Книга: Ружья, микробы и сталь. История человеческих сообществ
Назад: Глава 9 Зебры, несчастливые браки и принцип "Анны Карениной"
Дальше: Часть 3 От продовольствия ружьям, микробам и стали
Глава 10
Просторные небеса и наклонные оси
Взгляните на карту мира юл и сравните очертания и ориентацию разных континентов. Поразительно, насколько очевидна разница. Протяженность Америки с севера на юг значительно больше (9000 миль), чем с востока на запад (3000 миль в самом широком месте, сужающемся в районе Панамского перешейка всего лишь до 40). Иначе говоря, преобладающая ориентация Америки – “вертикальная”, по оси север-юг. То же самое, хотя в значительно менее выраженной степени, верно для Африки. Напротив, у Евразии главная ось – восток-запад. Имела ли эта разница в ориентации континентов какие-то последствия для истории человечества, и если да, то какие?
Я полагаю, что ее последствия были колоссальными, даже подчас трагическими, и именно им будет посвящена эта глава. Ориентация осей континентов повлияла на скорость распространения растительных культур и домашнего скота, а также, возможно, письменности, колесного транспорта и других изобретений. Тем самым один из основных географических параметров сыграл важнейшую роль в том, как сложились очень разные исторические судьбы евразийцев, африканцев и коренных американцев за последние 500 лет.
* * *
Для понимания географической неравномерности возникновения ружей, микробов и стали характер распространения производства продовольствия, как оказывается, имеет не менее принципиальное значение, чем рассмотренные нами в предыдущих главах место и время его происхождения. Регионов, где производство продовольствия зародилось самостоятельно, на планете было максимум девять, а может быть, и не больше пяти (см. главу 5). Однако уже в доисторический период помимо пяти-девяти независимых очагов оно успело укорениться во многих других регионах. Сельскохозяйственная история этих последних – результат распространения культур, домашнего скота, навыков их выращивания, а в некоторых случаях и экспансии самих земледельцев и скотоводов.
Главные направления распространения производства продовольствия были следующими: из Юго-Западной Азии в Европу, Египет и Северную Африку, Эфиопию, Центральную Азию и долину Инда; из Сахеля и Западной Африки в Восточную и Южную Африку; из Китая в тропическую Юго-Восточную Азию, на Филиппины, в Индонезию, Корею и Японию; из Мезоамерики в Северную Америку. Кроме того, производство продовольствия в самих очаговых регионах нередко обогащалось за счет культур, видов скота и сельскохозяйственных технологий, заимствуемых ими друг у друга.
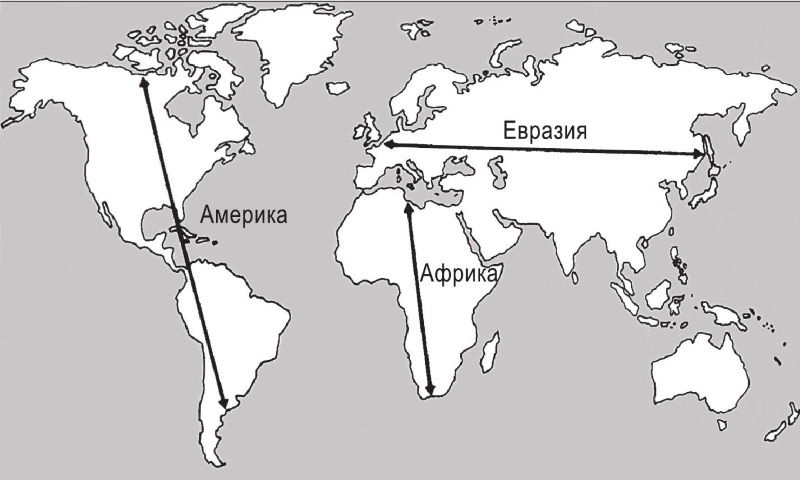
Рис. 10.1. Оси континентов.
Точно так же, как условия в одних местах гораздо больше благоприятствовали зарождению продовольственного производства, чем в других, существенно варьировались и условия его распространения. В некоторых регионах, экологически прекрасно подходящих для сельского хозяйства, в доисторические времена оно так и не появилось – несмотря на соседство регионов, где оно уже было. Вот наиболее яркие примеры: земледелие и животноводство не проникло ни с юго-запада США в Калифорнию (в доколумбов период), ни с островов Индонезии и Новой Гвинеи в Австралию; земледелие не проникло из южноафриканской провинции Наталь в соседнюю Капскую область. Но даже в те многочисленные регионы, до которых производство продовольствия дошло еще в древности, оно прибывало с разной скоростью и в разное время. На одном краю этого диапазона – быстрая миграция по оси восток-запад: из Юго-Западной Азии на запад – в Европу и Египет – и на восток – в долину Инда (с приблизительной средней скоростью 0,7 миль в год), а также с Филиппин на восток – на острова Полинезии (3,2 мили в год). На противоположном краю – медленное продвижение по оси север-юг: менее 0,5 мили в год для северной экспансии мексиканских культур на юго-запад США; менее 0,3 мили в год для проникновения мексиканской кукурузы и фасоли на восток Соединенных Штатов (здесь их начали активно возделывать только в 900 г. н. э.); 0,2 мили в год для миграции лам из Перу в Эквадор. Эти отличия могут быть еще более разительными, если отсчитывать возделывание культурной кукурузы в Мексике не от 3500 г. до н. э. (осторожная оценка некоторых современных археологов, взятая мной для расчета), а от какой-нибудь гораздо более ранней даты (что было бы верно с точки зрения большинства вчерашних и многих сегодняшних ученых).
Сильно варьировалось и то, в каком составе сельскохозяйственные комплексы (наборы растительных культур и домашних животных) усваивались на новых местах. Этот разброс снова указывает на наличие более и менее преодолимых препятствий. Например, если большинство первичных доместикатов Юго-Западной Азии легко перекочевало на запад в Европу и на восток в долину Инда, то из двух одомашненных в Андах млекопитающих (ламы/альпаки и морской свинки) в Мезоамерику в доколумбовы времена не попало ни одно. Этот поразительный факт мы просто не можем оставить без объяснения. В конце концов, в Мезоамерике существовали густонаселенные развитые аграрные общества, и при ином развитии событий андские домашние животные несомненно имели бы для них огромную ценность как источник пищи, шерсти и средство транспортировки – особенно учитывая, что, за исключением собак, в Мезоамерике совершенно отсутствовали домашние млекопитающие, способные удовлетворить эти потребности. В то же время несколько растительных культур Южной Америки – маниок, батат, арахис – каким-то образом сумели распространиться на север. Какой барьер мог избирательно пропустить эти культуры, но преградить дорогу ламам и морским свинкам?
Другим, менее наглядным выражением географического торможения сельского хозяйства является феномен, который называется упреждающей доместикацией. Большинство диких видов, из которых происходят культурные растения, в разных регионах имеют несколько отличающийся генотип. Это происходит потому, что среди древних популяций этих видов в каждом конкретном регионе закрепились разные мутации. Аналогичным образом, биологические изменения, которые сопровождают превращение дикого растения в культуру, в принципе могут быть вызваны отличающимися последовательностями мутаций или, иначе говоря, альтернативными траекториями отбора, которые дают равноценные результаты. Благодаря этому можно взять культурное растение, широко распространенное в доисторические времена, и проверить, обнаруживают ли все его разновидности одну и ту же дикую мутацию или одну и ту же трансформационную мутацию. Цель такой проверки – попытаться узнать, была ли выведена данная культура только в одном регионе или независимо в нескольких.
Если провести генетический анализ основных культур Нового Света, многие из них покажут наличие следов двух и более альтернативных диких вариантов или двух и более альтернативных трансформационных мутаций. Из этого, скорее всего, следует, что культура была самостоятельно выведена как минимум в двух разных регионах и что некоторые разновидности унаследовали мутацию одного региона, а другие разновидности той же культуры – мутацию другого. Именно так ботаники пришли к выводу, что лимская фасоль (.Phaseolus lunatus), обыкновенная фасоль (.Phaseolus vulgaris), два вида красного перца (Capsicum аппиит и Capsicum chinense) все были независимо окультурены по меньшей мере дважды – один раз в Мезоамерике и еще раз в Южной Америке. То же самое относится к тыкве обыкновенной (Cucurbita реро) и семяносному растению марь, только в данном случае их двумя родинами были Мезоамерика и восток США. Напротив, большинство древних культур Юго-Западной Азии обнаруживают только одну дикую или трансформационную мутацию, из чего следует, что все современные разновидности каждой из этих культур восходят к одной первичной доместикации.
О чем говорит факт независимого выведения одной и той же культуры в разных областях ареала ее дикого предка – в отличие от однократного в пределах одной территории? Как мы знаем, окультуривание подразумевает модификацию диких растений с целью увеличения их полезности для человека: получения семян большего размера, устранения горького вкуса и т. д. Поэтому, если первые земледельцы уже имели в своем распоряжении достаточно урожайную культуру, они определенно предпочитали выращивать ее, а не начинать новый цикл: собирать урожаи родственного, но менее ценного дикого растения и повторно его одомашнивать. Соответственно факты, устанавливающие однократность доместикации, указывают на то, что после выведения данной культуры она быстро распространилась по остальным частям ареала ее дикого предшественника и тем самым упредила необходимость повторной доместикации того же самого растения. В противном случае, когда мы находим свидетельства независимого одомашнивания одного растения в разных регионах, мы делаем вывод, что первичная культура распространялась слишком медленно, чтобы опередить выведение ее аналогов в других местах. Таким образом, данные, говорящие о том, что в Юго-Западной Азии преобладали однократные доместикации, а в Америке дело ими часто не ограничивалось, можно рассматривать как еще одно, чуть менее наглядное доказательство того факта, что природные условия первой были более благоприятными для миграции культур.
Стремительное распространение некоей культуры способно упредить доместикацию не только того же самого дикого вида-предка, но и родственных ему. Если вы уже выращиваете урожайный горох, будет явно нецелесообразно не только начинать с нуля и выводить новую культуру из того же дикого вида, но и выводить ее из близких родственников – то есть получать растение, с точки зрения потребителя фактически равноценное уже имеющемуся одомашненному гороху. Первые основные культуры Юго-Западной Азии сделали ненужной доместикацию всех своих близких родственников на всей территории Западной Евразии. Напротив, Новый Свет знает немало примеров, когда равноценные и близкородственные – но не идентичные – виды одомашнивались дважды: в Мезоамерике и Южной Америке. К примеру, 95 % хлопчатника, выращиваемого сегодня в мире, принадлежат к виду Gossypium hirsutum, одомашненному в доисторический период в Мезоамерике. Между тем доисторические земледельцы Южной Америки возделывали свой собственный вид того же рода – Gossypium barbadense. Очевидно, мезоамериканскому хлопчатнику было настолько трудно попасть в Южную Америку, что в доисторические времена он не смог опередить на этом континенте доместикацию своего родственника (и наоборот). Красный перец, тыква, амарант, марь – вот еще несколько растений, разные виды которых были окультурены в Мезоамерике и Южной Америке из-за того, что темпы распространения ни одного из них не позволяли сделать ненужным окультуривание их родственников.
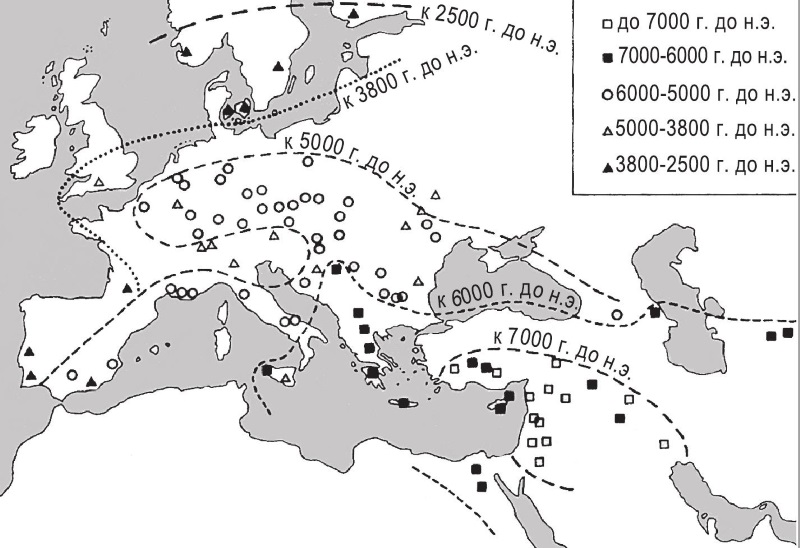
Рис. 10.2. Распространение сельскохозяйственных культур Плодородного полумесяца в западной Европе. Символами обозначены места, в которых были обнаружены остатки культур Плодородного полумесяца с наиболее ранней радиоуглеродной датировкой. ▄ = сам Плодородный полумесяц (стоянки до 7000 г. до н. э.) Обратите внимание, что по мере удаления от Плодородного полумесяца стоянки относятся ко все более поздним датам. Карта основана на данных карты 20 из книги “Доместикация растений в Старом Свете” Зохари и Хопф, но их неоткалиброванные датировки заменены откалиброванными.
Таким образом, несколько разных исторических феноменов подталкивают нас к одному и тому же выводу: производство продовольствия распространялось из Юго-Западной Азии куда активнее, чем внутри двух Американских континентов, а возможно, и активнее, чем в Африке к югу от Сахары. Об этом, в первую очередь, говорит совершенное отсутствие производства продовольствия в некоторых экологически благоприятных регионах, во-вторых, разница в скорости и избирательности его распространения, в-третьих, упреждающая роль наиболее ранних одомашненных культур в одних случаях и повторное одомашнивание идентичных или близкородственных видов – в других. Благодаря какой особенности Америки и Африки распространение производства продовольствия на их территории было более затрудненным, чем в Евразии?
* * *
Чтобы ответить на этот вопрос, проследим внимательней за тем, как именно проходила стремительная экспансия производства продовольствия из Юго-Западной Азии (Плодородного полумесяца). Вскоре после рождения местного сельского хозяйства, примерно в конце IX тысячелетия до н. э., его центробежная волна постепенно стала докатываться до других частей Западной Евразии и Северной Африки, все более и более удаленных от Плодородного полумесяца на запад и на восток. На рисунке ю.2 вы видите несколько переиначенную мной наглядную карту, которую составили генетик Дэниел Зохари и ботаник Мария Хопф. Карта показывает, что центробежная экспансия захватила Грецию, Кипр и Индийский субконтинент к 6500 г. до н. э., Египет – вскоре после 6000 г. до н. э., Центральную Европу – к 5400 г. до н. э., Южную Испанию – к 5200 г. до н. э. и Британские острова – около 3500 г. до н. э. В каждом из этих регионов начало производящей экономике было положено той или иной группой одомашненных растений и животных из ближневосточного набора “основателей”. Кроме того – хотя достоверно не установлено, когда именно, – ближневосточный набор частично проник в более южные районы Африки и достиг Эфиопии. Но поскольку в Эфиопии было выведено довольно много собственных культур, мы не можем точно сказать, чему обязан старт местного производства продовольствия – этим аборигенным доместикатам или доместикатам Плодородного полумесяца.
Разумеется, не все элементы ближневосточного набора распространились во всех названных регионах: к примеру, слишком жаркий климат Египта не позволил выращивать здесь пшеницу-однозернянку. В некоторые удаленные от очага регионы разные элементы попадали в разное время: так, в Юго-Западной Европе овцеводство было освоено значительно раньше возделывания хлебных злаков. В некоторых из них также были получены собственные культуры: в Западной Европе – мак, в Египте (предположительно) – арбузы. Однако основной объем производства продовольствия в заимствующих регионах все-таки изначально обеспечивался за счет доместикатов из Плодородного полумесяца. За культурами и видами скота вскоре последовали другие новшества из того же источника: колесо, письменность, металлургия, доение молочных животных, разведение плодовых деревьев, пивоварение и виноделие.
Почему один и тот же набор растений лег в основу производства продовольствия на всей территории Западной Евразии? По той ли причине, что одни и те же растения встречались в диком виде во множестве регионов, были оценены по достоинству их жителями так же, как жителями Плодородного полумесяца, и впоследствии подверглись независимому окультуриванию? Нет, причина не в этом. Во-первых, многие культуры Плодородного полумесяца за пределами Юго-Западной Азии в диком виде не произрастают вообще. Например, в Египте, за исключением ячменя, ни один из диких предков восьми ближневосточных первых основных культур не встречается. В то же время египетская долина Нила имеет экологические условия, аналогичные условиям ближневосточных долин Тигра и Евфрата, и поэтому земледельческий комплекс, доказавший свою эффективность в двух последних, прижился здесь настолько хорошо, что спровоцировал расцвет самобытной египетской цивилизации. Однако растения, буквально подпитывавшие этот яркий исторический феномен, изначально в Египте отсутствовали. Сфинксы и пирамиды были построены людьми, чью пищу составляли культуры не египетского, а ближневосточного происхождения.
Во-вторых, даже если взять те культуры Европы и Индии, чей дикий ареал простирается за пределы Юго-Западной Азии, в большинстве случаев мы можем быть уверены, что имеем дело с заимствованием, а не с локальной доместикацией. Например, дикий лен распространен от Британии и Алжира на западе до Каспийского моря на востоке, а ареал дикого ячменя на востоке доходит до самого Тибета. Однако почти у всех возделываемых сегодня в мире сортов ближневосточных первых основных культур присутствует один и тот же набор хромосом, несмотря на многообразие таких наборов у их прародителей. В оставшихся случаях это носители единственной из множества возможных мутаций, закрепляющих ценные для человека признаки и отличающих культурные разновидности от дикого предка. К примеру, все культивируемые сорта гороха содержат один и тот же рецессивный ген, не дающий спелым стручкам самопроизвольно раскрываться и высыпать зерна в почву, как это происходит у дикого гороха.
Очевидно, что после первичной доместикации на территории Плодородного полумесяца большинство исходных основных культур никто нигде больше не одомашнивал. Если бы повторная независимая доместикация имела место, эти культуры обнаружили бы следы своих нескольких родословных в виде варьирующегося набора хромосом или варьирующихся мутаций. Следовательно, они являют собой типичный пример феномена упреждающей доместикации, о котором мы говорили выше. Скорое распространение ближневосточного комплекса культур сделало ненужными любые другие попытки – внутри или за пределами Плодородного полумесяца – одомашнить те же самые растения. Как только люди получали доступ к данной культуре, нужда собирать ее в диком виде, то есть делать первые шаги к новой доместикации, исчезала.
У предков большинства первых основных культур есть дикие родственники на Ближнем Востоке и не только, которые пригодны к одомашниванию ничуть не меньше. Скажем, горох относится к роду Pisum, состоящему из двух видов: Pisum sativum, из которого и был получен наш посевной горох, и Pisum fulvum, который остался диким. Меду тем зерна Pisum fulvum, как свежесобранные, так и засушенные, приятны на вкус, а само растение встречается в природе довольно часто. Аналогично пшеница, ячмень, чечевица, нут, фасоль и лен – все они помимо своих непосредственных предков имеют много других диких родственников. Более того, некоторые из таких родственников фасоли и ячменя были самостоятельно окультурены в Америке и Китае – вдалеке от места первичной доместикации в Плодородном полумесяце. Однако в Западной Евразии из нескольких потенциально ценных диких видов одомашнен был только один – видимо, потому, что стремительность распространения культурного варианта позволила людям бросить собирание его диких родственников и питаться только им. Опять, как и сказано выше, быстрое распространение культуры опередило любые возможные попытки доместикации родственных растений или повторной доместикации ее собственного дикого предка.
* * *
Почему экспансия культур из Плодородного полумесяца происходила с такой скоростью? Ответ частично содержится в указании на широтную ориентацию Евразии, с которого я начал эту главу. Местности, расположенные к востоку и западу друг от друга на одной широте, имеют в точности ту же самую сезонную амплитуду продолжительности светлого времени суток. В несколько меньшей, но тем не менее значительной степени они сходны и по другим параметрам: характерным заболеваниям растений, режимам температуры и осадков, биомам (типам растительности). Так, Португалия, север Ирана и Япония – местности, расположенные примерно на одной широте, но последовательно отстоящие друг от друга на 4000 миль, – больше похожи друг на друга по климатическим условиям, чем каждая из них похожа на местность, находящуюся всего лишь в 1000 миль к югу от нее. Тип биома, называемый влажным тропическим лесом, на всех континентах ограничен десятью широтными градусами вокруг экватора, а средиземноморский вечнозеленый кустарник (например, калифорнийский чапараль или европейский маквис) произрастает в поясе между тридцатью и сорока градусами.
Между тем всхожесть, характер роста и сопротивляемость болезням у растений адаптированы как раз под эти климатические особенности. Сезонные изменения длины дня, температуры, количества осадков становятся сигналами, которые стимулируют прорастание семян, рост саженцев, цветение, образование семян и плодов у зрелых растений. Каждая популяция посредством естественного отбора становится запрограммированной на адекватную реакцию при получении сезонных сигналов, в условиях которых протекала ее эволюция. Эти условия значительно меняются в зависимости от широты. Например, длина дня – круглогодичная постоянная величина на экваторе, но в субтропических и тем более умеренных широтах она растет от зимнего к летнему солнцестоянию и уменьшается оставшуюся половину года. Период вегетации – то есть месяцы с температурой и продолжительностью дня, пригодными для роста растений, – короче всего в высоких широтах и длиннее всего на экваторе. Помимо этого, любое растение адаптируется к преобладающим на его широте болезням.
Горе тому растению, чья генетическая программа не знает широты, на которой его посадили! Представьте себе канадского фермера, которого угораздило посадить сорт кукурузы, адаптированный к природным условиям Мексики. Это несчастное растение, послушное своей “мексиканской” генетической программе, приготовится взойти в марте – и окажется под 10 футами снега. Даже если бы оно было перепрограммировано на прорастание в более подходящий для Канады срок – скажем, в конце июня, – у него всего равно оставалась бы масса других проблем. Его гены подсказывали бы ему не торопиться с ростом, поскольку до наступления зрелости у него в запасе целых пять месяцев. И если для мексиканского теплого климата такая стратегия совершенно оправданна, в Канаде она гарантированно погубит растение, потому что осенние заморозки здесь наступят скорее, чем у него сформируется полноценный початок. У этого несчастливца также не будет иммунитета к болезням северного климата, зато будут бесполезные гены, напрасно ждущие нападения южных инфекций. Из-за множества таких нюансов растения низких широт редко выживают в высоких широтах и наоборот. Соответственно, большинство культур Плодородного полумесяца нормально растут во Франции и Японии, но практически не способны расти на экваторе.
Животные тоже приспосабливаются к широтным особенностям климата. В этом отношении мы – типичные животные, и мы прекрасно это знаем. Одни из нас не переносят холодных северных зим с их коротким днем и специфическими инфекциями, другие – жаркий тропический климат и тропические болезни. За последние несколько столетий переселенцы из прохладной Северной Европы предпочитали оседать в регионах с похожим умеренным климатом – в Северной Америке, в Австралии, на юге Африки – или в прохладных высокогорных районах экваториального пояса – например, в Кении и на Новой Гвинее. Североевропейцы, которых посылали в жаркие низменные страны тропического пояса, в прошлом сотнями и тысячами умирали от болезней вроде малярии, к действию которых у коренного населения имелась некоторая наследственная устойчивость.
Частично именно в этом и заключается объяснение столь стремительной западной и восточной миграции доместикатов Плодородного полумесяца: они были уже достаточно адаптированы к климату регионов-реципиентов. Например, после того как примерно в 5400 г. до н. э. земледелие проникло с венгерских равнин в Центральную Европу, оно распространилось так быстро, что стоянки первых земледельцев на огромной территории от Польши до Голландии (отличающиеся особым типом керамики с линейным орнаментом) имеют практически совпадающие датировки. К началу нашей эры хлебные культуры ближневосточного происхождения произрастали в поясе длиной больше 8000 миль: от атлантического побережья Ирландии до тихоокеанского побережья Японии. Протяженность Евразии с запада на восток – самое большое сухопутное расстояние на Земле.
Итак, западно-восточная ориентация Евразии позволила культурам Плодородного полумесяца за короткое время дать старт сельскому хозяйству в субтропическо-умеренной полосе от Ирландии до долины Инда, а также обогатить сельское хозяйство, самостоятельно возникшее в Восточной Азии. Точно так же евразийские культуры, впервые выведенные вдалеке от Плодородного полумесяца, но в тех же самых широтах, были способны мигрировать в направлении, обратном распространению первых основных культур. Сегодня, когда зерно перевозится по всему земному шару океанским и воздушным транспортом, для нас само собой разумеется видеть на своем столе настоящий продовольственный интернационал. В типичном наборе из американского фастфуда мы встретим курицу (впервые одомашненную в Китае) с картошкой (выведенной в Андах) или кукурузой (выведенной в Мексике), которые мы приправляем черным перцем (индийского происхождения) и запиваем кофе (эфиопского происхождения). Но еще две тысячи лет назад у древних римлян имелся собственный смешанный набор продуктов, родиной которых, как правило, была совсем не Италия. Из римских культур только овес и мак являлись аборигенами Апеннинского полуострова, основой же римского рациона служил ближневосточный комплекс “основателей”, дополненный айвой (кавказского происхождения), просом и тмином (одомашненными в Центральной Азии), огурцами, кунжутом и цитрусовыми (из Индии), а также курицей, рисом, абрикосами, персиками и могаром (из Китая). И хотя яблоки, которые тоже присутствовали на римском столе, были результатом селекции садоводов Западной Евразии, они выращивались с помощью методики прививания, которая впервые появилась в Китае и лишь затем распространилась на запад.
Евразия включает в себя самый протяженный в мире одноширотный пояс суши и поэтому служит наиболее ярким примером быстрой миграции доместикатов. Однако этот пример не единственный. Соперничать по скорости с экспансией ближневосточного комплекса может восточная экспансия субтропического комплекса, изначально сложившегося в Южном Китае и пополнившегося позднее в тропической Юго-Восточной Азии, Индонезии, на Филиппинах и Новой Гвинее. За тысячу шестьсот лет результирующий набор культур (в который входили бананы, таро и ямс) и домашних животных (куры, свиньи и собаки) преодолел больше 5000 миль на восток, распространившись благодаря полинезийцам на огромном пространстве Тихого океана. Еще один пример того же рода – восточно-западная миграция культур внутри широкой африканской зоны Сахель, однако ее конкретные обстоятельства палеоботаникам еще только предстоит выяснить.
* * *
Сравните легкость, с которой происходила восточно-западная миграция доместикатов в Евразии, с тем, насколько затрудненной была их миграция в Африке вдоль меридианальной оси. Большинство первых основных культур Плодородного полумесяца достигло Египта за довольно короткий срок, после чего продолжило экспансию на юг, дойдя до высокогорья Эфиопии с его субтропическим климатом – и здесь остановилось. Южноафриканский средиземноморский климат идеально подходил для них, однако 2000 миль тропиков между Эфиопией и Южной Африкой оказались непреодолимым барьером. Вместо них в основание сельского хозяйства к югу от Сахары легли окультуренные дикие растения (в частности, сорго и африканский ямс), которые являлись аборигенами Сахеля и тропической Западной Африки и которые были приспособлены к высоким температурам, летним ливням и относительно постоянной продолжительности дня, характерным для этих низких широт.
Сходным образом южная миграция ближневосточного домашнего скота в Африке была полностью или частично приостановлена климатическими условиями и тропическими заболеваниями, особенно переносимым мухами цеце трипаносомозом. Лошади так и не прижились на континенте южнее западноафриканских королевств, расположенных к северу от экватора. Продвижение крупного рогатого скота, овец и коз затормозилось на северных границах равнины Серенгети на две тысячи лет, понадобившихся для развития новых форм хозяйствования и новых пород животных. В Южную Африку вся эта живность попала лишь в первые два столетия нашей эры – примерно через восемь тысяч лет после своего одомашнивания в регионе Плодородного полумесяца. Свои трудности были и у южной миграции тропических африканских культур, которые проникли на юг континента вместе с африканскими земледельцами банту сразу вслед за ближневосточным домашним скотом. Несмотря на это, они так и не смогли преодолеть южноафриканскую реку Фиш – естественный рубеж средиземноморской зоны, к условиям которой они не были адаптированы.
Результатом стала хорошо нам понятная очередность событий последних двух тысячелетий южноафриканской истории. Некоторые из коренных койсанских народов (иначе известных как готтентоты и бушмены) освоили разведение скота, но не приняли земледелие. К северо-востоку от реки Фиш их постепенно сменили численно превосходящие африканские аграрные племена, южная экспансия которых как раз у этой реки остановилась. Только когда в 1642 г. на юг Африки по морю прибыли европейские колонисты, завезшие ближневосточный набор культур, в местной средиземноморской зоне наконец появилось свое земледелие. Именно столкновениям между всеми этими народами Южная Африка обязана трагедиями своей современной истории: стремительным и резким сокращением численности койсанских народов, причинами которого были европейские патогены и огнестрельное оружие; столетием войн между европейцами и африканцами; еще одним столетием расового угнетения; препятствиями, которые стоят сегодня на пути белых и черных, ищущих способ ужиться друг с другом на бывших койсанских землях.
* * *
Теперь сравните легкость распространения доместикатов в Евразии с трудностями их миграции вдоль меридианальной оси двух Американских континентов. Расстояние между Мезоамерикой и Южной Америкой – конкретно, допустим, между мексиканским высокогорьем и Эквадором – составляет всего лишь 1200 миль, то есть примерно столько, сколько в Евразии отделяет Балканы от Месопотамии. На Балканах имелись идеальные условия для выращивания большинства месопотамских культур и домашних животных, и местное население получило доступ к этому сельскохозяйственному комплексу в пределах двух тысячелетий после его формирования в регионе Плодородного полумесяца. Столь скорое проникновение сделало ненужным доместикацию тех же самых и родственных видов на Балканах. Высокогорная Мексика и Анды сходным образом были бы вполне пригодны для многих культур и домашних животных друг друга. Некоторые культуры, главной из которых была мексиканская кукуруза, и в самом деле сумели проделать путь между двумя регионами в доколумбову эпоху.
Однако другие культуры и домашние животные не смогли преодолеть расстояние между Мезоамерикой и Южной Америкой. Прохладный климат мексиканского высокогорья сделал бы их идеальным местом для выращивания лам, морских свинок и картофеля, которые все были одомашнены в прохладном климате Анд. Однако на пути северной миграции этих андских доместикатов непроходимым препятствием стали низменные территории Центральной Америки. На протяжении пяти тысячелетий после одомашнивания ламы в Андах ольмекам, майя, ацтекам и другим коренным народам Мексики пришлось существовать без вьючных животных и без мяса каких-либо других домашних млекопитающих, кроме собак.
Точно так же разводимые мексиканцами индейки и окультуренный на востоке Соединенных Штатов подсолнечник замечательно прижились бы в Андах, однако их южная миграция была остановлена разделяющими эти регионы тропиками. Какие-то 700 миль по оси север-юг мешали мексиканской кукурузе, тыквам и фасоли достичь юго-запада США на протяжении нескольких тысячелетий после их окультуривания в Мексике, а мексиканские виды красного перца и семейства маревых так и не преодолели это расстояние в доколумбову эпоху. Тысячелетия после первичной доместикации кукурузы в Мексике она не могла распространиться севернее, на восток Северной Америки, из-за преобладающего здесь более прохладного климата и более короткого периода вегетации. Где-то между началом и 200 г. н. э. кукуруза наконец появилась на востоке США, но лишь в качестве второстепенной культуры. Лишь после 900 г., когда уже были выведены зимостойкие сорта кукурузы, адаптированные к северному климату, началось интенсивное возделывание кукурузы, которое со временем обеспечило расцвет самого сложного из коренных североамериканских обществ – так называемой миссисипской культуры, недолговечный расцвет которой был прерван появлением европейских патогенов, завезенных Колумбом и его последователями.
Вспомните, что у большинства культур Плодородного полумесяца генетический анализ выявляет наличие единственной родословной – после одомашнивания эти виды распространились с такой скоростью, что любая возможная доместикация их же или их родственников была прервана еще в зародыше. Наоборот, многие аборигенные культуры Америки, на первый взгляд оккупировавшие огромную территорию, как оказывается, представляют собой несколько родственных видов или генетически отличных разновидностей одного вида, независимо одомашненных в Мезоамерике, Южной Америке и на востоке Соединенных Штатов. Близкородственные виды сменяют друг друга от региона к региону у амарантовых, бобовых, маревых, красных перцев, хлопчатника, тыкв и табака. Разновидности одного вида сменяют друг друга у фасоли обыкновенной, фасоли лимской, красного перца Capsicum annuum/chinense и тыквы Cucurbito реро. Это наследие множественных независимых доместикаций может послужить еще одним свидетельством медленного распространения культур по американской оси север-юг.
Итак, Африка и Америка представляют собой два крупнейших массива суши с меридианальной ориентацией и, следовательно, медленной диффузией культур. В некоторых других частях мира медленная миграция по оси север-юг тоже сыграла важную, пусть и не такую масштабную роль, из примеров можно назвать прогрессировавший буквально черепашьим шагом взаимообмен культурами между долиной Инда (территория нынешнего Пакистана) и Южной Индией, долгое проникновение южнокитайского земледелия на Малайский полуостров и непроникновение доисторического тропического земледелия Индонезии и Новой Гвинеи на юго-запад и юго-восток Австралии соответственно. Эти уголки Австралийского континента сегодня являются двумя его главными житницами, однако они находятся в 1000 милях от экватора, и поэтому рождение местного земледелия произошло лишь после того, как из далекой Европы на европейских судах сюда прибыли культуры, адаптированные к европейскому прохладному климату и короткому вегетационному периоду.
* * *
Я уделил так много внимания географической широте – фактору, устанавливаемому простым взглядом на карту, – потому что именно она является главной определяющей климата, условий вегетации и легкости распространения производства продовольствия. Однако широта, разумеется, не единственная такая определяющая, и не всегда бывает так, что соседи по широте имеют один и тот же климат (хотя они всегда имеют одну и ту же продолжительность светлого времени суток). Ландшафтные и экологические барьеры, гораздо более выраженные на одних континентах, чем на других, локально тоже иногда оказывались важной помехой для распространения сельского хозяйства.
Так, например, взаимообмен растительными культурами между юго-востоком и юго-западом США был крайне неторопливым и избирательным, хотя оба региона находятся на одной широте. Причиной такого положения дел являлась разделяющая их территория Техаса и южной части Великих равнин, засушливый климат которой не благоприятствовал земледелию. Аналогичный пример из истории Евразии – задержка распространения культур Плодородного полумесяца на территории Индийского субконтинента. Несмотря на то что они быстро и практически беспрепятственно достигли атлантического побережья на западе и долины Инда на востоке, восточнее Инда смена преимущественно зимнего режима годовых осадков на преимущественно летний способствовала тому, что на северо-востоке Индии, в бассейне Ганга, земледелие развивалось гораздо медленней, с использованием других культур и аграрных приемов. Области субтропического климата, находившиеся еще восточнее, на территории Китая, были отделены от западноевразийских регионов с аналогичным климатом пустынями Центральной Азии, Тибетским нагорьем и Гималаями. Таким образом, первоначальное развитие производства продовольствия в Китае происходило независимо от его развития на той же широте, в регионе Плодородного полумесяца, и привело к появлению совершенно иных культур. Тем не менее даже эти барьеры между Китаем и Западной Евразией были как минимум частично преодолены в течение и тысячелетия до н. э., когда западноазиатские пшеница, ячмень и лошадь достигли Китая.
Точно так же степень преодолимости расстояния в 1000 миль по оси север-юг иногда зависела от местных особенностей. Ближневосточное производство продовольствия мигрировало на юг, до самой Эфиопии, а бантуское – из региона Великих озер до южноафриканской провинции Наталь – в обоих случаях как раз на это расстояние, – потому что промежуточные области имели сходный сезонный режим осадков и были пригодны для земледелия. Напротив, миграция культур Индонезии на юго-запад Австралии оказалась вовсе невозможной, а миграция из Мексики на значительно менее удаленные юго-запад и юго-восток США – очень медленной, поскольку в обоих случаях промежуток был заполнен враждебными земледелию пустынями. Отсутствие возвышенных плато южнее Гватемалы и радикальное сужение Мезоамерики к Панамскому перешейку по меньшей мере не уступали по значимости широтному фактору, создавая эффект бутылочного горлышка и затормаживая обмен культурами и домашними животными между Андами и высокогорными районами Мексики.
Различия в ориентации осей континентов повлияли на распространение не только производства продовольствия, но и других технологий и инноваций. Например, колесо, изобретенное около 3000 г. до н. э. в самой Юго-Западной Азии или ее окрестностях, за несколько столетий распространилось на запад и восток по значительной территории Евразии, а то же самое изобретение, самостоятельно сделанное в доисторической Мексике, до Анд так и не дошло. Аналогично если принцип алфавитного письма, придуманный в западной части Плодородного полумесяца не позже 1500 г. до н. э., был освоен к западу до Карфагена и к востоку до Индийского субконтинента уже через тысячу лет, то о мезоамериканской письменности жители Анд не имели сведений все две тысячи с лишним лет ее доисторического существования.
Само собой, колесо и письменность в отличие от растительных культур не привязаны к широте и продолжительности дня. Однако определенная зависимость работает и здесь, главным образом через посредство сельского хозяйства и результатов его освоения. Первые колеса были элементом запряженных быками повозок, использовавшихся для транспортировки земледельческой продукции. Первая письменность была прерогативой элит, живущих за счет крестьян-производителей, и она служила целям экономически и социально сложных аграрных обществ (монархической пропаганде, учету запасов, ведению административных записей и т. п.). В общем и целом общества, вовлеченные в обмен культурными растениями, домашними животными и технологиями, так или иначе связанными с производством продовольствия, с большей вероятностью вступали в обменные отношения и в других сферах человеческой деятельности.
В патриотической песне “Америка прекрасная” поется о наших просторных небесах, о волнующихся янтарных хлебах – от моря и до моря. Географическая реальность выглядела иначе. Как и африканские, небеса Нового Света были довольно тесны для распространения собственных культур и домашних животных, на пути которых к тому же стояли мощные экологические барьеры. Никакой хлебный пояс не связывал ни атлантическое побережье Северной Америки с тихоокеанским, ни Канаду с Патагонией, ни Египет с Южной Африкой – зато янтарные поля пшеницы и ячменя волновались от атлантического до тихоокеанского побережья под просторными небесами Евразии. Тот факт, что в Евразии сельское хозяйство распространялось куда быстрее, чем в Америке или Африке, сыграл свою роль (мы увидим это в следующей части книги) в ускоренной экспансии евразийской письменности, металлургии, технологий и имперских государств.
Акцентировать внимание на этой разнице не значит вынести суждение о превосходстве растительных культур, завоевавших сегодня огромные территории планеты, или как-то превознести находчивость и смекалку первых евразийских земледельцев. Нет, источник этой разницы – достоинства осевой ориентации Евразии по сравнению с ориентацией обеих Америк и Африки. На этих осях вращались колеса исторических судеб континента.
Назад: Глава 9 Зебры, несчастливые браки и принцип "Анны Карениной"
Дальше: Часть 3 От продовольствия ружьям, микробам и стали

