Книга: Онтогенез. От клетки до человека
Назад: Часть III Последние штрихи
Дальше: Глава 15 Как набраться ума-разума
Глава 14
Рожденные смертными
В расцвете жизни носим мы в себе смерть.Книга общественного богослужения
Один из многих парадоксов человеческой жизни заключается в том, что она зависит от смерти – смерти огромного количества отдельных клеток вполне здорового организма. Я говорю не о случайной гибели от неизбежных повреждений и не о смерти от бактериальных или вирусных инфекций. Речь идет о преднамеренном самоубийстве, когда клетки активируют белки, запускающие контролируемый процесс их разрушения. Было подсчитано, что таким образом погибают – в ходе абсолютно нормального развития – более половины клеток эмбриона. Поскольку клетки сами избирают путь саморазрушения, этот процесс также называют «избирательной гибелью клеток».
Добровольная гибель клеток может быть связана, например, с тем, что образуемая ими ткань нужна только на промежуточных этапах формирования организма, а затем становится лишней. Такие ткани можно сравнить со строительными лесами, необходимыми для возведения, скажем, арочного моста: когда мост готов и может держаться сам по себе, леса убирают. У эмбриона примером могут послужить, вероятно, ткани временных почек (см. главы 10 и 12). У низших животных, например у рыб, эти почки сохраняются и функционируют у взрослых особей, но у взрослых млекопитающих выделение осуществляют почки нового типа (глава 12). Однако первые клетки крови и кровеносные сосуды у эмбрионов млекопитающих образуются, как и у рыб, в комплексе тканей, который включает примитивные почки. В мужском организме часть «дренажной» системы временной почки идет на формирование каналов репродуктивной системы. Поэтому при эмбриональном развитии млекопитающих без временных почек не обойтись, хотя во взрослом организме они и не выполняют свою исходную выделительную функцию.
Избирательная гибель клеток играет важную роль при образовании ладоней и стоп эмбриона. Они закладываются как лопатообразные структуры, в которых кости пальцев, образующиеся за счет уплотнения мезенхимы, одеты, словно рукавицей, общим слоем эктодермы (глава 11). Затем клетки, расположенные между пальцами, погибают, кожа оттягивается к ладони, и кисть становится похожей уже не на рукавицу, а на перчатку. Роль избирательной гибели клеток при образовании пальцев была показана в экспериментах на эмбрионах птиц. У куриных эмбрионов она выражена очень сильно, и на лапах формируются длинные, хорошо разделенные пальцы, которыми удобно копаться в земле. У утиных эмбрионов, напротив, уровень клеточной смертности в промежутках между пальцами довольно низок. В результате формируются прекрасно приспособленные для плавания перепончатые лапы с прочным двойным слоем кожи и соединительной ткани между пальцами. При обработке куриных эмбрионов препаратом, ингибирующим избирательную гибель клеток, получаются цыплята с перепончатыми лапами (рис. 70).
Наряду с уничтожением вспомогательных тканей, избирательная гибель клеток также используется для ликвидации лишних клеток в постоянных тканях. Многие развивающиеся ткани изначально производят избыточное число клеток, а затем, используя сигналы от других тканей, определяют, какие клетки и в каком количестве нужно сохранить.
Ярчайший пример такого перепроизводства клеток – формирование популяции моторных нейронов развивающегося спинного мозга. (Моторные нейроны иннервируют мышцы стенки туловища и конечностей, отвечая за их движение.) Давайте рассмотрим общий принцип этого явления на примере рук. При нормальном развитии в области спинного мозга, управляющей руками, производится гораздо больше моторных нейронов, чем требуется взрослому организму., От этих моторных нейронов в развивающиеся конечности отходят аксоны, которые должны вступить в контакт с волокнами развивающихся мышц. Через некоторое время происходит всплеск избирательной гибели клеток, и численность моторных нейронов значительно снижается.
Первые проблески понимания того, что же управляет избирательной гибелью клеток, пришло в ходе экспериментов,,, в которых один зачаток передней конечности (крыла) куриного эмбриона удаляли, а другой оставляли на месте. Сначала по обе стороны спинного мозга образовывалось, как и следует, очень много моторных нейронов. После всплеска избирательной гибели клеток на стороне спинного мозга, связанной с развивающейся конечностью, погибало нормальное, относительно небольшое количество мотонейронов, а с той стороны, где конечности не было, наблюдалась массовая гибель мотонейронов. Этот классический эксперимент показал, что «решение» моторного нейрона (жить ему или умереть) зависит от того, удалось ли его аксону найти мышцу, к которой можно подключиться. Этот предварительный вывод был в дальнейшем подтвержден экспериментом по имплантации дополнительной конечности с одной стороны тела. Такая имплантация значительно снижала уровень гибели мотонейронов, как если бы им было нужно иннервировать мышцы двух конечностей.
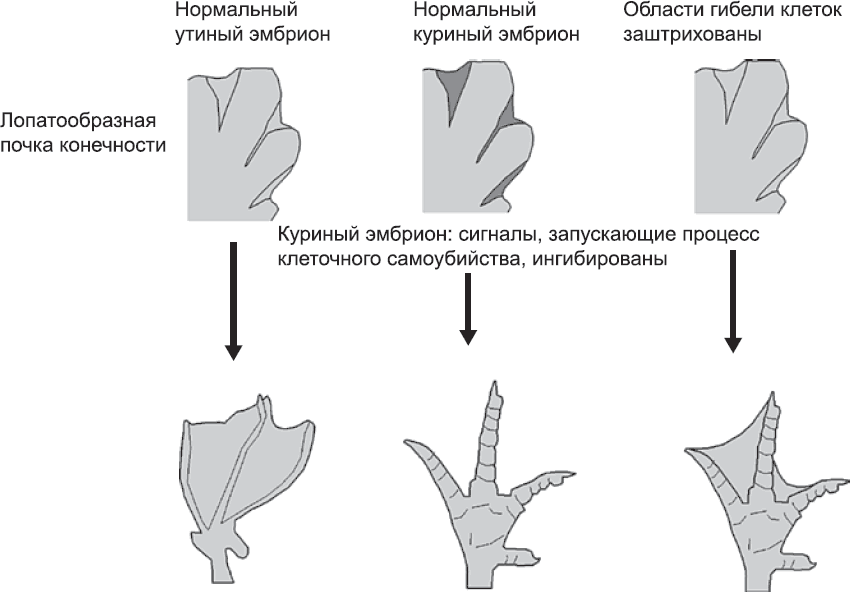
Рис. 70. Роль гибели клеток в исчезновении перепонок между пальцами. Слева показана схема развития нормальной утиной лапы: гибель клеток между пальцами в зачатке конечности незначительна, перепонки сохраняются. На схеме посередине показано развитие нормальной куриной лапы: избирательная гибель клеток между пальцами сильно выражена (как и в случае человеческой руки), и образуются конечности без перепонок. Схема справа показывает результаты эксперимента, в котором сигналы, в норме вызывающие избирательную гибель клеток в конечности цыпленка, были ингибированы. Образовавшаяся конечность похожа на утиную. Это позволяет предполагать, что избирательная гибель клеток имеет принципиальное значение для исчезновения перепонок. Я говорю «предполагать», а не «считать доказанным», так как нельзя полностью исключать, что метод, используемый для ингибирования сигналов, также влияет на какой-то иной, пока неизвестный процесс в конечности. «Неизвестные неизвестности», как метко окрестил это явление Дональд Рамсфелд, – вечная проблема, возникающая при интерпретации биологических экспериментов
Когда ученые поняли, что при изменении количества мышц может изменяться число погибающих мотонейронов, а некоторые мотонейроны гибнут даже при нормальном развитии, они предположили, что мышц всегда немного меньше, чем мотонейронов. Тщательный биохимический анализ развивающихся мышц показал, что они производят факторы выживания нейронов в очень ограниченном количестве. На ранних стадиях развития мотонейроны могут обходиться без этих факторов, но затем впадают в зависимость от них. Самоубийственный потенциал заложен в клетках изначально, и только сигналы от факторов выживания не дают привести его в исполнение. Мышцы рук нормального эмбриона производят недостаточно факторов выживания, чтобы сохранить все мотонейроны, и только клетки, образовавшие наилучшие соединения с мышцами, получают его в нужном количестве. Другие клетки получают слишком слабый сигнал, недостаточный для сдерживания «тяги к самоубийству». Поэтому такие клетки самоуничтожаются. Таким образом, многочисленные клетки конкурируют друг с другом за факторы выживания, и выживают лишь те, которым удалось создать оптимальное соединение с источниками этих факторов.
Перепроизводство клеток с последующим выбором лучших напоминает перепроизводство молодых животных с последующим отбором наиболее приспособленных (основной принцип естественного отбора, лежащий в основе дарвиновской теории эволюции). Параллель неточная, но обе системы действительно предоставляют способ выбрать «наиболее приспособленные» единицы, обладающие случайной изменчивостью. В случае дарвиновской эволюции изменчивость выражается в разных комбинациях генов, которые несут конкурирующие между собой молодые особи. Изменчивость мотонейронов эмбриона в основном выражается в том, насколько точно их аксоны находят путь к цели. В обоих случаях наиболее приспособленные выживают, а остальные погибают. Благодаря такой организации процесса отпадает необходимость в сверхточном программировании пути движения аксонов, а также повышается устойчивость эмбриона к сбоям в работе системы.
Принцип конкуренции клеток за поступающие от цели сигналы выживания не ограничивается мотонейронами спинного мозга. Он также проявляется в сенсорной системе и во многих областях головного мозга. Самое интересное, что этот принцип проявляется не только в нервной системе. Его широкое распространение легло в основу так называемой трофической гипотезы, согласно которой выживание любых клеток зависит от ограниченного количества факторов выживания, производимых другими клетками. Один из авторов этой гипотезы был настолько уверен в ее истинности, что, когда впервые рассказывал о ней на конференции, предложил внушительную премию тому, кто найдет противоречащий ей пример (оговорившись, что аномальные клетки и эмбрионы на самых ранних стадиях развития в расчет не принимаются). Премия так и осталась невостребованной.
Трофический принцип полезен для объяснения процессов развития, происходящих как на очень коротких, так и на очень длительных периодах времени. Во-первых, в соответствии с этим принципом клетки, которые образовались не там, где надо, или же «заблудились» и оказались далеко от будущих партнеров, просто покончат жизнь самоубийством, не причинив никому неудобств. Во-вторых, трофический принцип, возможно, обусловил эволюцию сложных организмов. Рассмотрим двух гипотетических животных с одинаковой формой тела. В организме одного из них сразу образуется именно столько мотонейронов, сколько нужно для обслуживания мышц рук, а в организме другого, как и у нас, сначала образуется слишком много мотонейронов, а затем избыток ликвидируется. Теперь представьте, что окружающая среда этих животных меняется, и открывается новая экологическая ниша, подходящая для животных с сильными конечностями (например, теперь нужно рыть землю или прыжками перемещаться с дерева на дерево). Чтобы животные первого типа получили более развитые, но функционирующие передние конечности, у одной особи должны одновременно произойти две мутации: одна, чтобы увеличить руки, а другая, чтобы увеличить количество мотонейронов ровно настолько, насколько нужно. Для животных второго типа будет достаточно мутации, которая приведет к увеличению рук, потому что количество выживших мотонейронов будет корректироваться автоматически с учетом нового, большего источника факторов выживания. Вероятность возникновения одной мутации мала; вероятность возникновения двух мутаций у одной и той же особи крайне мала, и такого стечения обстоятельств придется ждать очень долго. Животные, в организме которых имеет место перепроизводство клеток и последующее избирательное выживание, находятся в лучших условиях и имеют больше шансов выиграть эволюционную гонку – то есть эволюционировать быстрее и занимать новые экологические ниши. Поэтому неудивительно, что эти принципы лежат в основе развития сложных животных с долгой эволюционной историей, таких как мыши или люди. Не будь их, такие животные, возможно, просто не смогли бы возникнуть за всю эволюционную историю Земли.
Зависимость клеток от сигналов выживания, посылаемых другими клетками, имеет ряд интересных клинических следствий, и уже сейчас вмешательство в эти сигналы помогает спасать жизни людей. Раковые клетки утрачивают все обычные механизмы регулирования размножения (что, по большому счету, и делает их раковыми клетками), но многие из них все же зависят от сигналов выживания. Это ахиллесова пята некоторых опухолей, потому что иногда удается «убедить» их клетки совершить самоубийство, а это гораздо менее вредно для организма, чем традиционная химиотерапия. Такой подход особенно перспективен для борьбы с опухолями не жизненно важных тканей, потому что если в процессе лечения некоторые нормальные клетки тела, относящиеся к тому же типу ткани, тоже решат погибнуть, ничего особенно страшного не произойдет. Например, выживание клеток предстательной железы зависит от сигналов тестостерона. (К слову, наблюдение Карла Фогта (1840 г.), что простата молодых бычков резко уменьшается после кастрации, было первым четким описанием избирательной гибели клеток после нарушения сигнала выживания.) Многие виды рака и предраковых новообразований предстательной железы сохраняют свою зависимость от тестостерона и других гормонов, поэтому препараты, блокирующие передачу сигналов тестостерона, могут быть очень эффективны в борьбе с такими опухолями., Сходным образом, многие виды рака молочной железы зависят от сигналов эстрогена. Мощным ингибитором сигнализации эстрогена в тканях молочной железы является тамоксифен, и этот препарат действительно оказался очень эффективным при лечении рака молочной железы у многих пациенток. Использование этого подхода для борьбы с опухолями жизненно важных тканей осложняется тем, что полное блокирование сигналов выживания может повлечь за собой уничтожение не только опухоли, но и нормальной ткани. Однако, поскольку опухоли нередко расположены беспорядочно, их клетки и так находятся довольно далеко от источников сигналов выживания. Поэтому теоретически возможно подобрать такую дозу препарата, которая поддерживала бы уровень сигнала на достаточном уровне для выживания нормальной ткани на обычном месте, но на слишком низком уровне для выживания клеток опухоли. В сочетании с другими методами управление сигналами выживания может стать ценным подспорьем в лечении разнообразных раковых заболеваний.
Назад: Часть III Последние штрихи
Дальше: Глава 15 Как набраться ума-разума

