Книга: Лекции по общей психологии
Назад: ГЛАВА 1. Психология как наука. Ее предмет и практическое значение
Дальше: Глава 3. Сознательная деятельность человека и ее общественно — исторические корни
Глава 2. Эволюция психики
Мы остановились на том, как современная наука понимает предмет психологии и какие практические приложения имеет эта область знания.
Теперь нам следует осветить одну из важнейших проблем — эволюцию психической деятельности.
Происхождение психики
Донаучная психология, которая развивалась в ранней идеалистической философии, считала психику одним из первичных свойств человека и рассматривала сознание как непосредственное проявление «духовной жизни». Поэтому вопрос о естественных корнях психики, о ее происхождении и о ступенях ее эволюции даже не ставился. Дуалистическая философия предполагала, что сознание так же вечно, как и материя, что оно всегда существовало параллельно с материей.
Научная психология исходит из совершенно иных положений и ставит перед собой задачу подойти к ответу на вопрос о происхождении психики, описать условия, в результате которых должна была появиться эта сложнейшая форма жизни.
Известно, что основным условием появления жизни является возникновение сложных белковых молекул, которые не могут существовать без постоянного обмена веществ со средой. Для своего выживания они должны усваивать (ассимилировать) из окружающей среды те вещества, которые являются предметом питания и необходимы для поддержания их жизни; одновременно они должны выделять во внешнюю среду продукты распада, усвоение которых может нарушить их нормальное существование. Оба эти процесса — ассимиляция и диссимиляция — входят в процесс обмена веществ и являются основным условием существования этих сложных белковых образований.
Естественно, что эти сложнейшие белковые молекулы (иногда их называют «коацерватами») вырабатывают особые свойства, отвечающие на воздействие полезных веществ или тех условий, которые содействуют усвоению этих веществ, и на вредные воздействия, грозящие их дальнейшему существованию. Так, эти молекулы положительно реагируют не только на питательные вещества, но и на такие условия, как свет, тепло, которые содействуют усвоению. Они отрицательно реагируют на сверхсильные механические или химические воздействия, которые мешают их нормальному существованию. На «нейтральные» воздействия, не входящие в процесс обмена веществ, они не реагируют.
Свойство коацерватов реагировать на воздействия, входящие в процесс обмена веществ (оставляя без ответа посторонние «индифферентные» воздействия), называется раздражимостью. Это основное свойство проявляется при переходе от неорганической материи к органической. К нему присоединяется и второе свойство — возможность сохранять высокоспециализированные свойства раздражимости к воздействиям, передавая соответствующие модификации белковых молекул от одного поколения к другому. Это последнее свойство, по — видимому связанное с модификацией некоторых фракций аминокислот (в частности, рибонуклеиновой кислоты, или РНК, составляющей молекулярную основу жизни), принято рассматривать как важный процесс, лежащий в основе биологической памяти.
Процессы раздражимости по отношению к жизненно важным «биотическим» воздействиям, выработка высокоспециализированных форм раздражимости и сохранение их с передачей последующим поколениям, характеризует ту стадию развития жизни, которую обычно обозначают как растительная жизнь.
Этими процессами характеризуется вся жизнь, начиная от простейших водорослей и кончая сложными формами растительной жизни. Ими же обусловлены и так называемые «движения растений», которые, по существу, являются лишь формами усиленного обмена или роста, направляемого раздражимостью по отношению к биотическим воздействиям (влажности, освещенности и т. п.). Такие явления как рост корня растения вглубь почвы, или неравномерный рост ствола в зависимости от освещенности, или поворот растения в направлении солнечных лучей — все это является лишь результатом явлений «раздражимости» к биотическим (небезразличным для жизни) воздействиям.
Существенным для растительной жизни является одно важное обстоятельство. Растение, реагирующее усиленным обменом на биотические воздействия, не реагирует на посторонние воздействия, которые входят в процесс непосредственного обмена веществ. Оно не ориентируется активно в окружающей среде и может, например, погибнуть от отсутствия света или влаги, даже если источники света и влаги существуют совсем близко, но не оказывают на него непосредственного воздействия.
От этой пассивной формы жизнедеятельности резко отличаются формы существования на следующем этапе эволюции — на стадии животной жизни.
Характерным для каждого животного организма, начиная с простейших, является тот основной факт, что животное реагирует не только на биотические воздействия, непосредственно входящие в процесс обмена веществ, но и на «нейтральные», небиотические воздействия, если только они сигнализируют о появлении жизненно важных («биотических») воздействий. Иначе говоря, животные (даже простейшие) активно ориентируются в условиях среды, ищут жизненно важные условия и реагируют на всякие изменения среды, которые являются сигналом появления таких условий. Чем интенсивнее протекает обмен веществ, чем большую потребность испытывает простейшее живое существо в получении пищи, тем более активны его движения, тем в более оживленных формах протекает его «ориентировочная», или «поисковая», деятельность.
Эта способность реагировать на нейтральные «абиотические» раздражения при условии, что они сигнализируют о появлении жизненно важных воздействий, появляющаяся на стадии перехода к животному миру, называется, в отличие от явлений раздражимости, чувствительностью. Появление чувствительности и может служить объективным биологическим признаком возникновения психики.
Изменчивость поведения простейших
Чувствительность по отношению к «нейтральным» раздражителям, если они начинают сигнализировать о появлении жизненно важных воздействий, вызывает коренные изменения в формах жизни. Главное заключается в том, что живое существо начинает «ориентироваться» в окружающей среде, активно реагировать на каждое изменение, происходящее в ней, т. е. вырабатывать индивидуально изменчивые формы поведения, которые не существовали в растительном мире.
На первых порах выработка такого индивидуально меняющегося поведения происходит относительно медленно, однако его удается наблюдать даже в условиях эксперимента.
Приведем один из типичных экспериментов, проведенных немецким исследователем Брамштедтом.
Известно, что одноклеточные, па которых проводился этот эксперимент, чувствительны к теплоте (являющейся для них жизненно важным биотическим условием, необходимым для обмена веществ), но нечувствительны к свету. Поэтому, если поместить их в равномерно нагретую камеру, часть которой освещена, в то время как другая часть затемнена, они равномерно распределяются но всей камере. Если, наоборот, одну сторону равномерно освещенной камеры нагреть, они сосредоточиваются в нагретом конце камеры. Однако, если в течение длительного периода освещать нагретый конец камеры и затемнять ненагретый, положение дела меняется, и одноклеточные становятся чувствительными к свету, который сейчас приобретает для них значение сигнала к повышению температуры, и начинают сосредоточиваться в освещенном конце камеры, несмотря на разницы температур.
Характерно, что такая чувствительность к освещению формируется у одноклеточных постепенно и при длительном неподкреплении света теплотой может совсем исчезнуть.
Подобную же индивидуальную изменчивость поведения простейших можно вызвать на основе их оборонительных реакций, если изменять условия, вызывающие эти реакции. Примером может служить опыт известного польского исследователя Я. Дембовского.
Одноклеточные помещены в круглую пробирку с водой и обнаруживают характерные для них беспорядочные движения. Если поместить их в такую же трубку, но с четырехугольным сечением, они начинают ударяться о стенки этой пробирки, но скоро их движения приобретают измененный характер, обеспечивающий минимальные удары о стенки сосуда. Траектория этих движений, следовательно, начинает отражать конфигурацию сосуда. Выработанная у них траектория сохраняется даже тогда, когда они снова помещаются в пробирку круглой формы и некоторое время продолжают совершать те же движения по ромбической траектории.
Процесс изменчивости индивидуального поведения простейших, резко отличающий их от растений, происходит относительно медленно, и возникшие изменения так же медленно исчезают. Однако эти изменения настолько значительны, что возникшие новые формы поведения (приспособления к изменившимся условиям) позволяют осуществить нужные реакции приспособления к новым условиям на низшей ступени эволюционной лестницы.
Типичным примером этого может служить эксперимент, проведенный американским исследователем Смитом.
В узенькую пробирку с микроскопическим сечением помещалась туфелька (вид одноклеточных). Сечение трубки было так мало, что для того, чтобы выйти из трубки в направлении действия биотического агента (света), туфельке нужно было перевернуться, ударяясь о стенки трубки. В начале эксперимента на этот поворот у туфельки уходило 3–5 минут, однако если такие эксперименты повторялись много раз в течение 10–12 часов, поворот начинал выполняться много быстрее, и под конец на него требовалось всего 1–2 секунды. Таким образом, под влиянием новых условий вырабатывался новый «навык», который протекал в 180–200 раз быстрее, чем первоначальная реакция.
Как видим, формирование нового вида поведения, отвечающего измененным условиям, требует у простейших животных значительного времени. Характерно и то, что раз возникшее изменение поведения сохраняется у них достаточно долго, и нужно длительное время, чтобы оно исчезло.
Это можно наблюдать как у простейших одноклеточных, так и у относительно просто организованных многоклеточных.
Пример, показывающий такое медленное возникновение и столь же медленное исчезновение новой формы поведения, можно видеть в эксперименте, проведенном сначала бельгийским исследователем Блессом, а затем советским исследователем А. Н. Леонтьевым над плоским червем планарией (см.: Леонтьев А. Н. Проблемы развития психики).
Мы еще не знаем биологических механизмов такого появления чувствительности к ранее нейтральному агенту. Возможно, что оно связано с постепенным изменением биохимических свойств плазмы. Однако факт прижизненного появления новых форм указывает на возникновение индивидуальной изменчивости в поведении простейших и дает основание говорить о возникновении на этой стадии эволюции подлинного, хотя и очень элементарного, поведения.
Механизмы поведения простейших
Науке еще очень мало известно о физико — химических условиях поведения простейших и о тех причинах, которые вызывают положительные или отрицательные движения (движение по направлению к одним объектам и в направлении от других объектов соответственно).
Известно, что протоплазма, составляющая тело простейшего (одноклеточного) животного, состоит из внешнего, более плотного слоя (плазма — гель), и внутреннего, более жидкого слоя (плазма — золь). Известно также, что внешние слои протоплазмы одноклеточного более возбудимы, чем внутренние, и каждое внешнее действие вызывает усиленный обмен веществ, который постепенно распространяется от внешних слоев к внутренним, угасая по типу постепенно снижающегося градиента возбуждения. Эти градиенты возбуждения, по — видимому, и лежат в основе движений одноклеточного, возникающих, как только внешнее воздействие вызовет усиленный обмен веществ в соответствующей точке его поверхности или когда внутренние процессы приведут к необходимости активно искать вещества, необходимые для обеспечения такого обмена.
Некоторые авторы полагают, что адекватные воздействия умеренной силы вызывают положительную реакцию одноклеточного и приводят к движению по направлению к соответствующему воздействию, в то время как неадекватные (сверхсильные или вредные) воздействия вызывают отрицательное движение, направленное от воздействующего объекта. Положительное движение приводит к тому, что вызывающий раздражение объект сначала обволакивается струйками протоплазмы, которая у наиболее простейших одноклеточных (амеб) выпускается в место наиболее интенсивного обмена и замыкается вокруг этого объекта, включая его в состав тела одноклеточного. Если этот объект питателен, он усваивается телом одноклеточного, а продукт распада выделяется в среду. Если он не питателен, он таким же путем, как был захвачен, выбрасывается в окружающую среду.
Было бы, однако, неверно представлять, что все движения одноклеточных протекают по такой простой схеме. Особенностью поведения таких организмов является тот факт, что воздействия, доходящие до них, сами могут претерпевать сложнейшие изменения. Сама протоплазма одноклеточного никогда не находится в состоянии покоя, но характеризуется высоко дифференцированными, избирательными системами возбуждений, которые меняются в зависимости от протекающего в ней процесса обмена веществ и приводят к возникновению доминирующих форм возбуждения.
Как это было показано опытами Фоглера, механическое воздействие известной силы вызывает лишь относительно слабые реакции простейшего, а световое возбуждение может вообще не вызывать никаких реакций. Однако, если на простейшее воздействуют одновременно механическое раздражение данной силы и световое раздражение, оба эти воздействия суммируются и приводят к повышенным реакциям простейшего.
Известно, что реактивность голодной амебы более высока, чем реактивность сытой амебы, и у нее создается повышенная готовность реагировать на соответствующие полезные воздействия или на раздражения, которые сигнализируют об их появлении.
Наконец, одноклеточное может «привыкать» к соответствующим воздействиям, снижая реакции на них по мере их длительного повторного предъявления.
Механизмы, лежащие в основе этого явления, еще мало изучены, и лишь проведенные в последнее время наблюдения заставляют думать, что появление и сохранение этих воздействий является результатом известных модификаций и рибонуклеиновой кислоты, составляющей один из основных компонентов их плазмы.
Следует отметить, что тело простейших является относительно однородным и процесс наиболее интенсивного обмена может возникать у него в месте непосредственно возникающего возбуждения, образуя тем самым временный «головной» конец его тела.
Наиболее сложные одноклеточные имеют несравненно более сложную структуру. У них можно различить постоянные «органы» в виде чувствительных участков протоплазмы, которые образуют, например, «жгутики» у бактерий. Эти «жгутики» находятся в постоянном движении и несут наиболее существенные функции ориентировки во внешней среде; в них и возникают градиенты постоянного возбуждения, которые, распространяясь но остальному телу одноклеточного, приводят его в движение.
Богатство поведения одноклеточных очень велико и подлежит еще специальному изучению, так же, как и те механизмы, которые лежат в его основе. Однако то, что мы уже знаем о них, заставляет думать, что здесь заложены основы той сложной деятельности активной ориентировки в среде, которые в дальнейшем составят важнейшую черту психической деятельности.
Происхождение нервной системы и ее простейшие формы
Описанные процессы раздражимости по отношению к биотическим воздействиям, чувствительности по отношению к нейтральным воздействиям, сигнализирующим о появлении жизненно важных воздействий, и элементарного сохранения следов достаточны для поддержания жизни одноклеточных животных.
Однако они становятся недостаточными с переходом к многоклеточным.
Переход к многоклеточным существенно усложняет условия существования. Питание путем прямой диффузии питательных веществ, занимавшее ведущее место на уровне простейших, здесь заменяется питанием дискретной (концентрированной) пищей; повышается роль активной ориентировки во внешней среде. Становится необходимым обеспечить гораздо более дифференцированные движения и гораздо более быструю проводимость возбуждения, чем та, которая была доступна путем постепенного распространения градиентов возбуждения по протоплазме одноклеточных. Это и приводит к значительному усложнению строения тела многоклеточного, к выделению клеток специализированной рецепции раздражений, доходящих до тела животного, и появлению первых сократительных клеток, несущих ту функцию, которую на дальнейших этапах эволюции возьмут на себя мышечные клетки. Это приводит, наконец, к тому, что в местах прежних градиентов возбуждения начинают откладываться дорожки наиболее возбудимой протоплазмы и образуют наиболее элементарную форму нервной системы, которая у этих животных носит характер диффузной, сетевидной нервной системы.
Все это с особенной отчетливостью можно видеть в строении тела относительно простых многоклеточных, относящихся к классу кишечнополостных, например в строении тела медузы, актинии или морской звезды.
Характерным для этой стадии развития многоклеточных является то, что проводимость возбуждения ускоряется благодаря появлению сетевидной нервной системы во много раз. Если проведение возбуждения по протоплазме не превышает скорости 1–2 микрон в секунду, то с появлением простейшей (сетевидной) нервной системы скорость проведения равна 0,5 метра в секунду (заметим, что при дальнейшем развитии нервной системы и переходе на следующие этапы ее усложнения скорость проведения возбуждения еще более возрастает, доходя у лягушки до 25 метров в секунду, а у высших позвоночных до 125 метров в секунду).
Однако значительные преимущества, которые возникают с явлением первичной диффузной (сетевидной) нервной системы, имеют и свои границы. Как и на описанных выше этапах эволюции, у животных с сетевидной нервной системой еще нет постоянного головного конца, управляющего их поведением. Возбуждение равномерно распространяется по сетевидной нервной системе на все тело животного, а место приложения внешнего раздражения становится временным ведущим пунктом. Только у наиболее сложных кишечнополостных одна часть тела (например, один луч у морской звезды), по своему строению не отличающаяся от других частей тела (лучей), может стать доминирующей, «ведущей» и берет на себя функцию наиболее активного органа при движении. Если у морской звезды отрезать (ампутировать) этот «ведущий» луч, то роль «ведущего» переходит к другому, расположенному рядом с ним лучу.
Естественно, что такое отсутствие постоянного ведущего органа, который мог бы не только воспринимать, но и перерабатывать, кодировать полученную информацию и создавать программы дифференцированного поведения, существенно ограничивает возможности поведения на этом уровне. Эти недостатки устраняются на дальнейших этапах эволюции, особенно с переходом к земному существованию и формированием более сложно построенной ганглионарной нервной системы.
Ганглионарная нервная система и появление простейших программ поведения
Переход к наземному существованию связан со значительным усложнением условий жизни. Прямая диффузия питательных веществ из окружающей среды становится невозможной, наличие готовой плотной (дискретной) пищи — несравненно меньшим. Эта пища находится теперь в резко неоднородной среде, и ориентировка, необходимая для получения пищи, значительно усложняется.
Все это создает необходимость дальнейшего усложнения организма животных, и прежде всего дальнейшую эволюцию сложных органов рецепции и движений и формирования сложных и централизованных аппаратов переработки информации и регулирования движений. Именно этому и соответствует следующий этап в эволюции нервной системы, приводящий к возникновению цепочечной, или ганглионарной, нервной системы, которая впервые появляется у червей и приобретает максимальную сложность у высших беспозвоночных, и прежде всего у насекомых.
Как появление ганглионарной нервной системы, так и формирование поведения, которое осуществляется с ее помощью, знаменуют важнейший скачок в эволюции жизнедеятельности.
Уже у наиболее простых беспозвоночных (червей) можно проследить совсем новый принцип организации нервной системы по сравнению с предыдущей стадией. На переднем головном конце червя сосредоточиваются волокна, которые кончаются химическими и тактильными рецепторами, расположенными особенно густо. Эти аппараты воспринимают химические, термические, световые изменения, происходящие во внешней среде, а также изменения влажности. Сигналы этих изменений проводятся по волокнам и доходят до переднего узла или ганглия, где они сосредоточиваются в нервном «центре», впервые появляющемся на этой ступени эволюции. Здесь эти сигналы перерабатываются (кодируются), и возникающие «программы» поведения в виде двигательных импульсов распространяются по цепочке нервных ганглиев, каждый из которых соответствует отдельному сегменту тела червя. Доходящие до этих ганглиев импульсы вызывают соответствующие движения, направление которых программируется и регулируется передним ганглием.
Здесь возникает новый принцип — централизованность нервной системы, резко отличающейся от принципа построения диффузной (сетевидной) нервной системы. Головной конец червя, где сосредоточена особенно густая сеть хемо-, механо-, термо-, фото — и гигрорецепторов, приобретает ведущую роль, в то время как сегментарные ганглии сохраняют лишь относительную автономию. Это легко проследить, если разрезать тело червя на две половинки. В этом случае передняя половина будет закапываться в землю, сохранив свои организованные движения, в то время как задняя половина будет лишь беспорядочно извиваться, не проявляя никаких признаков организованного движения.
Усложнение строения нервной системы на стадии червей позволяет проследить у них более совершенные (хотя еще очень примитивные) виды формирования новых, индивидуально приобретенных видов поведения. Это было показано в свое время известным американским психологом Р. Йерксом. Он помещал дождевых червей в Т — образную трубку, простейший лабиринт. В левом конце этой трубки червь получал электрический удар, вызывавший оборонительную реакцию. При многократном повторении этого эксперимента у дождевого червя можно было выработать «навык» избегать электрического удара и двигаться направо. Насколько медленно шел этот процесс, видно из следующего: понадобилось свыше 150 проб, чтобы поведение червя приобретало организованный характер и в подавляющем числе проб он начинал двигаться направо, избегая электрического шока (табл. 1.1).
Таблица 1.1 — Формирование новых видов поведения у дождевых червей
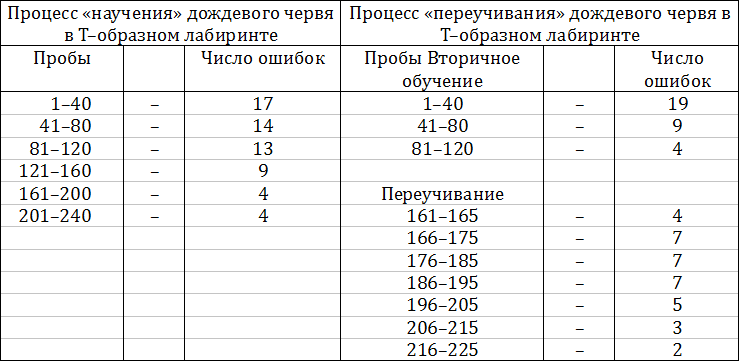
Если повторить этот же опыт после длительной паузы, «обучение» начинает протекать вдвое быстрее, и число ошибок достигает минимального количества уже после 80 опытов. Характерно также, что эксперименты с «переучиванием» червя (в этих опытах червь начинал получать электрический шок уже не слева, а справа) протекали значительно медленнее, и некоторый эффект «переучивания» начинал обнаруживаться лишь после 200 проб.
Отсюда видно, что ганглионарная нервная система червя позволяет не только вырабатывать новые формы поведения, но и сохранять выработанные «навыки», иначе говоря, что дождевой червь обладает примитивной формой «памяти».
В последнее время были проведены эксперименты, которые позволяют убедиться в возможности передачи такого «навыка» и сделать некоторые шаги к выяснению биохимических механизмов, лежащих в основе элементарной памяти. В этих экспериментах американский исследователь Мак Коннэл «обучал» группу червей нужному поведению в простейшем лабиринте. После этого он измельчал тела этих червей, делал из них вытяжку и скармливал ее другим, никогда не обучавшимся червям. Как показал эксперимент, черви, усвоившие эту вытяжку, вместе с этим «усвоили» и навыки, приобретенные первой группой червей. Когда их впервые помещали в лабиринт, они сразу же делали значительно меньше ошибок, чем обычные необученные черви. Этот факт заставил Мак Коннэла предположить, что выработка «навыка» на этих этапах эволюции связана с глубокими биохимическими изменениями протоплазмы и выработанный «навык» может «передаваться» гуморальным путем.
Наука не располагает окончательной оценкой и интерпретацией данных, полученных в этом эксперименте, можно лишь полагать, что изменения, вызванные подобным «обучением», связаны с модификацией рибонуклеиновой кислоты (РНК). Это было показано экспериментами, в которых тела «обученных» червей предварительно опускались в раствор рибонуклеазы (фермент, растворяющий РНК). После этого вытяжка из задних половин тела «обученных» червей не вызывала нужного эффекта, в то время как вытяжка из передних половин «обученных» червей (включавшая вещество переднего ганглия) продолжала вызывать подобный же эффект. Эти эксперименты говорят как о значении для сохранения «навыка» рибонуклеиновой кислоты, так и о том значении, которое для хранения «памяти» червя имеет передний ганглий, клетки которого защищены от растворяющего действия рибонуклеазы.
В науке до сих пор продолжаются споры о том, говорят ли эти эксперименты о «передаче» информации гуморальным путем или лишь об общем «повышении возбудимости», возникающем при усвоении вещества тел обученных червей. Несмотря на то что окончательного ответа на этот вопрос нет, результаты экспериментов показывают — на этом этапе эволюции возможна выработка прочного «навыка», и в сохранении этого «навыка», по — видимому, принимают участие все клетки тела животного.
Появление сложных форм наследственного программирования («инстинктивного») поведения
Дальнейшая эволюция поведения связана с появлением сложных дифференцированных аппаратов рецепции, позволяющих воспринимать высокоспециализированную информацию, приходящую от внешней среды. Она связана и с развитием сложнейших программ, которые позволяют животному приспособляться к сложным, хотя и постоянным, устойчивым условиям среды. Все это становится возможным на дальнейших этапах эволюции ганглионарной нервной системы и особенно отчетливо проявляется у членистоногих животных.
Усложненные условия существования делают необходимым формирование многообразных аппаратов чувствительности, позволяющих регистрировать разнообразные влияния внешней среды. Рассмотрим это на примере эволюции фоторецепторов. Сначала светочувствительные клетки были просто сосредоточены на передней поверхности тела. Это давало животному возможность воспринимать воздействие света, но еще не позволяло локализовать источник света в пространстве. На следующем этапе эволюции светочувствительные клетки сосредоточивались в двух светочувствительных пластинках, расположенных по обеим сторонам переднего конца тела. Это позволяло ориентироваться в пространственном положении источника света и поворачивать туловище в правую или левую сторону, но еще не давало возможности различать свойства действующего на организм предмета. Лишь на последнем этапе эволюции сверхчувствительные пластинки изгибались, принимая форму полого шара. Небольшое отверстие, которое затем заполнялось преломляющей средой (хрусталиком), позволяло падающему лучу преломляться, и воздействие светового объекта запечатлевалось на чувствительном слое этого полого шара. Возникал аппарат сложного светочувствительного рецептора — глаза, который впервые позволил не только реагировать на наличие света, но и отражать свойства воздействующего предмета.
Строение глаза — важнейшего световоспринимащего органа — у различных животных неодинаково. У насекомых оно носит характер «фасеточного глаза», построенного иногда из многих тысяч самостоятельных ячеек. У позвоночных принимает форму хорошо известного нам единого глаза, позволяющего воспринимать отражение предмета и менять четкость отражения с помощью саморегулирующейся системы преломляющего аппарата и мышц. Однако во всех случаях возникновение сложного аппарата, позволяющего на расстоянии ориентироваться в воздействующих предметах, остается одним из наиболее значительных достижений эволюции.
Насекомые располагают большим числом высокодифференцированных рецепторов. Наряду со сложным фоторецептором (глазом) они имеют:
• специальные тактильно — химические рецепторы (расположенные в усиках);
• вкусовые рецепторы (расположенные в полости рта, на ножках), которые улавливают тончайшие изменения вкуса;
• вибрационные рецепторы (расположенные в перепонках ножек), реагирующие на тончайшие ультразвуковые колебания, иногда до 600 тыс. вибраций в секунду.
Возможно, существует еще целый ряд неизвестных нам видов рецепторных аппаратов, специализация которых вырабатывалась у них в процессе миллионов поколений.
Возбуждения, вызванные воздействиями, падающими на эти рецепторные аппараты, распространяются по нервным волокнам и приходят в передний ганглий, который является прототипом головного мозга и аппаратом, объединяющим (кодирующим) доходящие до него импульсы и переводящим эти импульсы в сложнейшие системы врожденных программ поведения, которые лежат в основе приспособительных движений насекомого.
Передний ганглий высших насекомых, например пчелы, имеет очень сложное строение. Он состоит из скопления дифференцированных нервных клеток, к которым приходят импульсы от периферических рецепторов. В передней части этого ганглия распределены преимущественно зрительные клетки, в средней части — обонятельные клетки, в задней — чувствительные клетки ротовой полости. Характерно, что расположение этих клеток имеет организованный характер. В них уже можно наблюдать плоскостное «экранное» строение, позволяющее вызванным возбуждениям распространяться по нейронным структурам переднего ганглия в организованном порядке, обеспечивая тем самым отражение известных структурно организованных воздействий.
Характерно, как это было установлено последними исследованиями, что уже на этой ступени эволюции в состав переднего ганглия входят высокоспециализированные нейроны, которые реагируют на отдельные мельчайшие признаки доходящей до организма информации, разлагая ее на большое число составляющих элементов и позволяя в дальнейшем объединять их в целые структуры (о формах работы этих нейронов будет сказано ниже).
Все это делает передний ганглий высших насекомых сложнейшим центральным аппаратом, позволяющим улавливать многообразные воздействия среды и кодировать их в целые системы.
Коды возбуждений, возникающие при определенных раздражениях в переднем ганглии насекомых, передаются в виде сложных программ поведения на лежащий ниже грудной ганглий, где возникают импульсы сложных приспособительных движений насекомого, составляющие его поведение.
Сложнейшие программы поведения насекомых представляют не только большой интерес, но и требуют специального подробного рассмотрения.
Особенность сложнейших программ, которые составляют подавляющую часть поведения насекомых, состоит в том, что они являются врожденными и передаются по наследству, принимают широко известную форму инстинктивного поведения. Эти программы вырабатываются многими миллионами поколений и передаются наследственно, так же как особенности строения тела (формы крыльев, особенности хоботка, структура рецепторных органов) хорошо приспособлены к условиям существования насекомых.
Примеры врожденных программ поведения у насекомых очень многочисленны. Нередко они настолько сложны и целесообразны, что некоторые авторы считали их примером разумного поведения.
Известно, что личинка березового слоника разрезает березовый лист по идеальной геометрической форме, которая близка к оптимальной, математически рассчитанной структуре, чтобы затем свернуть его в трубочку и использовать для окукливания. Комар откладывает яички на поверхность воды и никогда не откладывает их на сушу, где они неизбежно высохнут. Оса сфекс откладывает яички в тело гусеницы, с тем чтобы появившиеся личинки не испытывали недостатка в пище. Для этого она предварительно прокалывает грудной ганглий гусеницы так, чтобы гусеница не погибла, а лишь была обездвижена, и делает это с удивительной точностью. Нужно ли говорить о врожденных программах поведения паука, ткущего удивительную по своей конструкции паутину, или о врожденных программах поведения пчелы, которая строит соты идеальной, с точки зрения экономии, формы, заполняет эти соты медом и запечатывает их воском, как только они оказываются достаточно наполненными.
Приведенные выше примеры сложнейшего целесообразного поведения и многие другие являются врожденными, им насекомое не должно учиться, оно рождается с этими формами поведения, так же как рождается с идеальной формой крыла или с удивительным по своей целесообразности строением органов чувств.
Все это давало основание многим авторам говорить о целесообразности инстинктов и сближать их с разумным поведением.
Лишь за последнее время исследования зоологов и, в частности, того направления науки, которое называется этологией (этос — поведение), внесли некоторую ясность в загадочную форму поведения и показали, что за этой формой деятельности, поражающей своей сложностью и видимой разумностью, скрыты элементарные механизмы. Эти исследования показали, что сложнейшие программы «инстинктивного» поведения на самом деле вызываются элементарными стимулами, которые пускают в ход врожденные циклы приспособительных актов.
Так, откладывание яичек комара на водной поверхности вызывается блеском воды; поэтому достаточно заменить воду блестящим зеркалом, чтобы комар начинал откладывать яички на его поверхности. Сложная врожденная программа деятельности паука, который бросается на муху, запутавшуюся в паутине, на самом деле вызывается вибрацией паутины, и если к паутине прикасается вибрирующий камертон, паук бросается на него так же, как он бросается на муху.
Описанные механизмы позволяют сделать существенный шаг в улучшении понимания процессов, лежащих в основе врожденного поведения, и перейти от простого описания к его объяснению, показать, насколько инстинктивное поведение отличается от разумного.
Приведем лишь один пример, показывающий, как сложно протекает такое исследование и к каким интересным результатам приводит.
Известно, что некоторые разновидности земляных червей, запасающих на зиму листья, втягивают их в свои норки за конец. Это считалось проявлением «рассудочной деятельности» червей, о которой в свое время говорил Ч. Дарвин, и заставляло предполагать, что червь воспринимает форму листа и «рассчитывает», каким концом лучше втянуть его в норку.
Это предположение существенно изменилось после того, как немецкий исследователь Ганели произвел следующий эксперимент. Он вырезал из листа кусочек, воспроизводящий форму этого листа, но обращенный острием книзу. В этом случае червь делал попытки втянуть лист в норку уже не острым, а тупым концом. Вопрос о том, почему он так делает, стал предметом исследования другого ученого — Мангольда. Этот исследователь предположил, что такое поведение червя диктуется не восприятием формы, а гораздо более элементарным химическим чувством. Для проверки этого он положил перед червем ряд одинаковых палочек, но смазывал один конец этих палочек вытяжкой из верхушек листа, а другой — вытяжкой из основания листа, или один конец вытяжкой из верхушки листа, а другой конец — вытяжкой из черенка. В качестве контрольных были проведены эксперименты, в которых один конец палочки смазывался вытяжкой из верхушки листа или черенка, а другой конец — нейтральным желатином. Результаты опыта показали, что в этих случаях частота, с которой червь втягивал палочку в норку за тот или другой конец, была неодинаковой, и что основным фактором, решающим дело, было различие в химическом отличии вершины листа от его черенка (табл. 1.2).
Таблица 1.2 — Результаты экспериментов Мангольда

Так, если один конец палочки был смазан вытяжкой из верха, другой — вытяжкой из основания листа, червь не оказывал какого — либо предпочтения тому или иному концу палочки. Если один конец палочки смазывался вытяжкой из верхушки листа, а другой — вытяжкой из черенка, червь вдвое чаще втягивал палочку концом, смазанным вытяжкой из верхушки листа. В контрольных экспериментах, где второй конец палочки смазывался нейтральным желатином, это предпочтение вытяжки из верхушки листа становилось еще отчетливее.
Таким образом, гипотеза, что червь реагирует на форму листа, была отброшена и показано, что решающую роль в протекании этой сложной формы деятельности играет гораздо более химическое чувство.
Все описанные наблюдения позволили убедиться в том, что, несмотря на очень сложные программы врожденного поведения, доминирующие у беспозвоночных (и особенно у насекомых), они могут запускаться в ход относительно простыми сигналами. Эти сигналы, запускающие сложнейшие врожденные механизмы, экологические (средовые) условия существования животного, и являются продуктами длительной эволюции.
Условия, которые пускают в ход врожденные программы поведения, особенно отчетливо выступили при анализе того, на какие признаки реагирует насекомое например пчела), когда она избирательно садится на те или иные виды медоносных цветов.
Оказалось, что такими признаками могут являться сложная форма цветка, а иногда его окраска.
Как было показано в исследовании известного немецкого зоолога К. Фриша, а затем экспериментами М. Герц, в которых пчела тренировалась в том, чтобы садиться на чашечки с сахарным раствором, прикрытые картинками с изображением различных геометрических форм, насекомое с трудом различает простые геометрические формы, такие как треугольник и квадрат, но легко различает такие сложные формы, как пятиугольная и шестиугольная звезда или крестообразные формы, расположенные под различным углом. Наконец, сколь легко пчела различала простой и изрезанный круг, не садясь на первый и избирательно реагируя на второй.
Эти исследования показывают, что фактором, позволяющим пчеле выделять соответствующие формы, является не их геометрическая простота, а их сходство с натуральными раздражителями — формой цветов.
Аналогичные результаты дали эксперименты с выделением пчелой различных окрасок. Они показали, что пчела с трудом различает чистые цвета, с гораздо большей легкостью различает смешанные цвета (красно — желтый, желто — зеленый, зелено — голубой и т. д.), которые воспроизводят окраску реальных цветов.
Все это показывает, что решающим фактором для выделения тех признаков, которые пускают в ход врожденные (инстинктивные) программы поведения, являются естественные условия существования (экологические условия), определяющие, какой именно признак выделяется животным.
Очень возможно, что эта высокая избирательность признаков, на которые реагирует насекомое, связана с появлением высокоспециализированных нейронов переднего ганглия, которые в процессе эволюции выработали способность избирательно реагировать на такие жизненно важные признаки, как смешанные цвета, звездчатость или изрезанность формы, расположение формы в пространстве и т. д.
Исследования, проведенные зоологами и психологами, позволили убедиться еще в одной важнейшей особенности врожденного «инстинктивного» поведения. Оказалось, что врожденные программы «инстинктивного» поведения являются целесообразными лишь в определенных, строго постоянных стандартных условиях, в соответствии с которыми в процессе эволюции сложились эти программы. Стоит, однако, немного изменить эти условия, чтобы врожденные программы переставали быть целесообразными и теряли свой «разумный» характер.
Это положение можно иллюстрировать двумя примерами. Известно, что у одной из пород ос сложилось очень «целесообразное» поведение. Подлетая с добычей к норе, в которую она помещает добычу, она оставляет эту добычу около норы, затем влезает в нору и после того, как она находит эту нору пустой, выходит наружу, втаскивает добычу в нору и улетает.
Дело, однако, существенно меняется, когда в специальном эксперименте добычу, лежащую перед входом, сдвигают на несколько сантиметров, проделывая это в тот период, когда оса производит свою разведку в норе. В этом случае оса выходит из норы, не находит добычу на прежнем месте, снова подтаскивает ее в исходное положение и… снова входит в нору, которую она только что обследовала. Поведение осы может повторяться много раз подряд, и каждый раз, когда добыча перемещается на несколько сантиметров, оса продолжает механически повторять обследование норы, потерявшее при этих условиях свою целесообразность.
Аналогичные наблюдения были проведены над пчелами. Известно, что пчела заполняет пустые соты медом и, отложив нужное количество меда, запечатывает соты. Однако, если в условиях социального опыта отрезается весь низ сот, и мед, который пчела кладет в соты, проваливается, пчела продолжает запечатывать пустые соты через определенный период времени, закрепленный в ее инстинктивной программе. И здесь поведение, которое было целесообразным в постоянных стандартных условиях жизни, теряет свою целесообразность, когда условия жизни меняются.
Все это показывает, что врожденные «инстинктивные» программы поведения, преобладающие в деятельности насекомых, являются косными, механическими, сохраняя свою кажущуюся «разумность» лишь в постоянных стандартных условиях, в соответствии с которыми они были выработаны в процессе эволюции.
Целесообразность такого преобладания видовых программ поведения соответствует основному биологическому принципу жизни насекомых. Как правило, они откладывают огромное число яичек, отличающееся большой избыточностью. Лишь небольшое число особей, которое появляется из этих яичек, выживает, однако это число особей является достаточным для сохранения вида. Поэтому, несмотря на то что большое число особей погибает, когда изменения условий делают врожденные программы поведения неадекватными, вид все же сохраняется, и врожденные программы «инстинктивного» поведения оказываются достаточными для сохранения вида.
Такой тип приспособления врожденных программ поведения с медленной и трудной изменчивостью является биологически целесообразным в условиях огромной избыточности воспроизведения, которая имеет место в мире насекомых. Однако он становится биологически недостаточным для другой ветви эволюции — позвоночных, у которых такая избыточность воспроизведения не имеет места, и создаются условия, делающие необходимым появление нового типа — индивидуально изменчивого поведения на более высоком уровне.
Центральная нервная система и индивидуально изменчивое поведение позвоночных
Все, что мы знаем о способах жизни и поведении позвоночных, показывает, что как формы жизни, так и формы поведения их построены по совершенно иному принципу.
Только у низших позвоночных, живущих в водной среде, избыточность воспроизведения столь велика, что приближает их к классу насекомых, лишь сравнительно небольшое число особей, возникающих из оплодотворенной икры, выживает. Поэтому только у рыб преобладание мало изменчивых наследственно программированных форм поведения достаточно для сохранения вида.
Иное мы имеем с переходом позвоночных к наземному существованию. Условия питания становятся сложнее, среда — изменчивее, и требования к ориентировке в этой постоянно меняющейся среде неизмеримо возрастают. Одновременно изменяется и тип воспроизведения: каждая особь воспроизводит только 2–3, реже 5–6 себе подобных, и выживание отдельных индивидов становится условием для сохранения вида.
Все это создает биологическую необходимость для появления, наряду с врожденным и мало изменчивым «инстинктивным» (или видовым) поведением, нового — индивидуально изменчивого поведения. Эта новая форма поведения существует в зачатке у низших позвоночных, но на дальнейших ступенях эволюции начинает занимать все большее и большее место, чтобы у высших млекопитающих (обезьян), а затем у человека окончательно оттеснить низшие, врожденные («инстинктивные») формы поведения.
Необходимость усложнения форм ориентировки в окружающей действительности и выработки новых возможностей индивидуальной ориентировки в меняющихся условиях среды и формирование новых индивидуально изменчивых форм приспособления приводят в процессе эволюции к созданию принципиально новых нервных аппаратов, которые могли бы не только получать сигналы из среды и пускать в ход врожденные, наследственно передаваемые программы «инстинктивного» поведения, но которые могли бы анализировать поступающую информацию и замыкать новые связи, обеспечивая новые индивидуально изменчивые нормы поведения. Таким аппаратом является головной мозг, строение которого отражает длительный путь, пройденный эволюцией.
Головной мозг животного построен по типу ряда надстраиваемых друг над другом уровней или этажей.
Низшие уровни, расположенные в стволе мозга, обеспечивают процессы:
• регуляции внутреннего состояния организма — обмена веществ (гипофиз);
• дыхания и кровообращения (ствол);
• рефлекторных ответов на элементарные раздражители, доходящие до животного из внешней среды (четверохолмие).
Эти аппараты, построенные по типу нервно — секреторных агрегатов или нервных узлов (ганглиев), преобладают у низших позвоночных и позволяют им осуществлять более элементарные врожденные программы приспособительной деятельности, которые немногим отличаются от видов инстинктивной деятельности, описанных выше.
Типичным примером такого строения мозга является мозг лягушки, который состоит из образований верхнего ствола с развитым четверохолмием и лишь намеченными более высокими структурами и обеспечивает прежде всего выполнение сложившихся в процессе эволюции «инстинктивных» и мало изменчивых программ поведения. Эти программы поведения имеют строение, близкое к описанному выше. Лягушка, замечая мушку, делает прыжок, раскрывая челюсти, и захватывает ее липким языком. Это сложное поведение определяется относительно элементарным стимулом — зрительно воспринимаемым мельканием и вовсе не является ответом на предварительно проанализированный сигнал. Это легко видеть, если подвесить на тонком волосе небольшую бумажку и привести ее в движение вращением этого волоса. В этом случае мелькание бумажки пускает в ход врожденные программы поведения, и лягушка автоматически бросается на мелькающую бумажку, как она раньше бросалась на мушку.
Над аппаратом ствола надстраиваются более высокие образования, включающие сначала таламо — стриальную систему (подкорковые узлы) и древнюю (обонятельную, или лимбическую) кору, а затем, у высших позвоночных — образования новой коры больших полушарий, которые все больше и больше развиваются и постепенно начинают полностью доминировать над образованиями более низкого уровня.
Аппараты таламо — стриальной системы, которые начинают преобладать у пресмыкающихся и у птиц, а затем и аппараты древней коры обеспечивают более пластичные формы индивидуального поведения, чем те формы, которые имелись у низших позвоночных. Однако эти формы поведения, приобретающие уже черты индивидуальной изменчивости, еще тесно связаны с более элементарными врожденными формами поведения и носят черты тех видов приспособительной деятельности, с которыми животное рождается на свет и которые отражают особенности его экологии (т. е. устойчивых биологических форм его существования).
Ближайший анализ показывает, что эти иногда очень сложные врожденные программы поведения могут вызываться относительно простыми стимулами, отражающими формы жизни животного. Как показали наблюдения ряда авторов, в том числе советского физиолога А. Д. Слонима, сложные сосательные движения только что родившегося животного фактически вызываются разными сигналами, отражающими биологические условия его существования: у щенка сосательные движения вызываются мягкой шерстью (основным признаком кормящей матки), у ягненка — затемнением темени (отражающим биологический факт, что ягненок, начинающий сосать, прежде всего подходит под овцу и сосет закидывая голову). Такая же экологическая обусловленность избирательных реакций характерна для многих животных и больше зависит от способов их существования, чем от того зоологического класса, к которому они относятся.
Так, домашняя утка (питающаяся растительной пищей) безразлична к гнилостным запахам, но тонко реагирует на растительные, в то время как кобчик, питающийся падалью, тонко реагирует на гнилостные запахи и остается безразличным к растительным.
Характерно, что экологические особенности отражаются и на более сложных формах поведения животного. Хорошо известно, что при виде пищи у собаки начинает выделяться слюна. Однако, что менее известно, у лисы, которая предварительно должна добыть пищу, вид ее приводит к торможению, задержке слюны.
Все эти примеры показывают, что основные формы натурального поведения, осуществляемого аппаратами более высоких разделов головного мозга, сохраняют теснейшую связь с врожденными программами поведения, которые формируются под влиянием условий существования (экологии) животных.
На только что описанные аппараты надстраивается аппарат следующего, наиболее высокого уровня нервной системы больших полушарий, в частности новой коры головного мозга, вес которой, как показывают приведенные ниже данные, начинает занимать все большее место по отношению к весу тела (табл. 1.3).
Таблица 1.3 — Вес мозга по отношению к весу тела
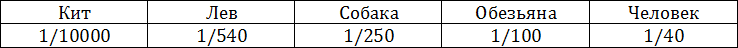
Большие полушария мозга не только начинают приобретать в процессе эволюции высших позвоночных большой относительный вес, но и получают важное значение. Если у низших млекопитающих кора еще не имеет ведущей роли и регуляция процессов поведения может успешно осуществляться даже при ее разрушении, у высших млекопитающих она приобретает более ведущее значение, сколько — нибудь сложные процессы поведения уже не могут осуществляться при ее разрушении.
Факты, указывающие на большую зависимость функций от коры мозга, иначе говоря, на прогрессивную кортикализсщию функций, можно видеть из результатов наблюдений немецкого исследователя Шафера, показывающих, какие процессы восприятия и движения могут оставаться на разных этапах эволюции млекопитающих после удаления коры. Мы приводим данные этих наблюдений в сводной таблице (табл. 1.4).
Таблица 1.4 — Влияние экстирпации коры головного мозга на зрительные и двигательные процессы на последовательных ступенях эволюции (по Шаферу)

Все это показывает, что большие полушария не только начинают занимать большое место по отношению к остальному мозгу, но играют ведущую роль в регуляции сложных форм поведения животного.
Главная функция больших полушарий и их основной составной части — коры головного мозга — заключается в том, что она (кора) является аппаратом, который не только воспринимает сигналы и пускает в ход заложенные видовые программы поведения, но и позволяет анализировать информацию, поступающую из внешней среды, ориентироваться в ее изменениях, замыкать новые связи и формировать новые индивидуально изменчивые программы поведения, соответствующие этим изменениям.
Этой новой задаче отвечает строение коры больших полушарий головного мозга, толща которой состоит из огромного числа изолированных нервных клеток (нейронов), расположенных по шести основным слоям. Причем четвертый слой служит местом окончания волокон, несущих раздражение от органов чувств и воспринимающих поверхностей тела, и их переключения на другие нейроны. Пятый слой является местом, откуда двигательные импульсы направляются к мышцам, и, наконец, последняя группа слоев (второй и третий) сосредоточивает огромное число нейронов, перерабатывающих (перекодирующих) импульсы, которые поступают в кору головного мозга и замыкают новые связи, обеспечивающие формирование новых, индивидуально изменчивых программ поведения (детальное описание строения коры больших полушарий будет дано в гл. IV).
Характерная особенность коры больших полушарий состоит в том, что ее разные области заняты проекцией различных органов чувств (обоняния, зрения, слуха, тактильной, мышечно — суставной чувствительности). Экологический принцип построения работы центрального нервного аппарата сказывается и здесь, и чем большее значение в жизни животного имеет тот или иной вид чувствительности, тем важнее его место в коре головного мозга.
Так, в коре головного мозга ежа (животное ориентируется преимущественно обонянием) обонятельные отделы мозга занимают почти треть полушарий, в то время как у человека обонятельные отделы мозга резко редуцированы.
У обезьяны ведущее место имеет ориентировка во внешнем мире, поэтому зрительные отделы коры занимают большое место, в то время как у собаки (преимущественно обонятельного животного), гораздо меньшее место занимают области больших полушарий.
То же самое можно сказать об относительном месте, которое занимают в коре головного мозга проекции органов тела. Это было показано экспериментами английского физиолога Э. Эдриана, который с помощью специальной электрофизиологической методики установил, что территория, которую занимает проекция бедра в мозгу свиньи, очень невелика, в то время как проекция «пятачка» свиньи (который является для нес наиболее важным органом ощупывания, обнюхивания и т. д.) занимает огромную, непропорциональную его размерам территорию. Характерно, что такое же непропорционально большое место занимает в больших полушариях овцы проекция губ — основной орган ориентировки. Все это показывает, что кора больших полушарий головного мозга, мощно развивающаяся на позднейших этапах эволюции высших позвоночных, формируется под прямым воздействием экологических условий, и именно это делает ее основным аппаратом, обеспечивающим сложнейшие формы анализа и синтеза информации, поступающей из среды, и основным органом регуляции индивидуального изменчивого поведения.
Механизмы индивидуально изменчивого поведения
Как показал американский исследователь Дэшналл, пробы, которые делает животное, помещенное в лабиринт, не случайны, они, как правило, всегда идут в общем направлении к цели, поэтому животное, которое хоть раз ориентировалось в лабиринте, создает общую систему направления, в котором оно бежит, и много раз чаще заходит в тупики лабиринта, расположенные по направлению данного пути, чем в тупики лабиринта, расположенные в направлении, обратном схеме движения.
Таким образом, можно наглядно видеть, что движения животного в лабиринте не носят случайный характер, а подчиняются одному направлению, которое другой американский исследователь, Креч, охарактеризовал как возникновение «двигательной гипотезы», руководящей общим направлением попыток, которые делают крысы, вырабатывающие «навык» в лабиринте. Подтверждением того, что попытки, которые делает животное при выработке новых программ поведения, не являются случайными, а есть результат активной ориентировки животного в условиях среды, являются экспериментами с так называемым «латентным обучением».
В этих экспериментах сравнивается быстрота выработки навыка у животных, которые сразу же помещаются в лабиринт с целью его прохождения, и у второй группы животных, которым разрешается просто бегать по лабиринту и, следовательно, дается возможность предварительно сориентироваться в условиях лабиринта.
Как показали данные эксперименты, проведенные американскими исследователями, вторая группа животных вырабатывала нужный навык значительно быстрее, чем первая. Характерно, что выработка успешного навыка была особенно велика, когда животному разрешалось активно ориентироваться в условиях лабиринта. Подобный эксперимент был поставлен американским исследователем Хелдом, который позволял животному активно двигаться по проблемному ящику, в то время как другое животное пассивно наблюдало этот ящик из коляски, которую возило первое животное. И в этом случае первое животное, активно ориентировавшееся в обстановке, значительно быстрее вырабатывало двигательный навык, чем второе. Этот факт показывает ту роль, которую играет в выработке новой программы действий предварительная ориентировка животного в условиях опыта.
Сложный динамический характер выработанного двигательного навыка, не являющегося простой цепью механически усвоенных, подтверждается, наконец, специальной серией экспериментов, которые показывают, что такой вновь выработанный «навык» может сохраняться даже и в тех случаях, когда для его выполнения нужен совсем новый набор движений.
Одно из таких исследований принадлежит американскому исследователю Хантеру. В этом эксперименте у крысы вырабатывался навык нахождения нужного пути в лабиринте. После того как такой навык был выработан, одна часть лабиринта выключалась, и на ее место ставился бассейн с водой. В этом случае крыса должна была переплывать бассейн, т. е. производить совершенно новые движения, но от этого выработанный навык не разрушался.
Аналогичные факты были получены американским исследователем Лешли другим путем. Выработав у крысы навык нахождения пути в лабиринте, он затем разрушал ее мозжечок. В результате этой операции у крысы, потерявшей равновесие, все прежние движения нарушались, передвигаясь кубарем (т. е. с помощью совершенно новых движений), она все же перемещалась в нужном направлении и достигала цели.
Все это показывает, что двигательные программы, вырабатываемые в условиях специальных задач, ни в какой мере не являются механической цепью рефлексов, и они скорее являются сложными динамическими схемами, формирующимися у животного в процессе его ориентировочной деятельности.
Две особенности характерны для выработки таких новых форм приспособительной деятельности у позвоночных.
Первая из них состоит в том, что процесс ориентировки поиска неотделим от активных движений и нахождение нужного решения не предшествует здесь выполнению двигательной программы, а формируется в процессе двигательных проб.
Вторая особенность заключается в том, что как характер ориентировочных действий животного, так и характер выработанных новых форм поведения в высшей степени зависит от натуральных форм поведения, сформированных в тесной связи с особенностью жизни (экологией) животного. У травоядных, и прежде всего у животных, питающихся готовой пищей (например, курица, овца, корова) ориентировочная деятельность носит пассивный, ограниченный характер, натуральные программы поведения преобладают, и выработка новых, индивидуально изменчивых форм деятельности протекает медленно. Наоборот, у хищников, которые в естественных условиях вынуждены разыскивать пищу, охотиться за жертвой (хищные птицы, лиса), ориентировочная деятельность протекает в виде активных поисков, новые формы поведения, которые соответствуют меняющейся обстановке, вырабатываются гораздо быстрее.
Существенно, что и характер выработанных форм нового, индивидуально изменчивого поведения сохраняет у этих животных связь с их врожденными («инстинктивными») формами деятельности, или «натуральными рефлексами». Так, морского льва, у которого ныряние, преследование жертвы входит во врожденные программы поведения, особенно легко обучить жонглированию мячом или ловле мяча. Енота или медведя, которые часто становятся на задние лапы, оставляя передние свободными, можно обучить движениям, имитирующим «стирку белья», и т. п.
Эта теснейшая связь врожденных программ поведения (или «натуральных рефлексов») с выработкой новых, индивидуально изменчивых форм деятельности остается характерной для поведения высших позвоночных на данном этапе развития.
«Интеллектуальное» поведение животных
Описанные формы возникновения индивидуально изменчивого поведения не являются, однако, высшей границей эволюции поведения в животном мире.
У позвоночных, стоящих на вершине эволюционной лестницы, в частности у приматов, возникают новые формы индивидуально изменчивого поведения, которые с полным основанием могут быть обозначены как «интеллектуальное» поведение.
Особенность «интеллектуального» поведения животных заключается в том, что процесс ориентировки в условиях задачи не протекает в условиях двигательных проб, а начинает предшествовать им, выделяясь в особую форму предварительной ориентировочной деятельности, в процессе которой начинает вырабатываться схема (программа) дальнейшего решения задачи, в то время как движения становятся лишь исполнительным звеном в этой сложно построенной деятельности. Таким образом, на высших этапах эволюции начинают формироваться особенно сложные виды поведения, имеющего сложную расчлененную структуру, в которую входят:
• ориентировочно — исследовательская деятельность, приводящая к формированию схемы решения задачи;
• формирование пластически изменчивых программ движений, направленных на достижение цели;
• сличение выполненных действий с исходным намерением.
Характерным для такого строения сложной деятельности является ее саморегулирующийся характер:
• если действие приводит к нужному эффекту, оно прекращается;
• если оно не приводит к нужному эффекту, в мозг животного поступают сигналы о «рассогласованности» результатов действий с исходным намерением, и попытки решить задачу начинаются снова.
Такой механизм «акцептора действия» (П. К. Анохин), т. е. динамического контроля действия, является важнейшим составным звеном всякого индивидуально изменчивого поведения животного, но проявляется с отчетливостью в наиболее сложной фазе эволюции поведения — интеллектуальном поведении.
Два существенных явления, зачатки которых можно видеть уже на наиболее ранних ступенях эволюции позвоночных, предшествуют формированию этой наиболее высокой формы поведения животных. Первым из них является возникновение особой формы ориентировочной деятельности, названной советским исследователем Л. В. Крушинским «экстраполяционным рефлексом»; вторым является факт более усложнявшихся форм развития памяти у животных.
В наблюдениях, проведенных Л. В. Крушинским, установлено, что некоторые животные проявляют в своем поведении способность подчиняться не непосредственному восприятию предмета, но прослеживать его движения и ориентироваться на ожидаемое перемещение объекта. Известно, что собака, перебегающая улицу, не бежит прямо под движущуюся автомашину, а делает петлю, учитывая движение машины и даже развиваемую ею скорость. Этот рефлекс, «экстраполирующий» наблюдаемое движение и учитывающий перемещение, Л. В. Крушинский проследил в ряде экспериментов.
В этих экспериментах животное помещалось перед трубой, которая в середине имела разрыв. На глазах у животного к проволоке, проходящей через трубу, прикреплялась приманка, которая двигалась по трубе; она появлялась перед глазами животного в разрыве трубы и двигалась дальше, пока не появлялась в конце трубы. Животное помещалось перед разрывом трубы и наблюдало движение приманки.
Данные наблюдения показали, что животные, стоящие на более низкой ступени эволюции, и, в частности, животные, которым свойственно лишь собирать готовую пищу (например, курица), непосредственно реагировали на место, где появлялась приманка, и не отходили от него. В противовес этому животные, стоящие на более высокой ступени эволюции, и, в частности, животные, ведущие хищный образ жизни, прослеживающие добычу и преследующие ее (ворон, собака), следили за движением приманки и, «экстраполируя» ее движение (очевидно, направляя свое поведение движением глаз), обегали трубу и ожидали приманку в месте ее появления.
«Экстраполяционный рефлекс», который имеет особую форму — «предвосхищающего» поведения, является одним из важных источников для формирования наиболее высоких «интеллектуальных» видов индивидуально изменчивого поведения высших позвоночных.
Выше было отмечено, что вторым фактом, создающим существенные условия для формирования «интеллектуального» поведения высших позвоночных, является возрастающая сложность процесса восприятия и большая прочность памяти на последовательных ступенях эволюции животных.
Известно, что если низшие позвоночные реагируют лишь на определенные признаки воздействий, идущих, из внешней среды, то высшие позвоночные больше реагируют на целые комплексы знаков или на образы окружающих предметов. Эта реакция животных была детально изучена советским физиологом академиком И. С. Беритовым и составляет важнейшее условие для эволюции сложных форм поведения.
Одновременно с формированием образного восприятия на высших этапах эволюции позвоночных наблюдается возрастающая прочность образной памяти. Этот факт был детально прослежен в экспериментах с так называемыми «отсроченными реакциями» животных.
Эксперименты с отсроченными реакциями проводились многими американскими исследователями, советским психологом Н. Ю. Войтонисом и польским физиологом Ю. Конорским. Суть эксперимента заключалась в следующем. Животное помещалось перед герметически закрытым ящиком, в который на глазах животного клалась приманка.
Животное, привязанное к стойке, задерживалось па привязи в течение известного времени, после чего отпускалось. Если в памяти животного сохранялся след приманки, положенной в ящик, оно сразу же бежало к этому ящику, если этот след исчезал, животное к ящику не подбегало.
В более сложных экспериментах, которые ставили задачей проверить четкость сохранившегося у животного следа, положенная в ящик приманка незаметно подменялась другой. Если след первой приманки у животного сохранялся, то подбегая к ящику и находя другую приманку, — оно ее брало. Это было признаком того, что у животного сохранился избирательный образ той приманки, которую оно видело.
В других экспериментах животное помещалось между двумя ящиками, в один из которых на глазах у животного помещалась приманка. После истечения некоторого времени животное спускалось с привязи. Если след от приманки, положенный в один из ящиков, сохранялся, то животное бежало к этому ящику, если след не сохранялся, направленного движения у животного не было.
Эксперименты с отсроченными реакциями показали, что на последовательных ступенях эволюционного развития позвоночных длительность сохранения соответствующих образов возрастает (табл. 1.5).
Таблица 1.5 — Длительность сохранения следов однократно вызванной образной памяти у различных животных

Естественно, что длительное сохранение образов памяти увеличивается по мере усложнения мозговых структур и создает второе важное условие для возникновения высших «интеллектуальных» форм поведения животного.
Систематические исследования «интеллектуального» поведения высших животных (обезьян) были начаты в 20–х гг. прошлого века известным немецким психологом В. Кёлером. Для изучения этой формы поведения В. Кёлер ставил обезьян (шимпанзе) в условия, где непосредственное достижение цели было недоступно, и обезьяна должна была ориентироваться в сложных условиях, в которых дана цель, и либо использовать обходной путь, чтобы получить приманку, либо обратиться для этой цели к использованию специальных орудий.
Опишем три типичных ситуации, в которых В. Кёлер проводил свои исследования «интеллектуального» поведения обезьяны.
Первая ситуация требовала «обходного пути». Обезьяна помещалась в большую клетку, рядом с которой была положена приманка, находившаяся на таком расстоянии, что рука обезьяны не могла ее достать. Для достижения цели обезьяна должна была прекратить попытки непосредственно достигнуть цели и использовать обходной путь через дверь, расположенную в задней стене клетки.
Вторая ситуация была близка к только что описанной, т. е. обезьяна помещалась в закрытую клетку, которая на этот раз имела двери. Приманка располагалась также в отдалении, и обезьяна не могла достать ее рукой. Однако в отличие от первой ситуации перед клеткой на расстоянии вытянутой руки лежала палка. Обезьяна могла достать приманку, дотянувшись до палки, и при ее помощи достигнуть цели. В усложненных экспериментах приманка была расположена еще дальше, но в поле зрения обезьяны лежали палки: короткая — на расстоянии руки, и длинная — несколько дальше. Решение задачи заключалось в том, что обезьяна должна была осуществить более сложную программу поведения. Сначала дотянуться до ближайшей — короткой палки, затем с ее помощью достать длинную палку, расположенную дальше от нее, и уже с помощью этой палки достать приманку.
Наконец, в третьем варианте экспериментов приманка подвешивалась так, что обезьяна непосредственно не могла ее достать. Однако на этой же площадке были разбросаны ящики; обезьяна должна была подтащить ящики к приманке, поставить их один на другой и, взобравшись на эти ящики, достать приманку.
Исследования, проведенные В. Кёлером, позволили ему наблюдать следующую картину.
Сначала обезьяна безуспешно пыталась непосредственно достать приманку, тянулась к ней или прыгала. Эти безуспешные попытки могли продолжаться длительное время, пока обезьяна не истощалась и не бросала их.
Затем наступал второй период, который заключался в том, что обезьяна неподвижно сидела и лишь рассматривала ситуацию; ориентировка в ситуации переносилась здесь из развернутых двигательных проб в «зрительное поле» восприятия и осуществлялась с помощью соответствующих движений глаз.
После этого наступал решительный момент, который В. Кёлер описывал как неожиданное появление «переживания». Обезьяна либо сразу же направлялась к дверце, расположенной в задней стенке клетки и «обходным путем» доставала приманку, либо переставала непосредственно тянуться к приманке, подтягивала к себе палку и с ее помощью доставала, либо подтягивала одну палку, доставала ею вторую, более длинную и уже этой палкой доставала приманку; наконец, в последней ситуации обезьяна прекращала всякие попытки непосредственно достать приманку, оглядывалась вокруг, а затем сразу подтаскивала ящики, ставила их один на другой и, взобравшись на них, доставала приманку.
Характерным для всех этих экспериментов был тот факт, что решение задачи перемещалось из периода непосредственных проб в период предшествующего попытке наблюдения, и движения обезьяны становились лишь исполнительным актом для осуществления заранее выработанного «плана решения».
Именно это и дало основания В. Кёлеру рассматривать поведение обезьяны как пример «интеллектуального» поведения.
Если описание поведения обезьян в экспериментах В. Кёлера является исчерпывающим, то объяснение тех путей, которыми животное приходит к «интеллектуальному» решению задачи, представляет большие сложности, и этот процесс трактуется разными исследователями неодинаково.
Известный американский психолог Р. Йеркс, повторивший исследование В. Кёлера, считает возможным сблизить эти формы поведения обезьяны с человеческим интеллектом и антропоморфически рассматривает их как проявления «творческого озарения».
Австрийский психолог К. Бюлер привлекает для объяснения этого поведения прежний опыт животного и считает, что использование орудий обезьянами следует рассматривать как результат переноса прежнего опыта (обезьянам, живущим на деревьях, приходилось притягивать к себе плоды за ветки).
Сам В. Кёлер высказывает предположение, что в «интеллектуальном» поведении обезьян анализ ситуации перемещается из сферы движений в план восприятия, и обезьяна, рассматривая ситуацию, «сближает» входящие в нее предметы в «зрительном поле», замыкая их в известные «зрительные структуры». Последующее решение задачи есть, по мнению В. Кёлера, лишь осуществление «зрительных структур в реальных движениях». Подтверждение этой гипотезы В. Кёлер видит в том факте, что в случаях, когда палка и приманка (плод) или две палки, которые обезьяна должна последовательно достать, расположены так, что они не попадают в одно зрительное поле, задача становится неразрешимой для обезьяны.
Свою гипотезу В. Кёлер пытается подтвердить экспериментами, в которых обезьяна должна раньше приготовить орудие, которое она в дальнейшем использует, чтобы достать приманку, обезьяна должна вставить одну бамбуковую палку в другую, с тем чтобы, удлинив ее, достать плод. Эти действия оказываются для обезьяны гораздо труднее и могут быть выполнены только в случае, если концы обеих палок попадают в наглядное поле; такое совмещение обеих палок в одном зрительном поле, по мнению В. Кёлера, и может привести к нужному решению задачи.
Вопрос о механизмах, лежащих в основе возникновения «интеллектуального» поведения обезьяны, нельзя считать окончательно решенным, и если одни исследователи противопоставляют его более элементарным формам индивидуально изменчивого поведения животных, то другие (как например, И. П. Павлов, проводивший наблюдения над поведением обезьян) считают возможным не противопоставлять его более простым формам поведения и рассматривают «интеллектуальное» поведение обезьян как своего рода «ручное мышление», выполняемое в процессе проб и ошибок и приобретающее более богатый характер лишь в силу того, что руки обезьян, освободившиеся от функции ходьбы, начинают осуществлять наиболее сложные формы ориентировочной деятельности.
Границы индивидуально изменчивого поведения животных
Мы отметили, что индивидуально изменчивое приспособительное поведение высших позвоночных может достигать очень сложных форм. Возникает естественный вопрос: каковы его характерные черты и те границы, за пределы которых поведение животных может выйти?
Анализ поведения животных позволяет наметить основные черты.
Первую из них составляет тот факт, что всякое, даже наиболее сложное, индивидуально изменчивое поведение животного сохраняет свою связь с биологическими мотивами и не может перейти за их границы. В основе всякого поведения животного лежат биологические влечения или потребности (потребность в пище, самосохранении или половая потребность). Лишь на наиболее высших этапах эволюции к этому присоединяется потребность ориентировки в окружающей среде, которую И. П. Павлов называл ориентировочным рефлексом и которая достигает значительного развития у обезьян. Никакая деятельность, не связанная ни с одной из этих биологических потребностей, невозможна для животного, поэтому поведение животного с полным основанием может рассматриваться как корково — подкорковое.
Вторая особенность, которая характеризует поведение всякого животного, заключается в том, что оно всегда определяется непосредственно воспринимаемыми стимулами («внешним полем») или следами прежнего опыта и не может протекать, отвлекаясь от них или тем более вступая в конфликт с ними.
Это положение хорошо иллюстрируется известным исследованием голландского психолога Ф. Бойтендайка, имеющим принципиальное значение.
Ф. Бойтендайк помещал перед животным ряд ящиков, в которые можно было класть приманку. В первом эксперименте приманка на глазах у животного клалась в первый ящик, и животному разрешалось брать ее. Во втором эксперименте приманка (также на глазах животного) перемещалась во второй; затем в третий ящик. Затем, в последующих экспериментах, приманка (уже незаметно для животного) начинала последовательно перемещаться в каждый следующий ящик, и животному каждый раз разрешалось свободно бежать к тому ящику, в котором оно предполагало найти приманку.
Исследование показало, что животное всегда бежит либо к тому ящику, куда на его глазах была положена приманка, либо к тому, где она лежала раньше. Никакое животное не может усвоить отвлеченный принцип «последовательного передвижения» и никогда не бежит с следующему ящику, в котором приманки еще не было, но в котором легко ожидать ее, если учесть абстрактный принцип.
Поведение животного всегда направляется непосредственным или прошлым опытом, и оно никогда не может затормозить реакцию на ранее подкрепляемое и направиться к ранее не подкрепленному месту. Опыт Ф. Бойтендайка, проведенный на животных разных групп, показывает, что животное, по выражению В. Кёлера, является «рабом своего зрительного поля», или «рабом своего прошлого опыта», и что его поведение никогда не освобождается от непосредственных влияний, не направляется абстракцией от них, иначе говоря, не становится свободным.
Третья особенность поведения животного заключается в ограниченности источников этого поведения.
Источником поведения животного могут быть программы, либо заложенные в его видовом опыте и передающиеся в наследственных кодах («инстинктивное» поведение), либо формируемые в непосредственном опыте данной особи (индивидуально изменчивое или условно — рефлекторное поведение). Никаких возможностей усвоения чужого опыта и передачи его, усвоенного одним индивидом, другому индивиду и тем более передачи опыта, сформированного в нескольких поколениях, у животных не имеется. Те явления, которые описываются как «подражание», у животных занимают в формировании их поведения относительно ограниченное место и являются скорее формой непосредственной практической передачи собственного опыта, чем передачей информации, которая накопилась в истории ряда поколений и сколько — нибудь напоминала бы то усвоение материального или духовного опыта прошлых поколений, которое характерно для общественной истории человека.
Эти три черты и составляют основные особенности поведения животного и коренным образом отличают его от сознательной деятельности человека.
Назад: ГЛАВА 1. Психология как наука. Ее предмет и практическое значение
Дальше: Глава 3. Сознательная деятельность человека и ее общественно — исторические корни

