Книга: Приспособиться и выжить!
Назад: Глава 8 Создание и эволюция сложного
Дальше: Биоразнообразие — результат использования одинаковых генов разными способами
Внешность обманчива: у всех животных один и тот же набор генов для построения тела и органов
Животные, которых я увидел на Большом барьерном рифе, были представителями многих ветвей эволюционного древа. Из 35 основных групп, или типов, животных здесь встречаются кишечнополостные (кораллы, морские анемоны), губки, моллюски (кальмары, осьминоги), членистоногие (крабы), иглокожие (морские звезды и ежи), а также позвоночные (акулы, костные рыбы, морские черепахи и киты). Многие из этих животных характеризуются какими-то уникальными особенностями (панцири черепах, щупальца осьминогов, раковины моллюсков, клешни крабов и т. д.), однако у всех есть органы, необходимые для одних и тех же целей, например, глаза.
Нет сомнений в том, что глаза приносят пользу своим владельцам. Однако многих биологов со времен Дарвина удивляло разнообразие устройства глаз в царстве животных. Человек и другие позвоночные обладают глазами камерного типа с единственным хрусталиком. Крабы и другие членистоногие имеют сложные (фасеточные) глаза, в которых зрительную информацию независимым образом собирает множество отдельных зрительных ячеек. Осьминоги и кальмары, хотя и не являются нашими родственниками, тоже имеют камерные глаза, а вот у их более близких родственников, двустворчатых моллюсков, глаза бывают трех типов: камерные глаза с единственным хрусталиком, зеркальные глаза с хрусталиком и отражателем, а также сложные глаза, составленные из 10–80 ячеек.
На протяжении 100 с лишним лет ученые считали, что большое разнообразие строения глаз является результатом независимых «изобретений», произошедших в разных группах. На основании клеточного строения глаз животных знаменитый биолог-эволюционист Эрнст Майр и его коллега Л. В. Сальвини-Плевен предположили, что глаза в ходе эволюции возникали независимым образом от 40 до 65 раз.
С одной стороны, это утверждение поддерживает теорию о воспроизведении эволюционных событий при возникновении одних и тех же потребностей (в данном случае речь идет о потребности видеть). Идея о повторении эволюции глаз была широко распространена. Однако новые открытия заставили ученых пересмотреть свой взгляд на эволюцию глаза. Основной вопрос заключается в следующем: возникли ли глаза «из ничего» или их эволюция строилась на готовых элементах, имевшихся у одного или нескольких общих предшественников. Именно от этого зависят наши представления о вероятности эволюции сложных структур. Конечно, кажется более «трудным делом» (менее частым или менее вероятным) создание какой-либо структуры на пустом месте, из ничего, по сравнению с ситуацией, когда части этой структуры уже существуют. Новые данные показывают, что совершенно разные глаза, имеющиеся у разных типов животных, имеют между собой гораздо больше общего, чем кажется на первый взгляд, и эта общность позволяет нам лучше понять процесс эволюции сложных структур.
История формирования нового взгляда на эволюцию глаза началась в 1994 г. Вальтер Геринг и его коллеги из Университета Базеля (Швейцария) занимались изучением гена, необходимого для развития сложного глаза у дрозофилы. Когда этот ген инактивировали с помощью мутаций, глаз не формировался. Еще раньше ученые, занимающиеся генетикой дрозофил, прозвали этот ген безглазым (eyeless) (многие гены получают свое название от той функции, которая нарушается в случае их мутации; на самом деле нормальная функция данного гена состоит в содействии формированию глаза). Когда ученые выделили ген eyeless, они, к своему большому удивлению, обнаружили, что он кодирует белок, который чрезвычайно сильно напоминает белки, кодируемые мышиным и человеческим генами. Мышиный белок назвали маленьким глазом (Small eye); он также необходим для формирования глаза. Человеческий белок получил имя аниридия (Aniridia), поскольку его дефект приводит к исчезновению радужной оболочки глаза. Сходство между белками человека, мыши и дрозофилы настолько велико, что становится ясно — это один и тот же белок у разных видов организмов (рис. 8.2).

Рис. 8.2. Фрагмент белковой последовательности, кодируемой геном Pax-6. Здесь представлены фрагменты белка дрозофилы, мыши и человека. Обратите внимание на большое сходство между белками дрозофилы и млекопитающих, а также на идентичность последовательностей белка мыши и человека.
Теперь этот белок носит общее и менее выразительное имя — Pax-6.
Открытие гена Pax-6 тут же вызвало новый вопрос: является ли наличие одинаковых генов у животных со столь разными глазами, как у насекомых и млекопитающих, простым совпадением или имеет какой-то глубокий смысл? Другими словами, использовали ли дрозофилы и млекопитающие ген Pax-6 для независимой эволюции своих глаз «из ничего» или кажущиеся столь разными глаза имеют между собой больше общего, чем представляется на первый взгляд, и их формирование при участии гена Pax-6 является отражением какого-то фундаментального принципа?
Теперь появилось множество новых данных, позволяющих ответить на этот вопрос. Сначала экспериментальным путем было показано, что гены Pax-6 мыши и дрозофилы являются взаимозаменяемыми. Швейцарские ученые активировали ген Pax-6 дрозофилы в необычных местах, таких как ноги, крылья или усики, и обнаружили, что это приводит к формированию тканей глаза! Затем они установили, что мышиный ген Pax-6 может индуцировать образование глазной ткани у дрозофил. Таким образом, эти гены имеют одинаковые функции, а не только очень похожие последовательности. Вспомните, в третьей главе мы говорили о том, что никакой ген не может сохраняться во времени без поддержки со стороны естественного отбора. По какой-то причине функция и последовательность белка Pax-6 сохранялась на протяжении длительного периода эволюции животных — более 500 млн лет.
Причина сохранности гена Pax-6 стала ясна в результате серии экспериментов, посвященных исследованию роли этого гена в образовании глаз у других животных. Изучение гена Pax-6 кальмаров и различных червей, таких как планарии и ленточные черви, показало, что ген также участвует в формировании сложных или простых глаз у этих животных.
Поскольку Pax-6 задействован в развитии глаз у столь широкого круга организмов, очень маловероятно, что все они стали использовать этот ген случайно. Участие гена Pax-6 в развитии глаза должно иметь исторические причины. Это означает, что общий предшественник всех этих животных использовал Pax-6 для создания каких-то, возможно очень примитивных, глаз. И все удивительные и сложные глаза, развившиеся у потомков этого общего предка, эволюционировали именно на этом основании.
Следующий вопрос, на который необходимо ответить, если мы хотим воссоздать картину эволюции сложных органов, заключается в том, что же это было за основание. Какие элементы, в дальнейшем использовавшиеся для эволюции более сложных глаз, уже существовали у общего предка животных?
Об этих элементах нам известно достаточно много. Глаза любого типа состоят из регистрирующих свет клеток, называемых фоторецепторными клетками, и пигментных клеток, определяющих угол падения света на фоторецепторные клетки. Таким образом, резонно предположить, что самые примитивные глаза состояли из этих двух типов клеток. Именно такое предположение и сделал Дарвин: «Самый простой орган, который можно было бы назвать глазом, состоит из зрительного нерва, окруженного пигментными клетками и покрытого прозрачной кожей, но без хрусталика или преломляющего тела».
Такие простые двухклеточные глаза действительно существуют. Они были обнаружены у личинок некоторых существ, таких как морские черви нереиды (Platynereis dumerilii). На второй день развития оплодотворенной яйцеклетки личинка имеет пару двухклеточных глаз, «пристально глядящих» с передней стороны туловища (рис. 8.3, верхний ряд).
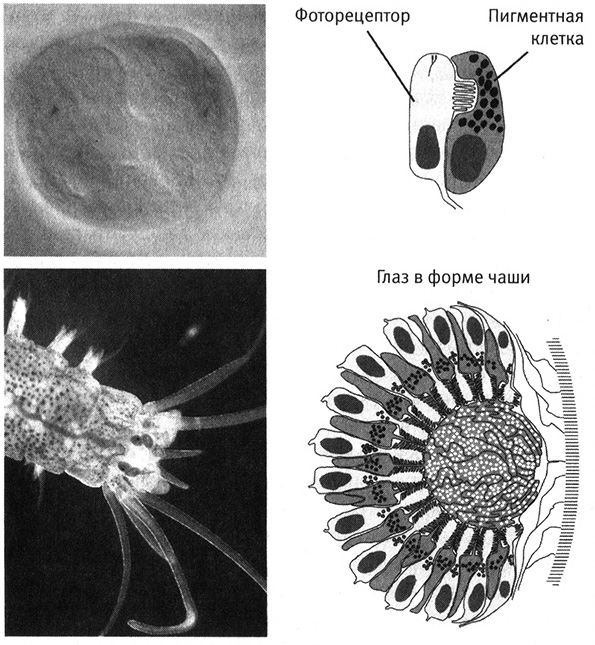
Рис. 8.3. Простые и более сложные глаза у морских червей. На второй день развития у личинки нереиды (вверху слева) уже существует пара простых глаз, каждый из которых состоит всего из двух клеток (вверху справа). У взрослого червя образуются две пары глаз (внизу слева), состоящих из гораздо большего числа клеток, организованных в форме чаши (внизу справа). В формировании обоих типов глаз задействованы одинаковые гены. Верхние рисунки, а также нижний правый рисунок любезно предоставлены Детлевом Арендтом из Европейской молекулярно-биологической лаборатории в Гейдельберге (из статьи Arendt et al., 2002, Development 129:1143, с изм.); нижний левый рисунок предоставлен Бенжамином Прюдоммом из Медицинского института Говарда Хьюза и Университета Висконсина.
Но простота строения этих глаз обманчива. Они построены из тех же компонентов, что и более сложные и совершенные глаза. Например, регистрация света фоторецепторными клетками этих простых глаз основана на действии опсинов — тех самых зрительных пигментов, о которых мы говорили в предыдущих главах. Все животные используют опсины для регистрации света. Объяснить этот факт можно единственным образом: опсин существовал уже в примитивных глазах общего предшественника большинства животных и с тех пор используется для детекции света во всех типах глаз.
Картину формирования и эволюции более сложных глаз исследовали на примере тех же личинок нереиды. Рядом с двухклеточными глазами личинки в какой-то момент начинают формироваться более крупные глаза взрослой особи, имеющие форму чаши и состоящие из гораздо большего числа фоторецепторных и пигментных клеток (рис. 8.3, нижний ряд). Сложность в данном случае является результатом организации большего количества таких же клеток в трехмерном пространстве — тот же строительный материал, другая конструкция. А для строительства используются те же инструменты. В этом процессе участвует ген Pax-6 и еще как минимум два других гена, таких же как у дрозофил и позвоночных. Создание более крупного, но все еще примитивного глаза взрослого червя из тех же основных типов клеток, а также использование тех же генов, что служат для построения более сложных фасеточных и камерных глаз, демонстрирует нам путь создания и эволюции сложных органов. Анализируя этот процесс, мы видим, что сложные органы строятся путем сборки большого количества клеток всего нескольких типов и что в ходе эволюции для построения глаз современных животных продолжали использоваться те же типы клеток и те же гены. Для создания глаз с различным строением у разных животных применяются те же «строительные кирпичики» и те же «инструменты».
С учетом этих новых представлений получается, что разные типы глаз являются продуктом разных эволюционных путей, начавшихся со сходных стартовых позиций, с некоей простой конфигурацией фоторецепторных и пигментных клеток, но не с пустого места. Также неверно, что камерный глаз эволюционировал из фасеточного глаза или наоборот. Представьте себе современные сложные типы глаз и попытайтесь превратить один тип в другой. Это невозможно сделать без потери эффективности органа у промежуточных вариантов. Но в ходе эволюции этого и не происходило.
Напротив, сегодня нам представляется, что история эволюции глаза состояла в повторяющемся акте построения более сложного органа из более простых «протоглаз» (рис. 8.4).
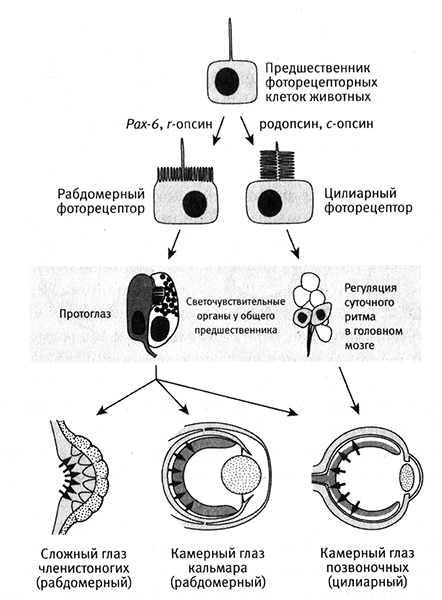
Рис. 8.4. Происхождение и эволюция сложного глаза. Под контролем гена Pax-6 у общих предшественников животных возникли фоторецепторные клетки, способные детектировать свет с помощью белков-опсинов. Сложные глаза эволюционировали из более простой структуры, состоявшей из фоторецепторных и пигментных клеток. Общий предшественник билатеральных животных имел два типа фоторецепторных клеток: рабдомерные фоторецепторы в протоглазах отвечали за зрение, а цилиарные фоторецепторы в головном мозге — за регуляцию суточного ритма. Рабдомерные рецепторы стали основой эволюции глаз членистоногих и головоногих животных, а в эволюции глаз позвоночных животных были задействованы оба типа рецепторов. Рисунок Лианн Олдс.
Роль естественного отбора в эволюции сложных глаз с лучшими оптическими характеристиками объяснить несложно. Начиная с простой структуры, постепенно накапливались небольшие изменения, улучшавшие функциональные характеристики органа. Если мы рассмотрим лишь одну ветвь на филогенетическом древе животных, например ветвь моллюсков, то обнаружим большое разнообразие глаз с разной степенью сложности (рис. 8.5).
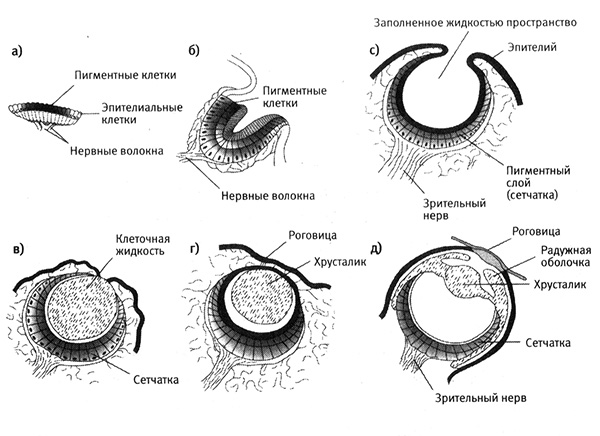
Рис. 8.5. Различные стадии эволюции глаза у моллюсков. Моллюски имеют глаза различного строения; это могут быть простые пигментированные глазки (а), глаза в форме чаши (б), глаза, заполненные клеточной жидкостью (в), глаза с наружной линзой (г) и сложные глаза (как у кальмара) (д). Из книги M. W. Strickberger, Evolution, © 1990 by Jones and Bartlett Publishers, Boston.
С помощью компьютерного моделирования Дан Нильсон и Сюзанна Пелгер из Университета Лунда (Швеция) показали, что отбор малых вариаций за 2 тыс. стадий на протяжении 500 тыс. лет мог привести к образованию камерного глаза из простого прототипа.
Детализация картины эволюции глаза помогла объяснить некоторые интересные различия между разными типами глаз. Например, в глазу человека фоторецепторы повернуты от света и находятся на задней стенке глазного яблока, тогда как у кальмара они направлены к свету и располагаются спереди (см. рис. 8.4).
Чрезвычайно сложно (и не нужно) пытаться представить себе, каким образом один тип строения перешел в другой. Очевидно, что для эволюции глаз камерного типа существовало несколько путей, и головоногие и позвоночные пошли разными путями.
Еще одним важным отличием глаз человека от глаз кальмара или дрозофилы является тип фоторецепторов: у человека и других позвоночных фоторецепторные клетки (палочки и колбочки) относятся к так называемому цилиарному типу, тогда как фоторецепторы кальмаров и дрозофил относятся к рабдомерному типу. Различие состоит в том, каким образом мембраны рецепторных клеток каждого типа расширяются для соединения с опсинами. Этот признак служил важным аргументом в пользу независимого происхождения глаз позвоночных и других животных.
Новые открытия, сделанные все на тех же незамысловатых нереидах, прояснили тайну происхождения наших глаз и фоторецепторов. Детлев Арендт и его коллеги из Европейской молекулярно-биологической лаборатории в Гейдельберге (Германия) обнаружили в развивающемся головном мозге нереид несколько цилиарных (реснитчатых) клеток, которые странным образом напоминали фоторецепторные клетки позвоночных. Дальнейшие исследования показали, что эти клетки синтезируют некий опсин, который больше похож на опсины позвоночных, чем на опсины, обнаруженные в фоторецепторных клетках в глазах самих нереид или других беспозвоночных животных. Опсин цилиарных клеток головного мозга (c-опсин), как выяснилось, нужен не для зрения, а для регуляции суточного ритма. Таким образом, оказалось, что кольчатые черви нереиды имеют оба типа фоторецепторов и опсинов. Это означает, что общий предшественник нереид, кальмаров и позвоночных также имел все эти типы фоторецепторов и опсинов. Рабдомерные фоторецепторы и их опсин (r-опсин) послужили для создания зрительной системы у членистоногих и головоногих организмов, тогда как для создания зрительной системы у позвоночных природа использовала цилиарные фоторецепторы и c-опсин. Кроме того, рабдомерные рецепторные клетки в глазах позвоночных превратились в так называемые ганглиозные клетки сетчатки, функция которых состоит в передаче сигнала в головной мозг. Таким образом, глаза позвоночных животных, по-видимому, сформировались из обоих типов фоторецепторных клеток.
Глаз, таким образом, не только перестал быть примером органа, которому трудно дать эволюционное объяснение, но и служит для нас одним из важнейших источников информации о том, как эволюция создает сложные структуры с помощью общих генетических инструментов. Открытия в области биологии развития показали, что для создания совершенно разных по строению сердец, пищеварительных систем, мышц, нервов и конечностей у всех видов животных используются одни и те же генетические инструменты. Совершенно очевидно, что подобно фоторецепторам, которые представляют собой древний тип клеток, другие типы клеток, из которых состоят многие ткани и органы, также имеют древнее происхождение. Более того, в результате анализа генов и геномов стало известно, что большинство животных снабжены сходными наборами инструментов (генов) для построения тела и органов (ветвь позвоночных имеет более широкий ассортимент этих генов в результате масштабных процессов удвоения генов). Все это говорит о том, что сам этот набор инструментов также имеет древнее происхождение и должен был существовать у общего предшественника еще до начала эволюции тел и органов большинства современных животных.
Мы не знаем, кто был этим общим предком. Но если попытаться изобразить это существо, то нужно представить себе небольшое свободно плавающее морское животное с мягким телом, напоминающее личинку нереиды (см. рис. 8.3, слева вверху), с полным генетическим «набором инструментов развития», множеством типов клеток и простыми органами. Такое животное является хорошей аппроксимацией того существа, которое послужило основой эволюции всего царства животных.
Эти новые открытия позволяют нам проследить за процессом возникновения и эволюции сложных структур. Но при этом возникает новый парадокс: почему при такой общности клеточных типов и генов возникло такое большое разнообразие форм?
Назад: Глава 8 Создание и эволюция сложного
Дальше: Биоразнообразие — результат использования одинаковых генов разными способами

