Квантовое измерение
Несмотря на свою объяснительную силу, квантовая механика не говорит нам ничего о том, как сделать шаг от уравнений и описания того, как электрон, скажем, движется вокруг атома, к тому, что мы видим при конкретном измерении этого электрона. По этой причине отцы-основатели квантовой механики предложили набор временных правил, которые стали приложением к математическому формализму. Они известны как квантовые постулаты и предоставляют своего рода руководство о том, как перевести данные математического моделирования уравнений в осязаемые объекты, которые мы можем наблюдать, такие как положение или энергия атома в любой заданный момент.
Что касается реального процесса, когда атом перестает находиться «здесь или там» и находится только «здесь», то никто не знает, что происходит. Большинство физиков с радостью приняли прагматичную точку зрения о том, что это «просто происходит». Проблема в том, что ситуация требует определения различий между квантовым миром, где происходят такие чудеса, и нашим ежедневным макромиром, где поведение объектов основано на органах чувств. Измерительный прибор, который обнаруживает электрон, должен быть частью этого макромира. Но как, где и когда этот измерительный процесс имеет место, основатели квантовой механики никогда не уточняли.
Вопрос хрупкости квантовой когерентности (удержание волновой функции от исчезновения), несомненно, является главной задачей группы МТИ, с которой мы познакомились в начале этой главы, и их коллег по всему миру в их стремлении создать квантовый компьютер. Вот почему они так скептически отнеслись к заявлению в New York Times о том, что растения представляют собой квантовые компьютеры. Физики применили все виды интеллектуальных и дорогостоящих уловок, чтобы защитить квантовый мир внутри своих компьютеров от разрушающей когерентность внешней среды. Таким образом, идея, что квантовая когерентность может поддерживаться в жарком влажном и молекулярно турбулентном климате внутри травинки, была, очевидно, воспринята как безумие.
Однако теперь мы знаем, что на молекулярном уровне многие важные биологические процессы могут на самом деле происходить очень быстро (порядка триллионов в секунду) и также могут быть ограничены короткими атомными расстояниями. Именно такие расстояния и скорости, на которых происходят квантовые процессы, например туннелирование, могут имеет эффект. Таким образом, несмотря на то, что полностью избежать декогерентности невозможно, ее можно отсрочить настолько, чтобы получить биологический эффект.
Путешествие к центру фотосинтеза
Взгляните на секунду на небо, и в ваш глаз попадет поток света длиной около 300 тысяч километров. В ту же секунду на земле растения и фотосинтезирующие микроорганизмы используют поток солнечного света для получения около 16 тысяч тонн нового органического вещества в виде деревьев, травы, водорослей, ландышей, гигантских секвой и яблок. В этом разделе мы должны разобраться, как действительно происходит этот первый шаг в трансформации неживого вещества в практически всю биомассу нашей планеты; и в качестве примера трансформации мы возьмем превращение воздуха Новой Англии в яблоко на дереве Ньютона.
Чтобы увидеть этот процесс в действии, мы снова позаимствуем наноподлодку, которую использовали для исследования действия ферментов в предыдущей главе. Когда вы взойдете на борт, держите курс наверх, в листву дерева, на один из его растущих листьев. Лист продолжает расти, пока его дальние края не скроются за горизонтом, а его на первый взгляд гладкая поверхность превращается в неровную платформу, вымощенную зелеными кирпичиками с вкраплениями небольших круглых светлых блоков, в каждом из которых расположена дырка-пора. Зеленые кирпичи называются клетками эпидермиса, а круглые блоки – устьицами: их функция – пропускать воздух и воду (субстраты фотосинтеза) с поверхности листа в его внутреннюю среду. Вы ведете аппарат над ближайшим устьицем и, когда длина судна уменьшается до микрометра (миллионной части метра), снижаетесь, чтобы пройти сквозь пору и оказаться внутри зеленой и яркой среды листа.
Оказавшись внутри, вы решаете отдохнуть в уютном и тихом интерьере листа, выстланном рядами зеленых клеток, похожих на камни, и накрытом толстыми цилиндрическими канатами. Канаты – это вены листа, которые либо несут воду от корней к листу (сосуды ксилемы), либо переносят только что синтезированные сахара от листа ко всему растению (сосуды флоэмы). Если вы продолжите уменьшаться в размере, то увидите, что клетка, похожая на камень, расширяется во всех направлениях, пока не увеличится до размеров футбольного поля. В таком масштабе – ваш рост равен примерно десять нанометров, или одна стотысячная миллиметра, – вы можете увидеть, что ее поверхность покрыта дерном из сети волокнистых ячеек, совсем как толстый джутовый ковер. Этот волокнистый материал представляет собой клеточную стенку, своего рода экзоскелет. Ваша наноподлодка оснащена инструментами, которые вы используете, чтобы проложить путь через этот волокнистый ковер и увидеть восковой нижний слой, клеточную мембрану, последний водонепроницаемый барьер между клеткой и внешней средой. При более близком рассмотрении выясняется, что она не полностью гладкая, а пронизана отверстиями, заполненными водой. Эти мембранные каналы называются поринами и представляют собой водопроводную систему клетки, пропускающую внутрь питательные вещества и выводящую продукты распада. Чтобы проникнуть в клетку, вам придется только подождать возле одного из поринов, пока он не расширится настолько, чтобы вы могли проникнуть в водную среду клетки.
Уже через канал порина вы сможете сразу увидеть, что внутренняя среда клетки очень отличается от внешней. Вы не найдете великолепных колонн и широких залов. Интерьер заполнен и в некотором роде беспорядочен. Он выглядит как очень шумный деловой центр! Водянистая жидкость, заполняющая клетку, цитоплазма, плотная и вязкая; местами она больше похожа на гель, чем на жидкость. В геле подвешены тысячи неправильных шаровидных объектов, которые находятся в состоянии непрерывного внутреннего движения. Это белковые ферменты, похожие на те, что мы видели в предыдущей главе, ответственные за проведение метаболических процессов в клетке, разрушают питательные вещества и синтезируют биомолекулы, такие как углеводы, ДНК, белки и жиры. Многие их этих ферментов привязаны к сети кабелей (цитоскелету клетки), которая, совсем как горнолыжный подъемник, переносит многочисленные грузы в различные места клетки. Транспортная сеть исходит из нескольких узлов, где кабели прикрепляются к большим зеленым капсулам. Эти капсулы представляют собой хлоропласты клетки, в которых происходит центральный процесс фотосинтеза.
Вы ведете подлодку через вязкую цитоплазму. Вы продвигаетесь медленно, но в конце концов подходите к ближайшему хлоропласту. Он лежит под вами, как огромный зеленый воздушный шар. Он, как внутренняя клетка, окружен прозрачной мембраной, через которую видны большие стопки зеленых монетоподобных предметов. Это тилакоиды, заполненные молекулами хлорофилла, пигмента, придающего растениям зеленый цвет. Тилакоиды – это механизмы фотосинтеза, которые при заправке фотонами света могут скреплять атомы углерода (полученные из углекислого газа воздуха) вместе с образованием сахаров, которые пойдут в наше яблоко. Чтобы лучше рассмотреть первый этап фотосинтеза, вы направляете свой аппарат в одну из пор мембраны хлоропласта, к верхней зеленой монете стопки тилакоидов. Достигнув своей цели, вы выключаете двигатель подлодки, позволяя ей дрейфовать над этой электростанцией фотосинтетических процессов.
Перед вами лежит только один из триллионов фотосинтетических механизмов, которые производят мировую биомассу. С вашей выгодной точки вы можете увидеть, что, как мы узнали при изучении ферментных механизмов в предыдущей главе, несмотря на множество происходящих вокруг вас турбулентных столкновений молекул, подобно бильярдным шарам, существует определенная упорядоченность. Поверхность мембраны тилакоида усыпана скалистыми зелеными островками, покрытыми древоподобными структурами с похожими на антенны пятиугольными пластинками на концах. Эти пластинки-антенны представляют собой светопоглощающие молекулы, хромофоры, самым известным примером которых является хлорофилл. Именно здесь происходит первый ключевой этап фотосинтеза: захват света. Вероятно, вторая по значимости молекула на нашей планете (после ДНК) хлорофилл заслуживает более подробного рассмотрения (рис. 4.5).
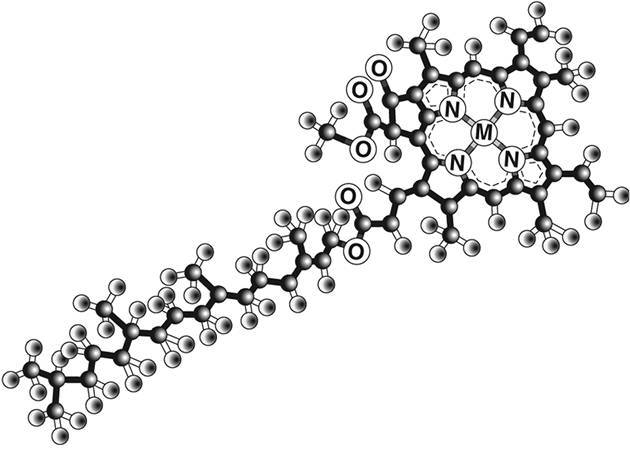
Рис. 4.5. Молекула хлорофилла
Это двухмерная структура, состоящая из пятиугольных элементов, включающих в основном атомы углерода (серые сферы) и азота (N), с атомом магния (М) в центре, с длинным хвостом из атомов углерода, кислорода (О) и водорода (белые атомы). Внешний электрон магния слабо связан с атомом и может выбиваться в окружающий углеродный каркас при поглощении фотона солнечного света. В результате вместо него остается пустое место и атом получает положительный заряд. Это пустое место, или электронную дырку, можно рассматривать с абстрактной точки зрения как «вещь в себе»: положительно заряженную дырку. Суть в том, что мы расцениваем оставшийся атом магния как нейтральный, пока посредством поглощения фотона создаем систему, состоящую из выбитого отрицательного электрона и оставшейся положительной дырки. Эта бинарная система называется экситоном (рис. 4.6). Ее можно считать крошечной батареей с положительным и отрицательным полюсами, способной хранить энергию для последующего использования.
Экситоны нестабильны. Электрон и его дыра ощущают воздействие электростатической силы, притягивающей их друг к другу. При их воссоединении солнечная энергия изначального фотона теряется в виде остаточного тепла. Таким образом, если растению необходимо использовать поглощенную солнечную энергию, оно должно очень быстро перенести экситон в производственную часть молекулы, известную как реакционный центр, где происходит процесс под названием «разделение зарядов». Фактически он включает полный отрыв энергетического электрона от атома и перенос его к соседней молекуле, подобно действию ферментов, которое мы наблюдали в прошлой главе. В результате этого процесса образуется более стабильная, чем экситон, химическая батарея (под названием НАДФН), которая используется во всех важных химических реакциях фотосинтеза.
Но реакционные центры обычно располагаются достаточно далеко с молекулярной точки зрения (на расстоянии нанометров) от возбужденных молекул хлорофилла, так что энергия должна переходить от одной молекулы-антенны к другой по хлорофилльному лесу, пока не достигнет реакционного центра. Это возможно благодаря плотно упакованной структуре хлорофилла. Молекулы по соседству с той, которая захватила фотон, также приходят в возбуждение, эффективно принимая энергию от первичного возбужденного электрона и затем перенося ее к собственному электрону атома магния.
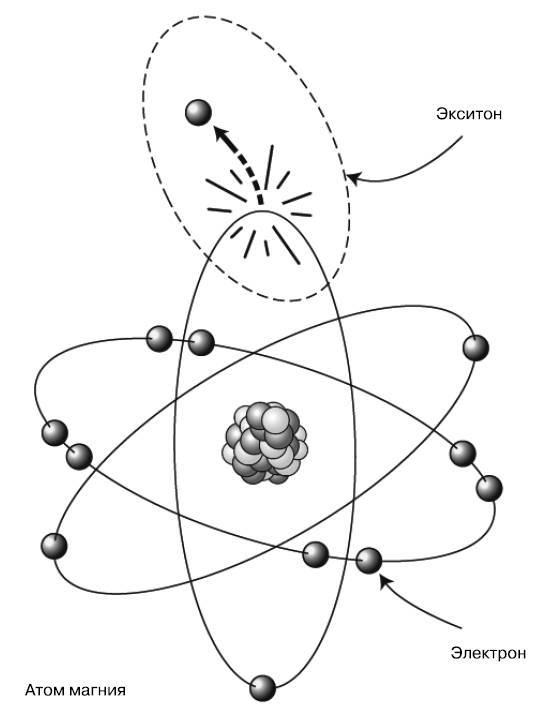
Рис. 4.6. Экситон состоит из электрона, выбитого со своей орбиты, и атома с оставшейся дыркой
Проблема заключается в том, каким путем должна происходить передача энергии. Если она выберет неправильное направление, в случайном порядке перескакивая от одной молекулы к другой в хлорофилльном лесу, энергия будет утрачена и не достигнет реакционного центра. Какой путь она должна выбрать? Путь к цели не должен занимать много времени, чтобы не утратить энергию экситона.
До недавних пор считалось, что перенос энергии от одной молекулы хлорофилла к другой носит случайный характер, в сущности принимая характер стратегии последней надежды, известной как метод случайного блуждания. Иногда это называют «пьяным блужданием», потому что оно напоминает маршрут пьяницы, вышедшего из бара, который блуждает в поисках пути, пока случайно не находит свой дом. Но случайное блуждание – не очень эффективный способ добраться куда-нибудь: если дом пьяного далеко, он может проснуться следующим утром в кустах в другой части города. Объект, участвующий в случайных блужданиях, имеет тенденцию удаляться от точки старта на расстояние, пропорциональное квадратному корню из времени. Если за одну минуту пьяный человек продвинется на один метр, то через четыре минуты он уйдет на два метра, а через девять минут – только на три метра. С таким вялым прогрессом неудивительно, что животные и микробы редко используют случайное блуждание, чтобы найти пищу или добычу, прибегая к этой стратегии только при отсутствии другого выхода. Поместите муравья в незнакомую местность, и, как только он уловит запах, он бросит случайное блуждание и будет следовать за своим носом.
Считалось, что, не имея ни носа, ни навыков навигации, энергия экситона продвигается по хлорофилльному лесу методом пьяницы. Но такая картина не имела особого смысла, так как известно, что первый этап фотосинтеза чрезвычайно эффективен. Фактически перенос энергии захваченного фотона от молекулы-антенны хлорофилла к реакционному центру знаменит свой эффективностью, большей, чем у любой естественной или искусственной реакции: почти 100 %. При оптимальных условиях почти каждая частица энергии, поглощенная молекулой хлорофилла, достигает реакционного центра. Если бы выбранный путь был блуждающим, то почти все они, по крайней мере большинство из них, должны были быть утеряны. Почему эта энергия фотосинтеза находит свой путь к конечной цели намного успешнее, чем пьяница, муравей или наша наиболее энергоэффективная технология? Это остается одной из величайших загадок биологии.

