересадку во время перелета с Маврикия в Париж, так что обстоятельства складывались в мою пользу. Я прибыл в дымный и гниющий портовый город Джибути и остановился в дешевом баре, он же отель, он же бордель, — единственном жилье, которое я мог позволить себе с моим бюджетом. В тот же вечер толпа пьяных матросов из Югославии совершила налет на это место. Они вламывались в комнаты с ножами и грабили постояльцев. Я был одним из немногих жителей отеля, которым удалось избежать ранений и больницы. Я помню, как провел целую вечность, изо всех сил удерживая на месте дверь в мой номер, дрожащую под яростными ударами снаружи, и как из соседних комнат доносились крики, удары и звон разбитого стекла.
Кто-то может сказать, что мне не хватало мотивации, но эта история поумерила мой энтузиазм относительно работы в Джибути. Более того, в итоге это место оказалось довольно бесперспективным. Через несколько лет французские ученые обнаружили здесь ископаемые останки животных эпохи плейстоцена, но единственным свидетельством присутствия гоминидов был фрагмент челюсти, который, по словам исследователей, «больше напоминал Homo sapiens, чем Homo erectus». Конечно, через некоторое время моя страсть к путешествиям вернулась и в итоге завела меня в такие далекие страны, как Йемен (дикое место с суровой красотой, где меня один раз чуть не похитили повстанцы, а еще один — едва не застрелили военные) и Вьетнам (пейзажи там были восхитительными, но вот окаменелости встречались редко). Но случай в Джибути заставил меня сделать вывод: важно не только находить новые окаменелости, но и правильно интерпретировать те, которые уже находились на музейных полках. Я очень четко понимал, что минималистический подход, унаследованный от Эрнста Майра, привел к недооценке разнообразия, присутствовавшего в палеонтологической летопись человечества, и, соответственно, к неправильному представлению о том, как мы стали людьми. По этой причине я начал размышлять об окаменелостях и о том, как именно палеоантропологи должны распознавать в них те или иные виды.
В то время мало кого интересовал этот вопрос. В конце концов, почти все ученые до сих пор верили, что виды — это всего лишь случайным образом выделенные сегменты эволюционных линий. Тем не менее точки зрения на сам эволюционный процесс различались, и в итоге мои коллеги оказались разделенными на два лагеря. К первому относились Милфорд Уолпофф, его соратники и ученики. Они быстро оправились от разочарования, пережитого в Восточной Туркане, и сформулировали обновленную версию гипотезы одного вида, которую вскоре начали называть мультирегиональной моделью. Основанная еще на довоенных идеях Франца Вайденрайха, эта модель объясняла современное географическое разнообразие Homo sapiens следующим образом: 1) основные географические группы человечества зародились в далеком прошлом, во времена яванского и пекинского человека, а различия между ними возникли в результате квазиизоляции; 2) при этом единство вида поддерживалось за счет обмена генами между географическими вариантами на границах регионов их обитания. В какой-то момент приверженцы этой теории поняли, что подобное объяснение означало бы включение в один вид нескольких эволюционных линий, но данная проблема была быстро решена — они попросту расширили свою концепцию и распространили ее на Homo erectus. Таким образом, редукционизм синтетической теории не только продолжал существовать, но и цвел буйным цветом.
Основным конкурентом мультирегиональной модели была гипотеза африканского происхождения человека, которую к тому моменту начали формулировать ученые из Англии и Германии. Они заявляли, что в палеоантологической летописи, какой бы скудной она ни была, первые указания на современную человеческую анатомию относятся к Африке. Вскоре у их гипотезы появилась неожиданная поддержка. В то время технологии молекулярной систематики постоянно улучшались и дорабатывались, и быстрее всего эти процессы проходили в лаборатории Алана Уилсона в Беркли, где работал Винсент Сарич. Ученым уже давно было известно, что физически гены состоят из последовательностей базовых элементов, расположенных в длинной цепочке ДНК. Таких базовых элементов насчитывается четыре — А, Т, Ц и Г. Расположенные в определенном порядке, эти «буквы» кодируют врожденные генетические инструкции, на основании которых строится новый организм. В 1970-х годах начали появляться технологии, позволяющие прочесть эти инструкции, и внимание ученых постепенно смещалось от белков (таких как альбумины, которые исследовали Сарич и Уилсон), кодируемых генами, к самой структуре ДНК.
Сравнив ДНК человека и шимпанзе, Уилсон и его студентка Мэри-Клер Кинг в 1975 году пришли к выводу, что структурные различия в их ДНК, в частности в генах, кодирующих белки, были слишком незначительными для объяснения существенных анатомических различий. Соответственно, они должны были объясняться разницей в активности генов в процессе развития. Это было весьма дальновидное и крайне важное заключение, подтвержденное более поздними исследованиями: ключевыми факторами эволюции являлись изменения в регуляции активности генов и их экспрессии. Данные факторы имеют огромное значение для процесса развития. В свою очередь, встречающиеся в избытке «факторы транскрипции», которые и активируют деятельность генов, кодирующих белки, почти наверняка могут помочь заполнить многочисленные анатомические пробелы в палеонтологической летописи. Вполне вероятно, что такие пробелы включают в себя внезапные появления современных анатомических черт (таких как у мальчика из Нариокотоме) и в результате возникновение анатомически уникальных Homo sapiens.
Вскоре Уилсон и его сотрудники сконцентрировались на ДНК определенного типа. Большая часть ДНК хранится в ядрах наших клеток, но небольшая доля находится в митохондриях — одном из типов органелл, расположенных в цитоплазме клетки между ядром и мембраной и отвечающих за клеточное функционирование. В митохондриях содержится гораздо меньше ДНК, чем в ядре, но клетка имеет несколько митохондрий, а значит, одна и так же молекула ДНК дублируется в ней несколько раз. Так называемая мтДНК имеет огромный потенциал для молекулярной систематики. Большая часть ядерной ДНК перемешивается после слияния яйцеклетки и сперматозоида, поэтому проследить ее между поколениями невозможно. Митохондриальная же ДНК дает четкое представление о наследственности, потому что она передается без всяких изменений по материнской линии (сперматозоиды отца, по сути, представляют собой всего лишь ядра клеток). Соответственно, любые различия в мтДНК между двумя особями, когда-то имевшими общего предка, объясняются исключительно мутациями (то есть спонтанными изменениями, постоянно возникающими в четырех возможных типах нуклеотидов).
В 1987 году Уилсон и его студенты Ребекка Кэнн и Марк Стоункинг исследовали мтДНК 147 человек из разных регионов мира. Предположив, что скорость изменчивости должна быть постоянной, и приняв во внимание количество различий между образцами, они рассчитали, что наш вид возник около 200,000 лет назад. Более того, полученное ими «дерево мтДНК» указывало на происхождение Homo sapiens с африканского континента. Это подтверждалось и тем фактом, что потомки жителей Африки имеют наибольшее разнообразие мтДНК, то есть люди живут и развиваются здесь дольше, чем где бы то ни было еще. Относительно точной даты возникновения Homo sapiens в молекулярной систематике ведутся жаркие споры, но два факта несомненны. Во-первых, наш вид появился относительно недавно, а во-вторых, через какое-то время после возникновения он прошел через «бутылочное горлышко», то есть существенное сокращение популяции, после чего покинул африканский континент.
Меня, естественно, очень привлекали эти открытия, и не только потому, что они подтверждали мои собственные размышления о палеонтологической летописи. Молекулярный подход Уилсона и его коллег указывал на то, что, даже если наш вид еще не достиг конца своего развития, оно по крайней мере имело четкое начало. Невозможно было заниматься молекулярной систематикой, не признавая, что виды действительно существуют, а такое признание давало возможность для более тонкого анализа палеонтологической летописи человечества. Но, чтобы научное сообщество могло это осознать, требовалось время. Должен признаться, в тот момент меня больше всего радовало, что неандертальцев с их уникальными характеристиками наконец-то признают отдельным видом, а не просто вымершей разновидностью Homo sapiens.
Я полагал, что неандертальцы существенно отличались от нас не только с анатомической, но и с поведенческой точки зрения, что подтверждалось археологическими находками. Мне казалось, что, отрицая уникальность этих далеких родственников человека, палеоантропологи отказывались от возможности понять их в их собственном контексте, как отдельный вид, которым они почти наверняка и являлись. В конце концов, мне надоело ждать, пока эксперты по неандертальцам разберутся с этой проблемой, и в 1986 году я написал работу, посвященную трудностям классификации окаменелостей по их морфологии. В ней подчеркивалось, что видообразование как процесс, результатом которого является возникновение новых биологических единиц, способных к размножению, очевидно, не пассивное последствие морфологических изменений. Одни виды накапливают множество таких изменений, но не разделяются на несколько разных частей, а другие, наоборот, едва различаются на вид, но при этом совершенно несовместимы в репродуктивном плане. Эти явные различия между морфологией и статусом вида не играют большой роли, когда у вас есть возможность наблюдать за взаимодействием (или его отсутствием) живых существ. Но вот если особи, которых вы анализируете, живут вдали друг от друга, или же если у вас имеются только их окаменелые кости, такие различия становятся крайне важным фактором.
Практический опыт (в том числе и моя работа с лемурами) показывал, что зачастую родственные виды приматов не имеют значительных различий в морфологии костей и зубов. Таким образом, если вы обнаруживаете в одном раскопе две явно различающиеся окаменелости твердых тканей, высока вероятность, что перед вами останки двух разных видов, разумеется, если вы сможете исключить половой диморфизм. Более того, так как различия между костями родственных видов обычно незначительны, палеоантропологи легко могут недооценить количество видов, представленных тем или иным собранием окаменелостей. Разумеется, мои рассуждения вводили некоторый элемент неуверенности, но даже он был предпочтительнее, чем излишнее упрощение эволюционных паттернов вместо их классификации. С другой стороны, если я был прав, то все попытки выделить на основании окаменелостей подвиды, то есть четко выраженные региональные варианты, возникающие внутри видов и быстро исчезающие в них же, были обречены на провал. Подвиды — это движущая сила эволюционных инноваций, основа для формирования новых видов. Но до сих пор, пока процесс видообразования не наделит их исторической индивидуальностью, они остаются эфемерными.
Практические трудности, связанные с распознаванием близко-родственных видов в палеонтологической летописи, никуда не делись с 1986 года, когда моя работа была опубликована. Я приложил все усилия, но так и не сумел найти способ, как их устранить. Тем не менее положительным результатом моей публикации стало то, что в последние десятилетия палеоантропологи все реже и реже употребляют термин «стадия». Стадия — это особая палеоантропологическая концепция, которую я однажды назвал самой ужасной «уткой», выдуманной моими коллегами. Стадией называют неопределенную группу окаменелостей, имеющих примерное сходство (например, по объему мозга). Такая группировка, разумеется, является продуктом человеческого сознания, а не природы. Стадии не существуют сами по себе, это всего лишь эвристические инструменты, которые тем не менее иногда путают с реальностью. Сам по себе этот термин не так уж плох, но если палеоантрополог мыслит «стадиями», он забывает о естественной классификации, созданной самой природой. Если кто-то говорит, что эволюция гоминидов прошла через «стадию Homo erectus», то подразумевается, что в ней отсутствуют такие понятия, как предки и наследование. Вместо этого всю работу выполнил естественный отбор, и дальнейшие исследования не имеют смысла. Придерживаясь идеи «стадии», мы проигнорируем тот факт, что за последние пару миллионов лет рост объема мозга в роду Homo происходил независимо в рамках трех разных линий развития: у Homo erectus в Восточной Азии, у предков неандертальцев в Европе и у предков Homo sapiens в Африке. Это очень важно, так как если рост мозга является свойством всего рода Homo в целом, а не конкретно нашего вида, то и корни этого явления нужно искать совершенно в другом месте.
В 1986 году моей главной задачей было показать, что Homo sapiens представляет собой во многом уникальный вид и что лишь некоторые окаменелости (имеющие сравнительно молодой возраст) вписываются в его морфологическую структуру. В палеонтологической летописи плейстоцена отсутствует и большая круглая черепная коробка, и достаточно маленькое лицо в ее нижней части, характерные для современного человека. Таким же уникальным является и Homo neanderthalensis с его длинным списком характеристик, не похожих ни на чьи другие (близкие родственники неандертальцев из Сима-де-лос-Уэсос к тому моменту еще не были обнаружены).
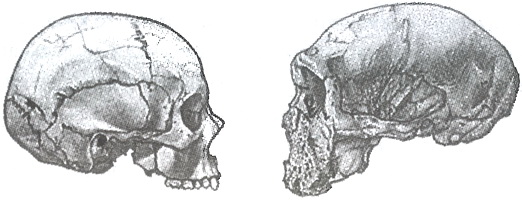
Различия в строении черепа между современным Homo sapiens (Эль-Хеса, Египет, слева) и Homo neanderthalensis (Гротта-Гуаттари, Монте-Сирсео, Италия). В масштабе. Рисунки Дона Макгрэнэгана (слева) и Дианы Саллес (справа)
Я не сомневался, что Homo sapiens и Homo neanderthalensis являлись двумя отдельными видовыми единицами, которые к моменту своего первого контакта уже давно двигались по собственным эволюционным траекториям. Мне казалось, что, включая неандертальцев в вид Homo sapiens в качестве странного и по определению низшего подвида, мы лишаем их индивидуальности. Тем не менее многие палеоантропологи твердо придерживались и придерживаются этого мнения. До сих пор существует научная школа, считающая, что во времена плейстоцена Земля была населена гибридами неандертальцев и современных людей. И не просто считающая, а проповедующая свои идеи с религиозным пылом. Я очень четко помню, как некоторое время назад один мой коллега поносил меня на весь Интернет за комментарий, который я считал крайне дипломатичным. Я просто призвал ученых, интерпретировавших недавно найденный скелет подростка в возрасте 24,5 тысяч лет как гибрид неандертальцев и Homo sapiens, к осторожности в высказываниях, а в итоге чувствовал себя еретиком, за которым пришла инквизиция. Стоит ли говорить, что этот подросток жил как минимум через 200 поколений после того, как вымерли последние неандертальцы!
Но включение неандертальцев в вид Homo sapiens не только ущемляло их идентичность. Оно скрывало и нашу собственную анатомическую уникальность, равно как и принятая среди моих англоязычных коллег классификация черепов из Кабве, Петралоны, Араго и Бодо в качестве «архаичных Homo sapiens». Что касается ученых из других стран мира, то они, признавая отличие этих окаменелостей от нашего собственного строения, относили их к Homo erectus. Такая интерпретация никем не оспаривалась, так как большинство все все равно полагало, что Homo erectus были предками Homo sapiens.
Лично я предпочитал относить останки из Кабве, Петралоны, Араго и Бодо к Homo heidelbergensis — виду, который Отто Шетензак выделил на основании челюсти из Мауэра (как оказалось впоследствии, ее строение и зубные характеристики были похожи на те, что обнаружились у челюстей из Араго). Но даже в этом случае некоторые находки времен позднего плейстоцена, например окаменелости из Лаэтоли и Ндуту, оставались в неопределенном таксономическом положении. Сложности в их классификации еще раз доказывали большое таксономическое разнообразие известных нам гоминидов. С какой стороны ни посмотри, на него указывали все базовые признаки окаменелостей. В своей работе я указал на то, что именно этого и следовало ожидать, учитывая нестабильные природные условия плейстоцена. Частые климатические изменения регулярно разбивали популяции гоминидов на более мелкие группы, создавая оптимальные условия для возникновения и закрепления эволюционных изменений. После смягчения климата разрозненные группы могли снова соединяться вместе. Таким образом, изменения оставались в популяции и приводили к возникновению конкуренции.
Назад: Исход
Дальше: Во всем виновата среда

