В глубоком синем море
На большие морские глубины солнечный свет проникает сквозь толщу воды, поэтому там темно. За зрение в слабом рассеянном свете отвечают фоторецепторы палочек, а опсин этих клеток называется родопсином. Родопсин человека и большинства наземных млекопитающих настроен таким образом, чтобы максимально поглощать свет с длиной волны около 500 нм.
На глубину около 200 м проникает лишь свет из узкого синего диапазона с длиной волны около 480 нм. Интересно, что родопсины глубоководных рыб и дельфинов сдвинуты в синюю область, то есть их максимум поглощения на 10–20 нм ниже максимума поглощения наземных млекопитающих. Ученые активно изучали тонкую настройку родопсина таких океанских жителей, как афалина, обыкновенный дельфин, черный дельфин, атлантический ремнезуб, солнечник и различные угри. Чтобы установить влияние отдельных аминокислотных остатков на различия между родопсинами разных видов животных, одни аминокислоты или их группы заменяли другими, обнаруженными у других видов. В частности, у афалины за сдвиг максимума поглощения родопсина на 10 нм в синюю область по сравнению с родопсинами наземных млекопитающих отвечают аминокислоты в положениях 83, 292 и 299 (рис. 4.6). Максимум поглощения родопсина ремнезуба еще сильнее сдвинут в синюю область (до 484 нм), и этот родопсин отличается от родопсина дельфина только аминокислотой в положении 299.
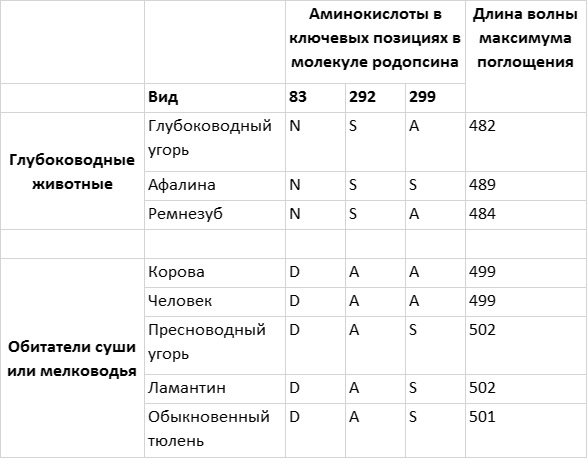
Рис. 4.6. Настройка родопсина зависит от глубины, на которой обитают морские животные. Глубоководные рыбы и китообразные имеют родопсин, максимум поглощения которого сдвинут в синюю область спектра по сравнению с родопсинами животных, обитающих на суше или на мелководье. У животных, обитающих на одинаковой глубине, часто в ключевых позициях в последовательности родопсина стоят одни и те же аминокислоты. Рисунок Джейми Кэрролл.
Связь «синего сдвига» родопсина с адаптацией к жизни в морских глубинах подкрепляется данными анализа родопсинов глубоководных и пресноводных угрей. Родопсин угря, обитающего на большой глубине, характеризуется сдвигом максимума поглощения в синюю область и в ключевых положениях имеет точно те же аминокислотные остатки, что и родопсин ремнезуба. Родопсин пресноводного угря имеет максимум поглощения, близкий к таковому у наземных млекопитающих, а в ключевых позициях в его последовательности находятся те же аминокислотные остатки, что и у ламантина и тюленя — двух обитающих на мелководье млекопитающих с типичным вариантом родопсина.
Эта поразительная связь между «синим сдвигом» родопсина и жизнью на большой глубине полна глубокого смысла. Резонно предположить, что естественный отбор способствовал настройке родопсина в соответствии со средой обитания. Однако существует еще более важная причина подозревать влияние естественного отбора, и она становится ясна при рассмотрении эволюционных связей между видами животных, перечисленных на рис. 4.6. Дельфины и ремнезубы относятся к отряду китообразных — млекопитающих, которые произошли от сухопутного предка, вернувшегося в водную стихию. Это может показаться удивительным, но ближайшими современными родственниками китообразных являются гиппопотамы, олени, коровы, свиньи и верблюды. Это следует из результатов анализа последовательностей SINE и LINE, а также других последовательностей ДНК. Поскольку родопсины наземных родственников китообразных настроены на длину волны 500 нм, можно с полным основанием заключить, что родопсин дельфинов и китов изменился после того, как их эволюционная ветвь отделилась от ветви других млекопитающих.
Однако угри — это рыбы, и их эволюционная ветвь отделилась от ветвей других позвоночных несколько сотен миллионов лет назад. Это означает, что одинаковые по своей сути различия между глубоководными и пресноводными угрями и морскими и наземными млекопитающими возникли независимо. Если в одинаковых позициях в каком-то белке у двух или нескольких видов животных при адаптации к определенным условиям появляются одни и те же аминокислотные остатки (как у китообразных и глубоководного угря в нашем примере), это говорит о работе естественного отбора. Замечу, что пример эволюции родопсина — лишь один из множества случаев, когда эволюция воспроизводит саму себя (мы подробнее поговорим об этом в шестой главе).
Теперь перейдем от красного, зеленого и синего к невидимым для нас цветам — и к целому миру взаимоотношений между животными, связанных с ультрафиолетовым зрением.
Назад: Красное и зеленое
Дальше: За пределами радуги

