Глава 7
Жизнь после триумфа
Границы подхода
В тот же самый период, когда бихевиоризм тихо угасал в своей глухой обороне – примерно с середины 1950-х до начала 1970-х, – этология, несмотря на острую критику извне и изнутри, быстро набирала вес и популярность, желающих заниматься ею становилось все больше. Рассмотрев причины неудачи бихевиористского подхода к поведению, мы должны сказать несколько слов о причинах успеха подхода этологического.
В предыдущих главах мы уже неоднократно убеждались, что всякое исследование поведения неизбежно требует выбора: либо как-то интерпретировать наблюдаемое поведение, пытаясь так или иначе понять, что стоит за ним, либо сразу отказаться от таких попыток и рассматривать поведение как окончательную реальность. Бихевиористы выбрали второе – и их опыт показал, что такой подход в любых вариантах и версиях приводит в тупик. Однако перспективы первого пути выглядели не более обнадеживающими. Ранняя попытка пройти по нему – истолкование поведения животных по аналогии с человеческим поведением – была окончательно дискредитирована опытом классической зоопсихологии, других сколько-нибудь проверяемых методов такой интерпретации не просматривалось. Казалось, вопрос «что стоит за поведением?» допускал только два типа возможных ответов: реконструкцию психики животного со всеми ее образами, эмоциями, намерениями, воспоминаниями, самосознанием и т. д. либо сведе́ние поведения к чисто физиологическим механизмам. Первое применительно к бессловесным существам представлялось абсолютно невозможным. Второе оставалось предметом споров – возможно ли это в принципе или это такая же утопия, как надежда рассчитать будущее мира, измерив координаты и импульсы всех составляющих его тел? Впрочем, эти споры носили сугубо академический характер: даже самые горячие сторонники сводимости поведения к физиологии (такие, как Павлов) прекрасно понимали, что современная им физиология еще невообразимо далека от того уровня развития, который необходим для решения этой задачи.
Успех этологии был обеспечен тем, что ее создатели нашли еще один путь. Морфологический подход позволил им определить структуру поведения. Опираясь на нее, они смогли разработать эвристические схемы тех механизмов, которые должны обеспечивать именно такую структуру. Эти схемы не были привязаны ни к каким конкретным нервным центрам, ядрам и образованиям и потому оказывались свободными от ограничений, налагаемых недостаточным развитием нейрофизиологии. Не были они и в полном смысле слова реконструкцией субъективного мира изучаемого животного – несмотря на то что этологи безусловно признавали существование у животных психической жизни и острое желание посмотреть на мир глазами своего объекта, увидеть его «умвельт» никогда не покидало их. «С каждым животным, которое я изучал, я становился этим животным. Я старался думать, как оно, чувствовать, как оно. Вместо того чтобы смотреть на животное с человеческой точки зрения – и делать серьезные антропоморфические ошибки в процессе, – я пытался как исследователь-этолог поставить себя на место животного так, чтобы его проблемы стали моими проблемами и я не вычитывал бы ничего в его образе жизни, что было бы чуждо этому конкретному виду», – писал один из первых оксфордских учеников Тинбергена, известный этолог Десмонд Моррис. При этом созданные этологами методы – анализ коммуникативного поведения, позволяющий вычленить те движения и позы, которые самими животными воспринимаются как «смысловые», определение ключевых стимулов и разработка на их основе сверхстимулов и т. д. – казалось бы, давали возможность отчасти осуществить эту мечту. Но только отчасти: можно экспериментально выяснить, какими признаками должно обладать яйцо, чтобы вызвать у гусей сильнейшие родительские чувства, – но как ощутить, как пережить сами эти чувства? Иэн и Ория Дуглас-Хэмилтон (ученики Тинбергена, изучавшие поведение слонов в Африке) рассказывают в своей книге, как Тинберген, приехав к ним в гости и став объектом атаки слонихи-вожака, сразу понял, что эта атака – не более чем демонстрация. Проницательность классика произвела сильнейшее впечатление на свидетелей этой сцены, знавших, что он никогда специально не занимался поведением слонов. Но даже и он вряд ли мог представить, что чувствовала, что переживала в этот момент сама слониха.
По сути дела, этологи реконструировали не внутренний субъективный мир изучаемых ими животных, а структуру психических процессов – то, что скрыто от самого субъекта, то, что «человеческие» психологи самых разных школ, от Зигмунда Фрейда до Александра Лурии, пытались выяснить применительно к человеку. Выражаясь фигурально и несколько упрощенно, этологический подход реконструирует не «картинку на мониторе сознания», а те программы, выполнение которых явлено самому субъекту поведения в виде этой картинки. Точнее даже – алгоритмическую схему этих программ (поскольку «внутренний язык программирования» поведения нам пока неизвестен).
Это был, конечно, огромный прорыв – но он относился только к области врожденного видоспецифичного поведения. За ее пределами лежало необозримое пространство поведения индивидуального – прежде всего разнообразных форм и видов научения. Этологи прекрасно сознавали необходимость исследования этих феноменов. Но как? Базовый для этологии морфологический подход к поведению – выделение характерных динамических форм – тут был почти бесполезен.
Некоторые этологи попытались работать с теми феноменами и аспектами научения, до которых можно было дотянуться, не покидая твердой почвы морфологического подхода. Прежде всего это, конечно же, относилось к импринтингу, который безусловно можно рассматривать как специфическую форму обучения. Именно к 1950–1960-м годам относится новый всплеск интереса к этому явлению. Этологи находили все новые и новые примеры импринтинга, порой весьма неожиданные и экзотические. В частности, феномен запечатления был обнаружен у муравьев, только что вышедших из куколки; выяснилось, что проходные лососи запечатлевают вкус и запах реки, в которой они вылупились из икринки, и т. д. Устанавливались критические периоды, критерии выбора объектов для запечатления, отдаленные проявления детского запечатления. Было открыто явление многократного импринтинга (у самок многих млекопитающих – на своих детенышей), сформулированы некоторые закономерности самого процесса. Словом, работы хватало.
Лоренц нашел и другие точки возможного приложения этологического метода к проблеме научения. Он, в частности, подметил высокую стереотипность выученного поведения животных в естественных условиях – и дал ей блестящую теоретическую интерпретацию. Но, пожалуй, самым значительным достижением Лоренца (и всего этологического направления) в этой области стал предложенный им общетеоретический подход к проблеме научения, трактовавший выученное поведение как «надстройку» над врожденной основой или «вставку» в нее. Сходным образом рассматривал такое поведение и Тинберген, но Лоренц проработал идею глубже – до выводов о видоспецифичной предрасположенности к тем или иным формам выученного поведения. Например, неспособность некоторых высокоразвитых животных обучиться выполнять команды человека отражает вовсе не их «тупость» или «низкий уровень умственных способностей» (как об этом писали многие авторы, начиная еще со времен Бюффона), а лишь то, что в природе эти животные ведут одиночный образ жизни и не имеют в своем естественном поведении такой формы, как «повиновение лидеру». Еще нагляднее этот эффект проявляется в том, чему именно может обучиться то или иное животное: зайца сравнительно легко научить барабанить, морского льва – удерживать на кончике носа мяч, енота – полоскать одежду и т. д. То есть легче всего животное учится тому, что требует естественных для него движений и действий.
Еще один крупный успех этологов в изучении научения (основанный именно на предложенном Лоренцем подходе) был достигнут в исследованиях формирования птичьей песни, начатых Торпом и продолженных многими другими. Успех был связан именно с тем, что не только песня в целом, но и отдельные ее элементы имеют определенную форму, пригодную для выделения и опознания. Позднее, используя наработанные в этой области методы, этологи сумели добиться впечатляющих успехов в расшифровке акустических коммуникативных систем ряда животных (сусликов, белоносых мартышек, дельфинов и т. д.).
Эти достижения позволили значительно раздвинуть границы применимости этологического метода – но не отменили их вовсе. Области, где можно было хотя бы опосредованно опереться на морфологический подход, где-то все же заканчивались, и этологический метод повисал в невесомости. Двигаться в которой, как известно, можно только путем отбрасывания чего-то существенного.
Такие попытки вскоре были предприняты следующим поколением исследователей, но об этом – несколько позже. А пока этологии предстояло пережить свой триумф.
Тут и сказке конец
Сюжеты из истории науки в учебниках и популярной литературе обычно выглядят стройными и законченными. Время все расставляет по местам, и мы сегодня уже точно знаем, кто был прав в яростных дискуссиях прошлого, а кто заблуждался; какая безумная гипотеза обеспечила прорыв в познании, а какие весьма правдоподобные и почти очевидные утверждения оказались ни на чем не основанным недоразумением. Почти как в сказке, где с самого начала ясно, кто герой и кто злодей, а в конце все непременно станут жить-поживать да добра наживать.
В октябре 1973 года в науке о поведении животных случился такой счастливый финал: основоположникам этологии Конраду Лоренцу и Николаасу Тинбергену (а также Карлу фон Фришу – первооткрывателю «языка танцев» у пчел) была присуждена Нобелевская премия по физиологии и медицине. Это стало наглядным выражением полного признания созданной ими теории и ее победы над альтернативными концепциями. Обычно все популярные изложения этологии и ее истории (да и вообще истории изучения поведения животных) заканчиваются именно этим звездным часом. О дальнейших событиях в лучшем случае повествует какая-нибудь ритуальная фраза, вроде «сегодня их идеи развивают многочисленные ученые во всем мире».
Но в науке никогда не бывает окончательного исхода. Любое открытие, любая теория – не последний взмах кисти мастера, завершающий картину, а путь к новым проблемам и новым открытиям. Что происходило с увенчанной нобелевскими лаврами теорией, да и вообще с науками о поведении в эти сорок с лишним лет?
Конечно, включать сегодняшнее состояние той или иной науки в рассказ о ее истории несколько рискованно. Исторический взгляд неизбежно выявляет некие тенденции, которые невольно хочется продолжить в будущее. Но никому не известно, какие из сегодняшних концепций окажутся плодотворными, а какие скоро будут сданы в архив. И тем не менее мы попытаемся это сделать, так как полагаем, что современную ситуацию в науках о поведении трудно понять вне их новейшей истории.
Итак, Лоренцу и Тинбергену была присуждена Нобелевская премия. Чтобы в полной мере оценить значение этого решения, следует учесть, что оно фактически было нарушением правил. Ни зоология, ни психология в завещании Альфреда Нобеля не значились, и основатели этологии до сих пор остаются единственными зоологами (и вообще натуралистами) среди нобелевских лауреатов. Премия была присуждена в номинации «физиология и медицина», но к физиологии работы новоиспеченных лауреатов имели в ту пору отношение весьма косвенное, а к медицине – и вовсе никакого. С учетом этого решение Нобелевской ассамблеи Каролинского института в Стокгольме выглядит как утверждение общенаучной и общекультурной значимости этологической теории, указание на нее как на своего рода маяк для будущих физиологических исследований.
Казалось бы, после такого триумфа этология просто обречена на расцвет. В самом деле, слово «этология» становится чрезвычайно популярным. Число публикаций, где оно в тех или иных сочетаниях употребляется, стремительно растет – как и число исследователей, именующих себя этологами, учебных курсов этологии в университетах, учебников по этой дисциплине и т. д. Привычным становится словосочетание «классическая этология» – так теперь называют работы Лоренца, Тинбергена и их ближайших последователей. Эпитет «классическая» не только выражал почтение к достижениям основателей, но и отражал необходимость как-то отличать этологию в узком смысле слова (как совокупность определенных теоретических взглядов и методов) от этологии в широком смысле – под которой в это время стали понимать едва ли не всякое изучение поведения животных. Если когда-то Кеннет Спенс с удовлетворением констатировал, что «сегодня практически все психологи готовы назвать себя бихевиористами», то спустя несколько десятилетий можно было сказать, что все исследователи поведения готовы назвать себя этологами.
И точно так же, как время, когда «все были бихевиористами», знаменовало застой в бихевиористской теории и канун кризиса, время, когда «все стали этологами», оказалось отмечено сходными симптомами в фундаментальной этологии. Общее число работ, относимых к рубрике «этология», действительно росло, но среди них было все меньше полевых исследований тех или иных конкретных форм поведения, особенно применяющих сравнительный подход. (Это тем более удивительно, что именно теперь, в 1970-е, множеству исследователей стала доступна аппаратура, позволяющая надежно документировать наблюдения: магнитофоны, фотоаппараты с мощной оптикой, а затем и видеокамеры.) Те работы, которые все-таки появлялись, тонули в методологических частностях, в обсуждении, насколько естественным и типичным является наблюдаемое поведение (например, не беспокоило ли наблюдаемых животных стрекотание кинокамеры), правильно ли были выделены ключевые позы и демонстрации и т. д. И самое главное – по прочтении почти любой такой работы возникал вопрос: ну и что?
Классическая этология оказалась прорывом в познании потому, что она позволяла интерпретировать с единых позиций самые разные формы поведения самых разных существ. Но именно этот универсализм теперь оборачивался против нее: типовая работа по этологии обычно представляла собой приложение классических теоретических схем к очередному ранее не исследованному в этом отношении виду (причем почти всегда речь шла только о взаимодействии между особями: брачном, агрессивном, иерархическом и т. п.). Схемы, разумеется, прилагались вполне успешно, все необходимые формы и элементы поведения у изучаемого вида неукоснительно обнаруживались. Правда, другой исследователь, обратившись к тому же самому виду и даже к тому же аспекту его поведения (например, брачному), мог выделить в нем совершенно другой набор элементов – но тоже, конечно, полностью соответствующий теории. Складывалось впечатление, что в этой науке никогда уже ничего не будет, кроме интерпретации поведения все новых видов в категориях классической теории.
Разумеется, время от времени появлялись работы, описывающие необычные, «не лезущие в теорию» феномены. Некоторые исследователи пытались сделать процедуру выделения ключевых элементов поведения объективной и проверяемой. Другие призывали к пересмотру тех или иных положений классической теории или даже к отказу от нее (не предлагая, правда, взамен ничего внятного). Однако от научной теории не отказываются «просто так», из-за того только, что работать в ее рамках стало скучно. Для такого отказа нужна новая, более мощная теория, включающая в себя старую на правах частного случая либо предлагающая совсем иную (и желательно – более плодотворную) интерпретацию всего известного массива фактов. Во всяком случае, именно это утверждает концепция научных революций, разработанная философом Томасом Куном незадолго до описываемого времени и как раз к интересующему нас периоду достигшая пика популярности.
Ничего подобного в науках о поведении не произошло до сих пор. Никто не предложил ни альтернативы классической этологической теории, ни даже ее радикальной модернизации. Можно сказать, что этология стала живой иллюстрацией одного из «законов Паркинсона»: крупный успех в какой-либо области исследований приводит к притоку в нее кадров, финансирования, административных возможностей и других ресурсов, и этот приток делает невозможным дальнейшее продвижение в данной области.
Между тем рядом с этологией и в значительной мере в «силовом поле» ее идей начали формироваться новые направления исследований, быстро ставшие многолюдными и модными. Исследователей, особенно молодых, манила открывавшаяся в этих направлениях возможность обойти ограничения, в которые уперся этологический метод, сохранив в то же время его эвристические возможности.
Гены строят планы,
или Экономика поведения
Начнем несколько издалека. Как мы помним, характерное для того или иного вида поведение так же обусловлено генами, как и характерные особенности строения, и представляет собой такой же результат предшествующей эволюции. Но, согласно теории Дарвина, в эволюции должны складываться и развиваться те признаки, которые обеспечивают наиболее эффективную передачу генов их носителей следующим поколениям. Между тем с давних пор было известно, что у так называемых общественных насекомых – пчел, муравьев, термитов и т. д. – большая часть особей вовсе отказывается от размножения, посвящая себя заботе о размножающихся членах семьи и их потомстве. Как же могло возникнуть такое поведение?
Первую попытку ответить на этот вопрос предпринял в середине 1960-х годов английский теоретик Уильям Хэмилтон. Он исходил из того, что у особей, связанных близким родством – например, у родителей и детенышей или у родных братьев и сестер, – значительная часть генов общая. Таким образом, ген, побуждающий своего обладателя к заботе о родичах, с некоторой вероятностью будет содействовать выживанию и размножению собственных копий. Если необходимые для этого действия не слишком сильно снизят вероятность перехода этого гена в следующее поколение «прямым путем» (через размножение особи-носителя), то такое поведение может быть поддержано отбором.
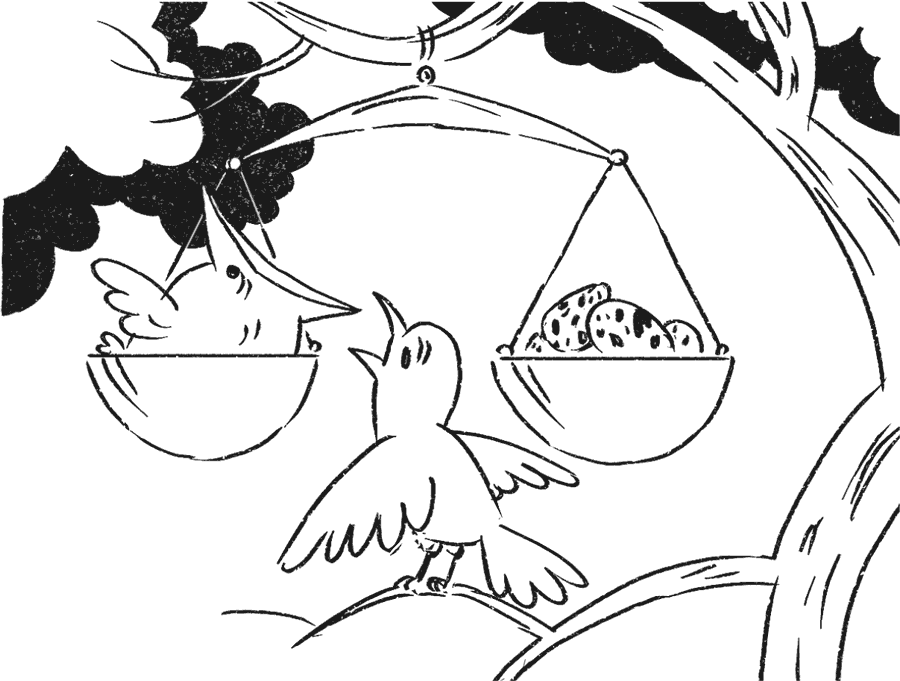
На первый взгляд кажется, что полный отказ от размножения вроде бы не может быть выгоден ни при каких условиях: ведь вероятность «косвенной» – через родственников – передачи гена никогда не будет стопроцентной. Но ведь при половом размножении и родные дети не являются точной генетической копией родителей: для каждого родительского гена вероятность попасть в геном данного конкретного детеныша – всего ½. Между тем именно у общественных перепончатокрылых – пчел и муравьев – работает довольно оригинальный механизм определения пола. Самцы развиваются из неоплодотворенных яиц и в силу этого несут не двойной, а одинарный (гаплоидный) набор генов. Они полностью передают его всем своим детям (а их детьми могут быть только дочери – ведь из оплодотворенного яйца неизбежно разовьется самка), а значит, половина генов у всех самок одной семьи гарантированно одинакова. Вторая половина, получаемая от матери, совпадает, как и положено, в среднем на 50 %, и, таким образом, у двух наугад взятых шестиногих сестер будет примерно ¾ общих генов. Получается, что гипотетическому гену выгоднее побуждать свою обладательницу заботиться о юных сестрах, нежели заводить собственных детей, с которыми у нее общих генов будет только половина!
Такое объяснение происхождения эусоциальности (так называется тип сообщества, члены которого разделены на размножающихся и рабочих), конечно, не может считаться универсальным: у термитов, например, всей этой генетической экзотики нет и в помине, а эусоциальность характерна для всех видов без исключения, в то время как многие виды перепончатокрылых (в том числе пчел) ведут одиночный или «малосемейный» образ жизни, при котором все особи способны к размножению. Но простота и изящество объяснения сложнейшего эволюционного феномена завораживали – тем более что в работах Хэмилтона это объяснение было частным следствием из предложенного им ответа на фундаментальный вопрос: что такое «приспособленность» и как ее можно измерить? Подход Хэмилтона – объяснять особенности социального поведения животных, исходя из соображений эволюционного выигрыша, поддающегося строгому расчету, – выглядел очень заманчиво, и многие исследователи устремились по этому пути. В 1975 году уже довольно известный к тому времени американский энтомолог, специалист по муравьям Эдвард Уилсон выпустил книгу с амбициозным названием «Социобиология. Новый синтез». В ней, по сути дела, была предложена обширная и потенциально плодотворная программа исследований, основанная на подходе Хэмилтона. С легкой руки Уилсона это направление стало именоваться социобиологией.
Новое направление и в самом деле предложило убедительные решения для широкого круга феноменов, выглядевших необъяснимыми в рамках классической этологии, – причем не только в области социального поведения, для которого оно было предложено, но и в других областях. С его позиций, например, становилось совершенно ясно, почему застигнутый на открытом месте леопард безропотно отдает добычу даже одиночной гиене, отнюдь не превосходящей его силой и вооруженностью и значительно уступающей ему в ловкости. Или почему певчие птички довольно часто выкидывают из гнезда яйцо кукушки (если оно сколько-нибудь заметно отличается от их собственных яиц), но никогда не выкидывают и не отказываются кормить кукушонка, разительно непохожего на их птенцов.
Социобиологический взгляд выявлял в этих странных на первый взгляд феноменах четкую логику: для хищника-одиночки (такого как леопард) любая серьезная травма – например, перекушенная лапа – может оказаться смертельной: пока перелом срастется, леопард может умереть с голоду. Кроме того, гиены живут стаями, и даже если на леопарда наткнулась одна, остальные наверняка где-то неподалеку и появятся на месте схватки через несколько минут – а тогда уже не то что добычу сохранить, а и собственную-то шкуру спасти будет нелегко. Леопарду выгоднее отдать задавленную антилопу наглому рэкетиру и поймать себе другую – для такого совершенного хищника это не составит труда и не связано ни с каким риском. При этом никакого инстинктивного страха перед гиенами у леопарда нет, и если ему представится другой выход (например, дерево или скала, куда можно затащить добычу), он без колебаний им воспользуется.
Не менее убедительное объяснение социобиология дает и противоречивому на первый взгляд отношению птиц к кукушачьему потомству. Оно основано на том, что, решая вопрос о выбрасывании или оставлении подозрительного объекта, птица может ошибиться двояким образом: при чересчур строгих «стандартах» она может выбросить собственное яйцо, при чересчур мягких – оставить кукушечье. На этапе яиц цена первой ошибки намного меньше, чем второй: в только что снесенное яйцо вложено еще не так уж много ресурсов и усилий, в то время как оставление в гнезде «троянского коня» будет означать гибель всего выводка и напрасно потраченный сезон. Значит, на этом этапе естественный отбор будет поддерживать максимальную придирчивость птиц к облику своих яиц. К моменту же вылупления птенцов времени и сил именно в этот выводок вложено уже гораздо больше, и бросить по ошибке своего птенца означает слишком большой проигрыш. С другой стороны, если это все-таки кукушонок, отказ от его выкармливания уже никаких особенных выгод не дает: это означает, что собственный выводок уже погиб и терять все равно нечего. Часто уже упущено и время для обзаведения вторым выводком. Поэтому на этом этапе бросить по ошибке своего птенца означает куда больший проигрыш, чем продолжать кормить паразита, и отбор поддерживает «нетребовательность» птиц к облику птенца.
Можно сказать, что социобиология, будучи естественным продолжением и обобщением классической этологии, одновременно оказалась ее отрицанием. Полностью разделяя взгляд на поведение как на продукт эволюции и средство индивидуальной и видовой адаптации, социобиологи, однако, сделали из этого тезиса гораздо более категоричные выводы. Как мы помним, одним из главных вопросов этологии был вопрос о соотношении врожденных (видоспецифичных) и индивидуально приобретенных компонентов поведения: занимаясь в основном врожденным поведением (и достигнув наибольших успехов именно в этой области), этологи тем не менее никогда не пытались свести к нему все поведение животных. При этом они настаивали на строгом разграничении врожденного и приобретенного (даже внутри одного целостного акта), причем не только в теории, но и в методике: приступая к изучению того или иного поведения – например, пения птицы, – необходимо первым делом выяснить, какие его элементы наследуются, а каким птица обучается (и каков механизм этого обучения). Социобиология же предлагает рассматривать любое сколько-нибудь регулярно наблюдаемое поведение как «хотя бы немножко врожденное». В самом деле, способность к любому поведению в конечном счете обеспечивается работой генов, и совершенно невозможно себе представить, чтобы все версии (аллели) всех генов обеспечивали эту способность абсолютно одинаково. Возвращаясь к примеру с птичьим пением: допустим, мы изучаем вид, у которого никакие особенности видовой песни не наследуются, и каждый самец этого вида «сочиняет» свою песню сам, комбинируя в ней элементы песен других видов и вообще любые понравившиеся звуки. (Такие птицы действительно существуют; собирательно их называют пересмешниками.) Но разные особи в разной мере способны к обучению, и эти различия отчасти определяются генетически. Некоторая генетическая основа есть, вероятно, и у разницы «художественных вкусов» (кому какие звуки кажутся достойными включения в песню) и других индивидуальных особенностей. А раз так, то мы имеем полное право рассматривать «вокальное поведение» пересмешников как признак, подлежащий действию естественного отбора, – и смотреть с этой точки зрения, какие особенности песни оказываются эволюционно выигрышными, а какие – проигрышными, не выясняя детально, что именно наследуется в данном случае.
Казалось бы, такое рассуждение – блестящая находка, позволяющая распространить прекрасно зарекомендовавшие себя методы этологии практически на любые формы поведения. Однако при подобном обобщении неизбежно теряется основа этологического метода – морфологический подход к поведению.
В рамках социобиологического подхода любое поведение рассматривается исключительно с точки зрения его эволюционной выгоды – то есть сугубо функционально. Это не только закрывает для исследования важнейший аспект поведения, но и делает социобиологические объяснения тех или иных поведенческих феноменов гораздо более абстрактными по сравнению с объяснениями этологическими. Там, где этолог должен хотя бы наметить возможный эволюционный путь формирования той или иной особенности поведения, социобиолог неизбежно ограничивается лишь указанием на его причину – эволюционную выгоду. Это диктуется самим подходом и не зависит от желаний применяющего его исследователя, но социобиологи часто рассматривают такое ограничение как своего рода преимущество: мы, мол, занимаемся не «проксимальными механизмами» (к коим относится любая биологическая конкретика – от групп атомов, по которым клеточный рецептор узнает «свой» гормон, до признаков, по которым мать узнает своего детеныша), а «конечными объяснениями», первопричинами того или иного поведения.
Отсюда неизбежно следует еще одна особенность социобиологических трактовок: в них не остается места для неадаптивного поведения – рудиментов былых приемов (вроде «львиного захвата» у кошек – см. главу 4), неадекватного срабатывания инстинктов, наконец, просто ошибок (например, опознания). С точки зрения социобиолога любое регулярно наблюдаемое поведение в конечном счете выгодно – если не самому животному, то тем генам, которые обеспечивают именно такое поведение; если не в данном конкретном случае, то статистически. Скажем, нерест лосося или массовый лет муравьиных самцов и самок привлекает множество хищников, до отвала наедающихся легкодоступной и питательной добычей. Во время хода лососей все бурые медведи, живущие в бассейнах нерестовых рек, выходят на берега и добрых полмесяца кормятся только рыбой и икрой. Но при всей своей прожорливости они не могут сожрать больше лососей, чем вмещают их желудки. Если бы лососи кишели в реках круглый год, медведи могли бы увеличить свою численность. Но размножиться за пару недель звери не могут – да и чем им тогда питаться весь остальной год? Получается, что чем короче сезон нереста, тем меньшую долю всех пришедших на нерест рыб сожрут хищники – и тем, следовательно, выше шансы каждой конкретной рыбы успеть оставить потомство. Естественный отбор не ошибается – он всегда поддерживает поведение, увеличивающее вероятность распространения данных генов, даже если человеку такое поведение кажется глупым и самоубийственным.
Часто такой подход и в самом деле позволяет понять, почему в эволюции возникают и сохраняются, казалось бы, невыгодные формы поведения. Например, сойка – птица неглупая, не обиженная ни памятью, ни способностью ориентироваться в пространстве – имеет привычку прятать избыток желудей, большинство которых потом никогда не находит. Но в конечном счете это служит к ее же пользе: зарытые и забытые желуди прорастают, превращаясь в дубы – кормовую базу для будущих поколений соек.
Лукавая метафора
Тем не менее установка «что ни делает животное – все к лучшему» при одновременном исключении из рассмотрения конкретных поведенческих механизмов в конце концов неизбежно возвращает мысль ученых к натурфилософии XVIII века с ее представлениями о «бесконечно предусмотрительной Природе». Образцом такого способа теоретизирования может служить, например, весьма популярная в социобиологической литературе «теория гандикапа» израильского орнитолога Амоца Захави.
Как известно, у самцов многих видов животных имеются признаки, явно не способствующие выживанию их обладателя, такие как хвост самца павлина, огромные рога оленей, глаза на длинных стебельках у стебельчатоглазых мух Cyrtodiopsis dalmanni и т. д. Со времен Дарвина возникновение столь неадаптивных признаков принято объяснять половым отбором: подобные украшения привлекают самок, и потому проигрыш в безопасности с лихвой компенсируется репродуктивным успехом. В последние десятилетия прямые полевые эксперименты показали, что по крайней мере в некоторых случаях такое объяснение справедливо: если самцу длиннохвостой вдовушки Euplectes jackstone укоротить его несообразно длинный хвост, он станет гораздо менее привлекателен для самок; если же с помощью клея и чужих перьев надставить хвост дополнительно, любой невзрачный самчишка станет неотразимым. Однако столь же убедительно было показано, что подобные предпочтения самок в значительной мере определяются генетически и, следовательно, находятся под действием естественного отбора. Почему же он не «вразумит» любительниц экстравагантных форм и не привьет им вкус к скромным, но функциональным нарядам кавалеров?
Захави предположил, что выбор самок имеет глубокий смысл: если уж самец с таким хвостом (рогами, глазными стебельками и т. д.) ухитрился дожить до брачного возраста – значит, какие-то не менее важные, но скрытые, не воспринимаемые непосредственно достоинства (например, устойчивость к холоду, жаре, болезням, эффективность утилизации пищи и т. д.) у него, скорее всего, намного выше среднего. А значит, и дети от такого отца будут самыми лучшими – особенно дочери, у которых эти хвосты-рога в любом случае не вырастут и которым, следовательно, не придется тратить на них ресурсы. Получается, что гипертрофированные структуры – это фора, которую их обладатели дают другим самцам в борьбе за существование. И именно по этой форе самки опознают в них сильных игроков и стремятся заполучить их в мужья. Отсюда и название – «теория гандикапа», то есть форы.

Звучит вроде бы убедительно, но неужели пава при виде павлина с особенно роскошным хвостом проделывает в уме все эти логические выкладки? Любой социобиолог с негодованием отвергнет такое предположение. «Чтобы сделать подобные определения более ясными и сократить их, биологи и прибегают к антропоморфным уподоблениям: например, говорят, что животное „выбирает“ сделать то-то и то-то или следует определенной стратегии. Эти метафоры не должны ввести читателя в заблуждение и заставить его думать, будто животные совершают сознательный выбор… поступки каждого из них генетически запрограммированы, они обусловлены их анатомией и инстинктами», – поясняет подобные фигуры речи известный орнитолог и популяризатор Джаред Даймонд. Иными словами, пава никаких расчетов не делает – просто естественный отбор на протяжении многих поколений благоприятствовал тем павам, которым нравились ухажеры с самыми длинными хвостами.
Но как он мог это делать? Предполагается, что оба признака – необычно пышный хвост самца и слабость самки именно к таким хвостам – независимы и должны появиться случайно в одной и той же популяции и более-менее в одно и то же время (чтобы, когда носитель одного из них войдет в брачный возраст, носитель другого не успел из него выйти). При этом первый обладатель пышного хвоста отнюдь не обязательно будет иметь какие-то «скрытые достоинства»: это лишь статистическая корреляция, проявляющаяся только в большом числе случаев. Их пока нет, а значит, отбор пока не будет благоприятствовать «гену любви к хвостам». В то же время сам по себе, вне выбора самок, такой хвост скорее вреден (демаскирует, ухудшает летные качества). Значит, пока такие хвосты нравятся только одной самке-мутантке, отбор не будет содействовать и распространению «гена пышнохвостости», даже скорее будет работать против него. Получается замкнутый круг: пышный хвост не дает преимущества, пока нет множества предпочитающих его самок, а тяга к таким хвостам бесполезна, пока нет множества обладающих ими самцов. А естественный отбор – не профессор социобиологии, просчитывать выгоды заранее он не умеет и поддерживает только то, что выгодно здесь и сейчас.
Между тем в гипертрофии «гандикапных» структур нет ровным счетом ничего загадочного, если вспомнить о феномене сверхстимулов (см. главу 4). Мы уже говорили, что, как только тот или иной признак начинает выполнять функцию полового релизера, его дальнейшая эволюция неизбежно будет идти в сторону наибольшей выраженности, по сути дела – в сторону приближения к сверхстимулу. То, что с человеческой точки зрения результат такой эволюции может выглядеть довольно гротескно, – проблема не эволюции, а человеческого восприятия. (Как мы помним, «идеальный образ мамы-чайки» в представлении птенцов вовсе не похож на то, как видит чайку человек.) Нарастание выраженности признака будет идти до тех пор, пока либо он не совпадет со сверхстимулом, либо связанные с этим неудобства и опасности не перевесят выигрыша в привлекательности.
Косвенным образом это подтверждается тем, что «гандикапные» признаки и даже выраженный половой диморфизм редко встречаются у видов, где процесс формирования пары достаточно длителен, и совсем не встречаются у тех, у кого в выращивании потомства участвуют оба родителя и вклад самца существенен для выживания детей. Это можно наблюдать даже у видов, принадлежащих к одному семейству, – например, куриных. Участие самцов павлинов, фазанов, индюков в продолжении рода заканчивается совокуплением, при этом один самец в один сезон может осчастливить своими генами изрядное число самок – если будет достаточно привлекателен. Это уравновешивает повышенную вероятность его гибели из-за обремененности гротескными признаками. С точки зрения естественного отбора риск оправдан и относительно невелик: дополнительная опасность грозит только самцу, его многочисленные дети вырастут без него так же успешно, как и с ним. А вот самцу куропатки во время насиживания предстоит подменять подругу на гнезде, а потом на пару с ней (а если с ней что-то случится, то и одному) водить и беречь птенцов до подъема на крыло. Демаскирующие «гандикапные» признаки создали бы дополнительный риск не только для него самого, но и для его птенцов. И у куропаток мы не видим ни пышных хвостов, ни яркой, бросающейся в глаза окраски. Петушок куропатки отличается от курочки разве что скромными красными «бровями», да и они заметны лишь в сезон активного образования пар. Хотя вроде бы логика «гандикапной гипотезы» должна быть одинаково справедливой для куропаток и павлинов.
Но вернемся к теориям социобиологов. Теория Захави – не единственный пример того, как в их построениях разговор об объективных эволюционных выгодах то и дело незаметно съезжает на субъективные расчеты и прикидки индивида. Такова же, например, теория «многих отцов», предложенная антропологом Сарой Хрди из Калифорнийского университета в Дэвисе для объяснения происхождения скрытой овуляции у предков человека. По мнению Хрди, скрытая овуляция нужна для предотвращения инфантицида: самцы, имеющие дурную привычку убивать детенышей, рожденных самкой от других самцов, не будут это делать, зная, что это может быть их собственный отпрыск. Нетрудно показать, что в этом случае никакие оговорки насчет «метафор» и «простоты изложения» не работают: такая защита детей могла бы быть эффективной только в том случае, если потенциальный детоубийца сознательно заботится о том, как бы ненароком не убить собственного детеныша. На то, что не воспринимается органами чувств, а лишь логически допустимо, не может срабатывать никакой инстинкт.
С годами все отчетливее стала проявляться еще одна неустранимая слабость социобиологии. Схемы, в которых фигурировали «ген альтруизма», «ген групповой сплоченности», «мутации, способствующие склонности всерьез влюбляться», и тому подобные абстракции, выглядели красиво и убедительно до тех пор, пока невозможно было проследить механизмы связи между генами и поведением. Было понятно, что первые сильно влияют на второе, но как именно они это делают – оставалось совершенно неизвестным. Конечно, специалисты и тогда понимали, что любой поведенческий акт – результат работы множества разных генов, сложным образом взаимодействующих между собой и с сигналами из окружающей среды. Но почему бы не предположить, что один аллель некого гена повышает вероятность, скажем, проявления заботы о потомстве по сравнению с другим аллелем того же гена? Тогда можно будет оперировать этой «заботой о потомстве» как старым добрым менделевским наследственным признаком – разумеется, произнося время от времени ритуальные оговорки насчет «склонности», «предрасположенности» и «сложной организации генетического контроля поведения». Более того – предполагаемую эволюцию каждого такого гипотетического «гена» и связанного с ним поведения можно моделировать изолированно, отдельно от эволюции других аналогичных «генов», рассматривая все поведение организма как мозаику почти независимых функциональных блоков. Это сулит головокружительные перспективы: ведь мы до сих пор не знаем, как именно кодируются в геноме врожденные программы поведения, да и вообще не очень умеем работать с целостными генетическими программами. А вот с отдельными генами, да еще и представленными разными версиями, работать куда легче.
Туман, скрывающий цепь причин и следствий между последовательностью нуклеотидов в молекуле ДНК и поведением обладателя этой молекулы, все еще весьма плотен, но сегодня нам иногда удается различить в нем контуры отдельных звеньев этой цепи. И почти всякий раз при этом оказывается, что конкретные нейробиологические и нейрохимические механизмы невозможно отождествить ни с какими элементами социобиологических схем – и наоборот.
Известно, например, что в Х-хромосоме человека (и других млекопитающих) среди прочих генов есть ген, кодирующий фермент моноаминоксидазу А (МАО-А). Этот фермент занимается утилизацией некоторых нейромедиаторов и гормонов. У человека ген МАО-А известен в двух версиях, различающихся своей регуляторной частью (промотором). Это приводит к тому, что у обладателей одной версии гена активность МАО-А заметно ниже, чем у обладателей другой. Поскольку этот ген находится в Х-хромосоме, у мужчин есть только одна его копия. Так вот, оказывается, что обладатели «неактивной» версии более склонны к насилию и нарушению социальных норм.
Казалось бы, вот прекрасная модель для социобиологического подхода: ген, который без всякой натяжки можно назвать «геном агрессии» или, наоборот, «геном социализации». Но как ее истолковать? Если мы предполагаем, что эволюция человека шла в сторону усиления социального контроля над агрессивностью и сексуальностью, то придется допустить, что первичной была именно «неактивная» версия. Но сам характер различия говорит о том, что исходная форма – именно «активная», а «неактивная» – результат мутации. Но как тогда она смогла так распространиться (среди белых американцев частота «неактивной» версии составляет 33 %, а в некоторых восточноазиатских популяциях превышает 60 %)? Получается, что генные частоты менялись в одну сторону, а поведение их обладателей эволюционировало в прямо противоположную!
В довершение всего обнаружилось, что эффект «неактивной» МАО-А сильно зависит от обстоятельств начального периода жизни индивидуума: те носители «неактивной» версии фермента, которые выросли в нормальных любящих семьях, склонны к насилию не больше, чем счастливые обладатели «активной» МАО-А. Это ставит под сомнение корректность уподобления социобиологически значимых качеств менделевским признакам – и уж во всяком случае не позволяет нам рассматривать поведение как механическую результирующую работы того или иного числа независимых друг от друга генов. А ведь это всего лишь одно конкретное исследование, выхватившее одно-единственное звено в цепочке неизвестной длины!
Все это приводит к нарастающему разочарованию исследователей поведения в социобиологическом подходе. Хотя сегодня он остается чрезвычайно распространенным и в самом деле позволяет решать множество частных задач и выявлять причинно-следственные связи, невидимые с иных точек зрения, уже ясно: по-настоящему глубоких прорывов в понимании поведения животных от социобиологии ждать не приходится.

