Книга: Век самопознания. Поиски бессознательного в искусстве и науке с начала XX века до наших дней
Назад: Глава 14 Обработка мозгом зрительных образов
Дальше: Глава 16 Реконструкция видимого мира: зрение и обработка информации
Глава 15
Деконструкция зрительных образов: «строительные блоки» восприятия форм
Зрение играет в нашей жизни исключительно важную роль. Мир, в котором мы живем, – во многом видимый. Поиски пищи и питья, партнеров и друзей осуществляются при помощи информации, получаемой сетчаткой глаз. Сетчатка поставляет около половины всей сенсорной информации, поступающей в мозг. Если бы у нас не было зрения, не существовало бы и изобразительного искусства, а наше сознание, вероятно, было бы более ограниченным. Так что неудивительно, что биологи, наряду с художниками и философами, а также искусствоведами, психологами и другими учеными, давно интересуются устройством зрения.
У истоков биологических исследований зрительного восприятия стоит Стивен Куффлер – выдающийся ученый, связанный с венской интеллектуальной традицией, современник Эрнста Криса и Эрнста Гомбриха. В 50-х годах XX века Куффлер и его младшие коллеги Дэвид Хьюбел и Торстен Визель занялись изучением деконструкции образов в ходе обработки мозгом зрительной информации. (Этим же занимались в свое время Крис и Гомбрих.) Куффлер, Хьюбел и Визель изучили реакции нейронов зрительной системы на различные стимулы и подготовили почву для перехода от когнитивной психологии восприятия к его биологическому анализу. Их исследования позволили ответить на ряд принципиальных вопросов. Имеются ли в мозге специфические клетки, ответственные за кодирование базовых фигур, этих “строительных блоков” всех видимых форм? И если да, возникают ли наши представления о видимых формах в результате совокупной активности этих клеток? Где именно осуществляется реконструкция изображений, деконструкция которых происходит на сетчатке?
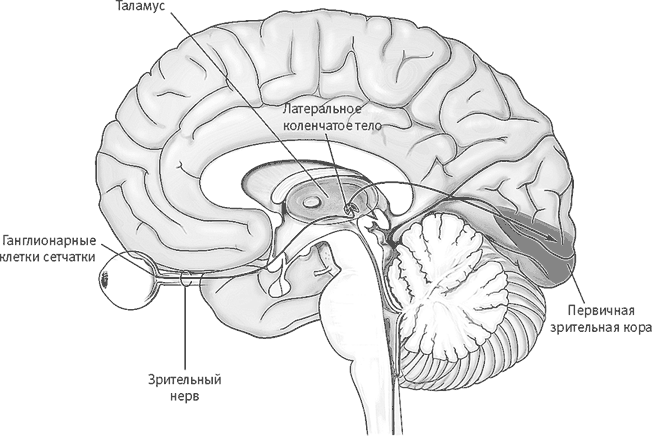
Рис. 15–1. Упрощенная схема передачи зрительной информации от сетчатки в зрительные отделы таламуса (латеральное коленчатое тело), а затем в зрительную кору.
Обработка зрительной информации начинается на сетчатке и продолжается в латеральном коленчатом теле таламуса и трех десятках зрительных зон коры больших полушарий (рис. 15–1). Куффлер, Хьюбел и Визель показали: наше сознательное восприятие отдельных аспектов видимых образов обеспечивают последовательности сигналов, посылаемых нейронами зрительной системы. Ученые выяснили, что нейроны низших отделов зрительной системы (сетчатки и латерального коленчатого тела) особенно эффективно реагируют на небольшие пятна света. Нейроны следующего передатчика, первичной зрительной коры (V1, где V происходит от английского названия зрительной коры – visual cortex), извлекают из зрительной информации сведения о контурах, очертаниях и углах, после чего все эти элементы объединяются, образуя представления очертаний и базовых фигур. Дальнейшие передатчики зрительной коры, получающие информацию от зоны V1, специализируются на иных функциях: V2 и V3 реагируют на виртуальные линии и границы, V4 – на цвет, V5 – на движение. Впоследствии другие нейробиологи выяснили, что нейроны нижней височной коры, где располагаются высшие отделы зрительной системы, реагируют на сложные формы (целые сцены, места, руки, тела, особенно лица), а также на цвет, положение в пространстве и движение всех подобных указанным форм.
Чтобы видеть, необходим свет. Видимый свет – одна из разновидностей электромагнитного излучения. Он состоит из волн разной длины, распространяющихся в виде порций (фотонов), которые отражаются от видимых объектов. Человеческое зрение улавливает лишь небольшую часть спектра электромагнитных волн: от 380 (мы воспринимаем такой свет как темно-фиолетовый) до 780 нм (темно-красный). Этот диапазон называют видимой частью спектра (рис. II–17, 15–2).
Фотоны видимого света, отражаемые различными объектами, достигают хрусталика глаза, фокусирующего свет на сетчатке, где его поглощают фоторецепторы – особые нервные клетки. Фоторецепторы реагируют на световые волны определенным образом, зависящим от длины и направления распространения этих волн. Реагируя на свет, фоторецепторы генерируют последовательности электрических сигналов, в конфигурациях которых закодирована информация о нем. Сигналы фоторецепторов передаются на ганглионарные клетки сетчатки. Длинные отростки (аксоны) ганглионарных клеток сетчатки образуют оптический нерв, по которому зрительная информация направляется в первичную зрительную кору (рис. 15–3). Этот механизм позволяет сетчатке получать и обрабатывать сведения и передавать их в другие отделы зрительной системы.
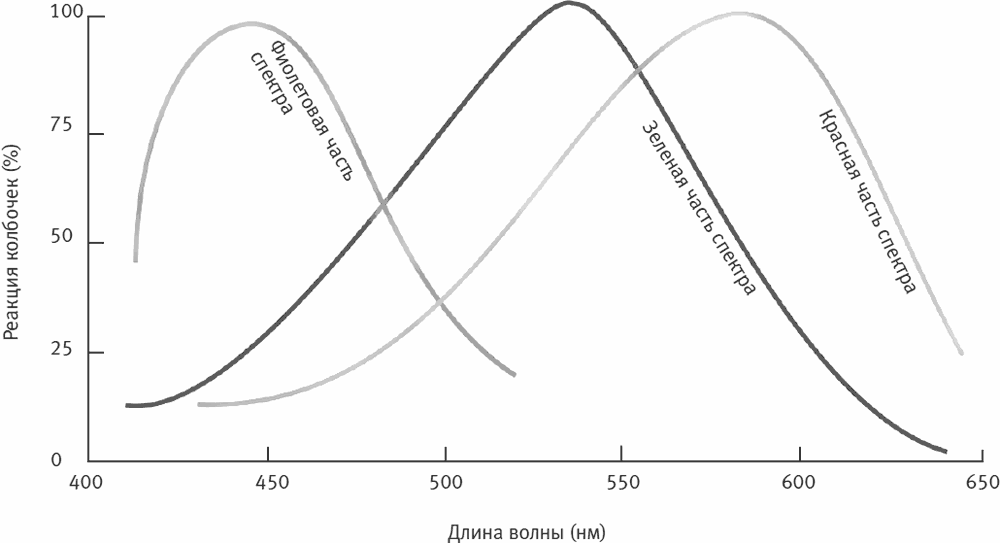
Рис. 15–2. Чувствительность трех типов колбочек.
В сетчатке есть фоторецепторы четырех типов: три типа колбочек и один тип палочек. Колбочки позволяют в подробностях воспринимать окружающее. Это они позволяют нам наслаждаться искусством. Они работают при дневном свете и при ярком искусственном освещении и отвечают за чувствительность глаз к контрасту, цвету и мелким деталям (рис. 15–4). Колбочки разбросаны по сетчатке, а в центральной ямке – самом чувствительном участке сетчатки – других фоторецепторов вовсе нет. Кроме того, в центральной ямке плотность расположения колбочек особенно велика, и от этого участка зависит наша способность различать лица, руки и другие объекты и образы, а также цвета. По направлению от центральной ямки к периферии сетчатки плотность колбочек уменьшается. Разрешение зрения на периферии сетчатки гораздо меньшее, чем разрешение в центральной ямке, а зрительная информация, поступающая с периферии сетчатки, гораздо менее подробна.
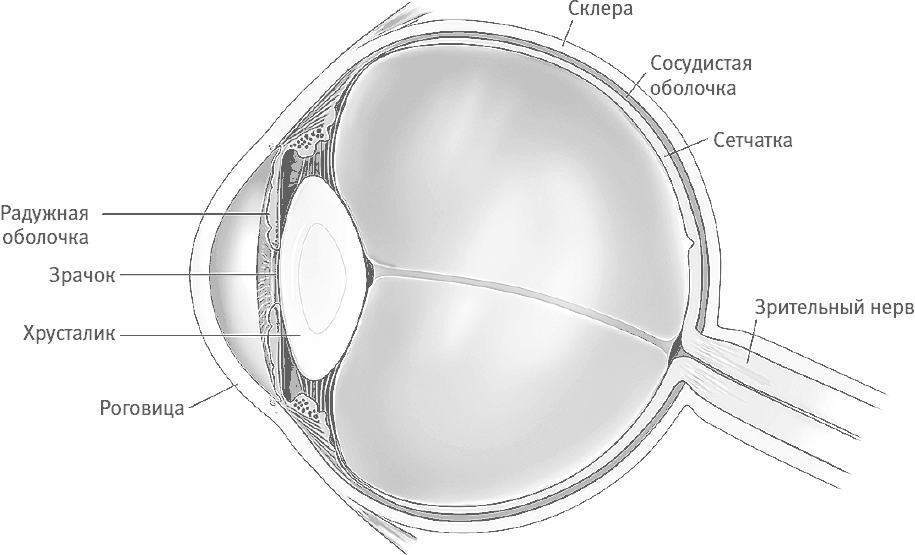
Рис. 15–3. Наружная оболочка глаза (склера) поддерживает его форму. Сквозь роговицу, прозрачную часть склеры, в глаз попадает свет. Цветную часть глаза называют радужной оболочкой. В ней имеется круглое отверстие – зрачок, расширение или сужение которого зависит от перемены яркости света. Свет, проходящий через зрачок, преломляется хрусталиком и фокусируется на сетчатке, выстилающей глаз изнутри.
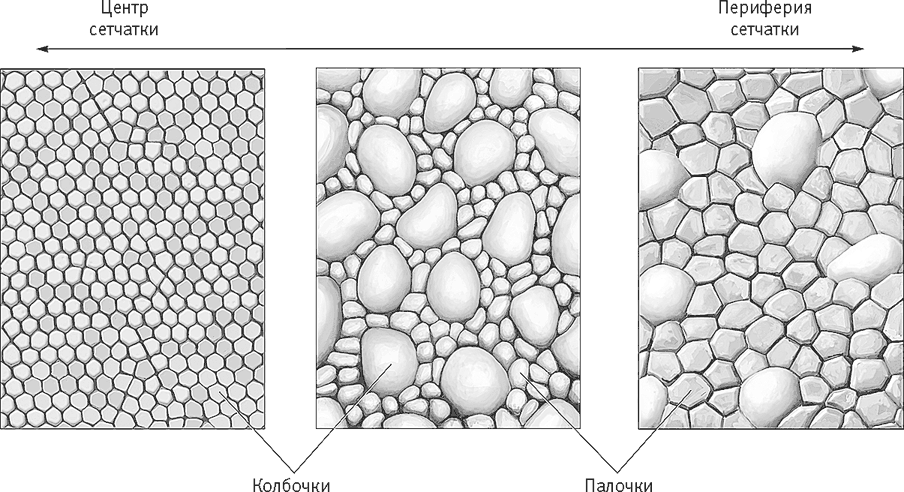
Рис. 15–4. В центре сетчатки из фоторецепторов имеются лишь плотно упакованные колбочки. По направлению к периферии колбочки становятся крупнее и появляются палочки, и чем дальше, тем их становится больше.
Колбочки каждого из трех типов содержат свой пигмент и чувствительны к своему компоненту видимой части спектра: фиолетовому, зеленому или красному. Например, когда мы видим зеленый автомобиль, его поверхность отражает световые волны той длины, которая соответствует зеленой части спектра, а остальные поглощает. На отражаемый этой поверхностью свет реагируют колбочки соответствующего типа, и мозг воспринимает автомобиль как зеленый.
Цветовая чувствительность играет ключевую роль в нашей способности различать видимые объекты. Она позволяет замечать детали. Наряду с чувствительностью к яркости, цветовая чувствительность существенно увеличивает контраст между элементами зрительного образа. Но сами по себе цвета, без изменений яркости, на удивление мало позволяют сказать о подробностях пространственного расположения объектов (рис. II–18).
Кроме того, цвет обогащает эмоциональную жизнь человека. В нашем восприятии цвета обладают эмоциональной окраской, и реакция на тот или иной цвет может меняться в зависимости от настроения. Для разных людей один и тот же цвет также означает разное. Художники, особенно модернисты, часто использовали неестественные цвета ради эмоционального эффекта, но сила и даже характер этого эффекта зависят от зрителя и контекста. Неоднозначное восприятие цветов может быть одной из причин того, что одна и та же картина нередко вызывает у зрителей разную реакцию, даже у одного зрителя в разное время.
Палочек в сетчатке человеческого глаза гораздо больше, чем колбочек (около 100 млн против 7 млн), и при дневном свете или ярком искусственном освещении они неэффективны: слишком сильный свет подавляет их. Кроме того, они не передают информацию о цвете и поэтому в обычных условиях не участвуют в восприятии произведений искусства. Однако палочки гораздо чувствительнее к свету, чем колбочки, и лучше работают при слабом освещении.
За ночное зрение отвечают исключительно палочки. В этом можно убедиться в звездную ночь, присмотревшись к не особенно яркой звезде. Такую звезду бывает сложно разглядеть, если смотреть прямо на нее, потому что колбочки центральной ямки не реагируют на слабый свет. Но если немного повернуть голову и посмотреть на ту же звезду боковым зрением, задействовав палочки на периферии сетчатки, она станет видна вполне отчетливо.
Плотно упакованные колбочки центральной ямки успешнее улавливают мелкие детали, а разреженные колбочки на периферии сетчатки – более грубые. Поэтому зрительная информация обрабатывается в мозге двумя способами: в большем разрешении (детали) и в меньшем (цельные образы). Части образа, используемые для распознавания лиц (например размеры и форма носа), воспринимаются с помощью колбочек центральной ямки, чувствительных к деталям и дающим высокое разрешение, а части, используемые для определения эмоционального состояния, – с помощью колбочек периферии, чувствительных к более грубым компонентам образа (его гештальт-элементам).
Маргарет Ливингстон обратила внимание на эту двойственность восприятия в интересном исследовании “Моны Лизы” (рис. II–19). Внимание, которое уделяли этой картине искусствоведы и психоаналитики, отражает общее отношение к ней как к шедевру и одному из лучших примеров неоднозначности в живописи. Мона Лиза стала символом ренессансного идеала таинственного женского начала, воплощением “вечной женственности” (выражение Гете). Одна из самых очаровательных и вместе с тем загадочных особенностей портрета – это выражение лица Моны Лизы. Какие эмоции выражает ее лицо? Кажется, что она лучезарно улыбается в один момент, а в другой выглядит задумчивой, даже грустной. Как художнику удалось добиться такого эмоционального эффекта?
Эрнст Крис утверждал, что выражение лица Моны Лизы кажется изменчивым оттого, что заложенная в портрете неоднозначность позволяет интерпретировать его по-разному в зависимости от настроения зрителя. Традиционное объяснение “загадки Моны Лизы” гласит, что Леонардо воспользовался особым приемом, изобретенным в начале эпохи Возрождения: сфумато (ит. затушеванный). Техника сфумато подразумевает нанесение полупрозрачной темной краски, подмешивание к ней непрозрачных белил и смягчение резких очертаний – в данном случае в уголках рта.
Маргарет Ливингстон предложила иное объяснение. По ее мнению, выражение лица Моны Лизы связано с двойственностью вложенной в портрет информации. Когда мы смотрим на губы Моны Лизы, мы не сразу замечаем знаменитую улыбку: центральное зрение концентрируется на деталях, а они не свидетельствуют однозначно об улыбке. Но когда мы смотрим на очертания лица или на глаза Моны Лизы, она вполне откровенно улыбается. Это происходит оттого, что периферическое колбочковое зрение, плохо улавливающее детали, воспринимает цельный образ, открывая нам эффект сфумато на губах и в уголках рта (рис. II–20).
Итак, мы можем воспринимать периферическим зрением то, что ускользает от центрального. Поскольку выражение лица зависит от глубоких мимических мышц, а их сокращения могут быть “затушеваны” подкожным жиром, периферическое зрение порой лучше центрального позволяет улавливать выраженные на лице эмоции. Центральное зрение, в свою очередь, позволяет нам легко узнавать в лицах лица по одним лишь их очертаниям.
Научное изучение зрительного восприятия млекопитающих началось со Стивена Куффлера (рис. 15–5). Он родился в 1913 году в Венгрии, входившей в то время в состав Австро-Венгерской империи. В 1923 году Куффлер переехал в Вену, где учился в иезуитском интернате, а в 1932 году поступил на медицинский факультет Венского университета. В 1937 году он окончил университет, где специализировался на патологической анатомии. В 1938 году, когда Гитлер вошел в Вену, Куффлер, член антигитлеровской студенческой организации и на четверть еврей (по бабушке с отцовской стороны), уехал в Венгрию, затем в Англию, а после в Австралию. В 1945 году он поселился в США и стал работать в Глазном институте им. Уилмера при Университете им. Джона Хопкинса. В 1959 году он перешел на медицинский факультет Гарварда, где в 1967 году основал первое в США отделение нейробиологии. Там производились исследования в области физиологии, биохимии и анатомии мозга.
Куффлер в Университете им. Джона Хопкинса изучал механизмы взаимодействия нейронов в мозге беспозвоночных, например речного рака. (Фрейд еще в 1884 году выяснил, что нейроны позвоночных и беспозвоночных устроены очень похоже.) Обычный нейрон состоит из тела клетки, одного аксона и множества дендритов (рис. 15–6). Аксон – тонкий длинный отросток, отходящий от тела клетки – передает информацию, часто на немалое расстояние, на дендриты других нейронов. Густо ветвящиеся дендриты – короткие отростки – обычно отходят от противоположного конца тела клетки и получают информацию, поступающую от других нейронов. Куффлер изучал процесс синаптической передачи информации в синапсах – местах, где контактирует аксон нейрона, посылающего сигнал, и дендриты нейрона, принимающего этот сигнал (клетки-мишени).

Рис. 15–5. Стивен Куффлер (1913–1980).
Нейроны генерируют кратковременные электрические импульсы – потенциалы действия. Стоит импульсу возникнуть, и он без изменений передается по всей длине аксона до его окончания. Здесь аксон образует один или несколько синапсов, связывающих его с клеткой-мишенью. Сила сигнала не меняется при движении по аксону, потому что потенциал действия генерируется на всем протяжении аксона в полную силу, по принципу “все или ничего”. Клетка-мишень получает сигналы и от других клеток. Эти клетки могут быть возбуждающими, сигналы которых увеличивают число возникающих в клетке-мишени потенциалов действия, и тормозными, сигналы которых уменьшают это число. Чем дольше активен возбуждающий нейрон, тем дольше будет активен и нейрон-мишень.
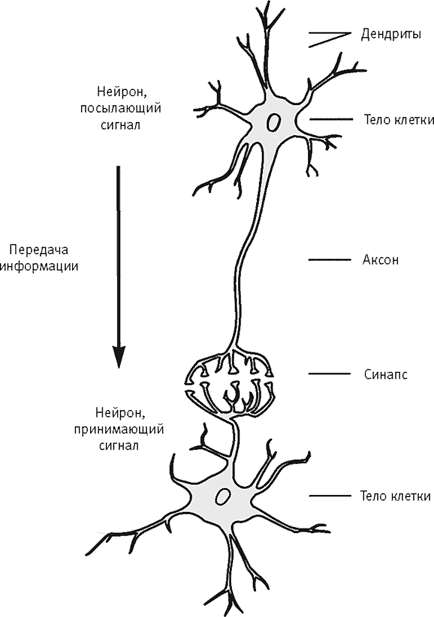
Рис. 15–6.
Куффлер понимал, что взаимодействие возбуждающих и тормозных нейронов, управляющих конфигурацией импульсов одного нейрона-мишени, отражает основной принцип устройства всей нервной системы. Нейроны в мозге суммируют возбуждающую и тормозную информацию, получаемую из разных источников, и определяют, передавать ли информацию дальше. Британский физиолог Чарльз Шеррингтон (получивший в 1932 году Нобелевскую премию за изучение взаимодействия нейронов спинного мозга) называл этот механизм интегративной деятельностью нервной системы. Шеррингтон утверждал, что ключевая функция нервной системы состоит в оценивании сравнительной значимости входящей информации и использовании оценки для принятия решений о действиях.
Результаты экспериментов, посвященных изучению синаптического возбуждения и торможения у раков, вдохновили Куффлера на исследования более сложных взаимодействий нейронов в сетчатке млекопитающих. Он занялся не только механизмами синаптической передачи, но и механизмами обработки информации в одной из сенсорных систем мозга. Как Куффлер впоследствии говорил, он решил разобраться в том, как работает мозг.
Куффлер, а впоследствии Хьюбел и Визель (рис. 15–7) изучали зрительное восприятие животных. Они понимали, что у разных нейронов могут быть разные функции, механизмы работы и свойства, а значит, чтобы разобраться в работе мозга, нужно разобрать его по клетке. Куффлер, а затем Хьюбел и Визель вводили в сетчатку животных микроэлектроды и регистрировали возникающие в клетках сетчатки электрические импульсы. Электроды были присоединены к осциллографу и репродуктору с усилителем, что позволяло наблюдать потенциалы действия клетки на осциллографе и одновременно слышать их треск. Так исследователи изучали, как клетки разных отделов зрительной системы реагируют на элементарные стимулы и как информация трансформируется на пути от сетчатки через передатчики в направлении высших зрительных зон мозга.

Рис. 15–7. Дэвид Хьюбел (1926–2013) и Торстен Визель (р. 1924; справа).
Куффлер начал с регистрации потенциалов действия, генерируемых отдельными ганглионарными клетками в центре и на периферии сетчатки. Он обнаружил, что эти специализированные нейроны получают от колбочек и палочек информацию о зрительных образах, кодируют ее в виде последовательностей импульсов и передают в мозг. Регистрируя эти импульсы, Куффлер сделал первое неожиданное открытие: ганглионарные клетки сетчатки никогда не спят. Они самопроизвольно генерируют потенциалы действия даже без света или какой-либо другой стимуляции (рис. 15–8). Как в устройстве автоматического пуска, эта медленная самопроизвольная активность создает фон для поиска сигналов извне, так что любые зрительные стимулы не запускают возбуждение нейронов, а лишь меняют его конфигурацию. Возбуждающие стимулы усиливают это возбуждение, а тормозные – ослабляют.
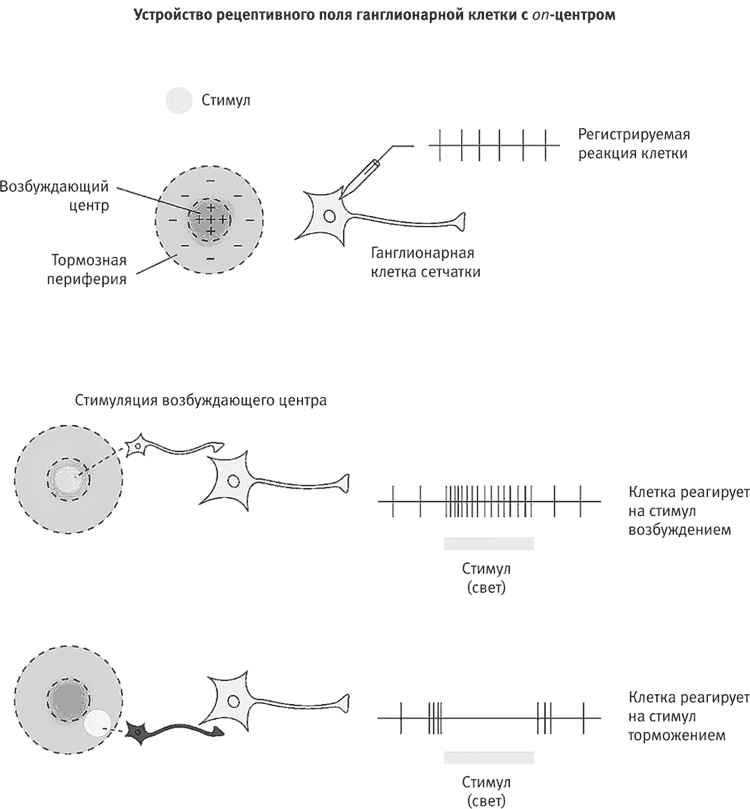
Рис. 15–8. Устройство рецептивного поля ганглионарной клетки с on-центром
Вскоре Куффлер установил, что самый эффективный способ менять конфигурацию самопроизвольного возбуждения ганглионарных клеток – не светить на всю сетчатку сильным рассеянным светом, а освещать одно пятнышко на ее поверхности. Эксперименты подтвердили, что каждый ганглионарный нейрон получает информацию от собственного участка сетчатки – рецептивного поля, на которое проецируется крошечная часть картины. Каждый из этих нейронов реагирует на стимуляцию своего рецептивного поля и передает информацию в мозг только от него. Кроме того, Куффлер обнаружил, что частота возбуждения ганглионарных нейронов зависит от силы света, падающего на рецептивное поле, а продолжительность их возбуждения зависит от продолжительности действия светового стимула. Поскольку вся сетчатка покрыта рецептивными полями ганглионарных клеток, то, на какой бы участок ни падал свет, некоторые из этих клеток будут на него реагировать. Наша зрительная система хорошо приспособлена к восприятию деталей.
Ганглионарные клетки с самыми маленькими рецептивными полями располагаются в центре сетчатки. Они получают информацию от наиболее плотно упакованных колбочек, отвечающих за самое изощренное зрительное восприятие (например, рассматривание деталей картины). Другие ганглионарные клетки, чуть в стороне от центра сетчатки, имеют несколько более широкие рецептивные поля и получают информацию уже от многих колбочек. С этих клеток начинается обработка грубых, цельных компонентов зрительного образа. Куффлер установил, что рецептивные поля ганглионарных клеток сетчатки постепенно увеличиваются в направлении периферии. Поэтому периферические клетки сетчатки не в состоянии обрабатывать информацию о незначительных деталях и дают лишь размытые изображения.
Методично обследуя сетчатку с помощью луча света, направляемого на рецептивные поля разных ганглионарных клеток, Куффлер сделал третье открытие. Он нашел, что ганглионарные клетки сетчатки бывают двух типов, поровну распределенных в ней и отличающихся характером реакции клеток на стимуляцию центральной и периферической частей рецептивного поля. Клетки с on-центром возбуждаются, когда крошечное пятнышко света падает в самый центр их рецептивного поля, но их возбуждение ослабевает, когда свет падает на его периферию. Клетки с off-центром, напротив, возбуждаются, когда свет падает на периферию рецептивного поля. Их возбуждение тормозится, когда свет падает в его центр (рис. 15–9).
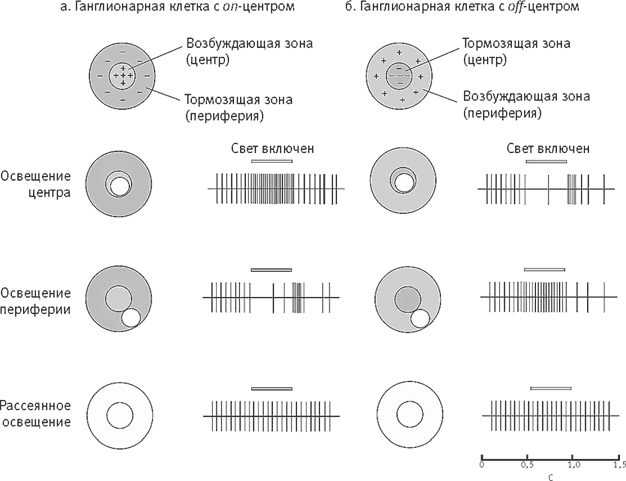
Рис. 15–9. Ганглионарные клетки сетчатки лучше всего реагируют на контрастное освещение своих рецептивных полей. Эти поля имеют округлую форму со специ-ализированными центром и периферией. Клетки с on-центром возбуждаются при стимуляции светом центра и тормозятся при стимуляции периферии, клетки с off-центром – наоборот. Здесь показаны реакции клеток обоих типов на три разных световых стимула (стимулируемый участок рецептивного поля выделен). Паттерны активности клеток в ответ на каждый из стимулов записаны с помощью внекле-точной регистрации. Продолжительность освещения показана полоской. Kuffler 1953, с изменениями. а) Клетки с on-центром лучше всего реагируют на стимуляцию светом всей цент-ральной области их рецептивного поля. На стимуляцию некоторой части этой области они реагируют не так сильно. Освещение участка периферии рецептивного поля частично или полностью подавляет возбуждение клетки, возобновляющееся вскоре после выключения света. На рассеянное освещение всего рецептивного поля клетка реагирует слабо: эффекты освещения центра и периферии компен-сируют друг друга. б) Самопроизвольное возбуждение клеток с off-центром подавляется освещением центральной области рецептивного поля, но ненадолго усиливается после выключения света. Освещение периферии рецептивного поля усиливает возбуждение таких клеток.
Открытие принципов работы ганглионарных клеток сетчатки свидетельствовало о том, что глаза млекопитающих реагируют только на те участки зрительного образа, где сила света меняется. По сути, Куффлер выяснил, что облик объекта определяется скорее контрастом между этим объектом и его фоном, чем силой света, попадающего от этого объекта на сетчатку.

Рис. 15–10. Облик видимого объекта принципиально зависит от контраста между ним и его фоном. Правое и левое серые кольца окрашены одинаково, при этом левое кажется светлее, поскольку оно расположено на более темном фоне.
Куффлер пришел к выводу, что ганглионарные клетки сетчатки реагируют не на абсолютные показатели силы света, а на контраст между светом и темнотой. Именно поэтому такие клетки нельзя эффективно стимулировать обширными световыми пятнами или рассеянным светом: рассеянный свет падает и на тормозную, и на возбуждающую области рецептивного поля каждой клетки. Кроме того, эти результаты указали на биологические основы общего принципа работы сенсорных систем мозга – принципа пренебрежения постоянными стимулами и отчетливых избирательных реакций на контрасты. Один из примеров показан на рис. 15–10. Два серых кольца окрашены одинаково, но левое кажется светлее, потому что более темный фон создает более сильный контраст. Принципы работы ганглионарных клеток сетчатки также позволяют объяснить, почему зрительная система настолько чувствительна к неравномерности падающего на сетчатку света и почему нейроны сетчатки сильнее реагируют на резкую перемену яркости изображения или освещенности. Тем самым открытия Куффлера подтвердили прогноз Гомбриха: изучение особых зрительных стимулов позволит проникнуть в тайны нейронных механизмов зрения.
Зрительная система человека развилась в ходе эволюции в соответствии с потребностями наших предков. Ранние этапы работы этой системы, изучением которых занимался Куффлер, красноречиво говорят о действии дарвинистских механизмов. Наши глаза приспособлены для обработки поступающей извне информации. Предел остроты зрения (максимальный уровень разрешения) определяется и разрешающей способностью глаз, и плотностью колбочек в центральной ямке. Колбочки передают информацию ганглионарным клеткам сетчатки. Их рецептивные поля организованы так, чтобы извлекать важнейшую информацию о зрительных образах и сводить к минимуму избыточность получаемых сведений. Принцип их организации позволяет не тратить впустую сигнальные способности всей сетчатки. Размеры центра рецептивного поля ганглионарных клеток относительно размеров его периферии также наилучшим образом приспособлены для выделения информативных элементов образа.
Куффлер продемонстрировал, что сетчатка осуществляет активное преобразование зрительной информации и ее кодирование в виде последовательностей потенциалов действия. В этих процессах задействовано огромное число фоторецепторов и других параллельно работающих нервных клеток. Вычислительная мощность этих процессов весьма велика. Получаемые на выходе из сетчатки последовательности электрических импульсов поступают в латеральное коленчатое тело таламуса, а затем в кору больших полушарий, где происходит деконструкция образов и их последующая реконструкция в виде внутренних представлений. Куффлер доказал исключительную важность контраста для сигнальной активности сетчатки, чем подготовил почву для еще более удивительных открытий.
Назад: Глава 14 Обработка мозгом зрительных образов
Дальше: Глава 16 Реконструкция видимого мира: зрение и обработка информации

