Лотерея мутаций: все мы мутанты
Источник любых изменений — мутация. Слово это приобрело множество значений, что стало причиной двух широко распространенных заблуждений, которые я сразу же хочу развеять. Первое заблуждение заключается в том, что все мутации вредны и, следовательно, не могут быть созидательными, лишь разрушительными. Это совершенно неверно, как показывает пример ледяной рыбы. Мы увидим, что вероятность появления полезной мутации гораздо выше, чем вероятность покупки выигрышного лотерейного билета. Второе заблуждение заключается в том, что если мутации возникают случайно (а это так и есть), то они не могут вносить вклад в создание сложных и упорядоченных систем в живой природе. Это заблуждение связано с неумением различать мутацию и отбор. Возникновение мутаций — слепой процесс, а естественный отбор таковым не является. Мутации приводят к появлению случайных вариаций, а отбор разделяет выигравших и проигравших. Кроме того, естественный отбор — это кумулятивный процесс. Рим и его жители возникли не за один день, и не за мгновение замерзли антарктические моря и сформировались ледяные рыбы. Эволюция создавала ледяную рыбу, человека и другие виды животных на протяжении сотен тысяч и миллионов поколений. В уже функционировавших существах появлялись новые мутации. Они не могли и не должны были приводить к мгновенному появлению сложных структур или функций.
Чтобы оценить созидательную способность мутаций, мы должны знать, какие бывают мутации и с какой частотой они происходят. За полвека исследований у нас сложилась четкая картина динамических изменений в ДНК. Я коротко остановлюсь на том, каким образом мутации могут изменять последовательность ДНК.
Для собственного воспроизводства любой организм должен уметь копировать свою ДНК. Создание копии ДНК — сложный биохимический процесс. В ходе этого процесса возникают ошибки, и если они не исправляются быстро и должным образом, рождаются мутации. Существует великое множество видов мутаций. Если рассматривать ДНК как текст, то разные типы мутаций соответствуют разным типам ошибок при переписывании. ДНК любого вида представляет собой от миллионов до миллиардов комбинаций букв A, C, G и T. Самая распространенная ошибка (опечатка) заключается в замене правильной буквы на неправильную. Но могут происходить и другие события, такие как инсерции (включения) или потери (делеции) отдельных букв и групп букв. Также возможны ошибки копирования и вставки: так возникают удвоения (дупликации) фрагментов текста. С разной частотой могут возникать повторы нескольких букв, целых генов или даже групп генов (удвоение генов может расширять информационное содержание ДНК, и в четвертой главе мы увидим, что дупликация ДНК является важным источником новых признаков). Участки последовательностей ДНК также подвергаются изменениям в результате инверсий и перестановок фрагментов. Таким образом, в каждом новом организме имеются новые мутации.
Ученые изучали частоту возникновения мутаций у многих видов организмов. На семь миллиардов оснований ДНК в организме каждого человека приходится примерно 175 новых мутаций. Как сказал биолог Арман Леруа, «мы все мутанты».
«Подождите! Как же это возможно? — воскликнете вы. — Разве мутации не вредны?» Да, некоторые вредны, но не все. Мы в большинстве случаев не страдаем от этих 175 мутаций по той причине, что они: 1) происходят в тех участках ДНК, где нет значимой информации; 2) возникают внутри гена или рядом с ним, но не влияют на его работу; 3) компенсируются наличием второй копии гена, которая обычно имеется у каждого индивида, или 4) воздействуют на ген таким образом, что его функция изменяется лишь в допустимых пределах. Именно вариации размера, формы, цвета и других физических и химических признаков делают каждого из нас уникальным существом. Это и есть сырьевой материал для работы эволюции.
Я приведу конкретный пример, который иллюстрирует высокую вероятность возникновения полезных для адаптации мутаций (в последующих главах я приведу множество других примеров адаптационных изменений). Представим себе популяцию диких мышей, все представители которой имели светлую окраску и жили на песчаной почве. И вот через столетия или тысячелетия геологическая активность в местах обитания этих мышей привела к извержению вулкана и выбросу лавы. Остывшая лава образовала на поверхности земли черные каменистые выступы. Окраска мышей перестала соответствовать цвету почвы: на темном фоне их светлая шкурка стала хорошо видна хищникам, таким как филины и совы. Темные мыши были бы в этих условиях более защищенными. Итак, мы хотим узнать:
• сколько времени потребуется для появления в популяции светлых мышей мутаций, обеспечивающих темную окраску;
• как быстро подобные мутации распространятся в популяции.
Ответ на первый вопрос зависит от комбинации вероятности и времени. Для решения следует прибегнуть к тем же расчетам, которыми пользуются для определения вероятности выигрыша в лотерею. Ответ на второй вопрос зависит от связи между отбором и временем. И об этой математике мы уже говорили.
Скорость появления мутаций у мышей достаточно хорошо изучена. Исследователи из лаборатории Джексона в Бар-Харборе, штат Мэн, занимались скрещиванием мышей на протяжении нескольких десятилетий и располагают информацией о частоте спонтанных мутаций, основанной на изучении миллионов мышей. Если говорить о мутациях отдельных оснований ДНК, то они у каждой мыши происходят примерно в двух позициях на каждый миллиард оснований (всего в геноме мыши этих оснований около 5 млрд). В гене среднего размера существует около 1 тыс. позиций, в которых может произойти мутация. Учитывая наличие 1 тыс. позиций для мутации в каждом гене и возможность возникновения двух мутаций в каждом миллиарде позиций, получаем, что мутация в конкретном гене имеет место у одной из 500 тыс. особей. Таким образом, процесс копирования ДНК является достаточно точным, но не безупречным.
Кроме того, нам известно, что мутации многих генов могут привести к изменению окраски животного. Я расскажу о роли всего одного гена, мутантные варианты которого были получены в лаборатории Джексона и о котором мы знаем достаточно много. Многие мутации этого гена, названного MC1R, приводят к тому, что мышь приобретает черную окраску, если у нее есть хотя бы одна копия мутантного гена.
Чтобы рассчитать частоту появления мутаций, отвечающих за темную окраску мышей, я буду исходить из следующих допущений.
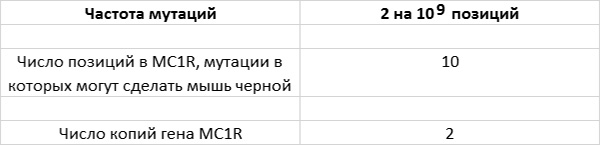
Теперь перемножим эти значения: 10 позиций в гене MC1R умножим на 2 гена у каждой мыши и на 2 мутации на каждый миллиард позиций и получим 40 мутантов на каждый миллиард мышей. Таким образом, каждая мышь имеет один шанс из 25 млн получить мутацию гена MC1R, способствующую появлению темной окраски.
Может показаться, что это очень маленький шанс, но мы пока не учли численность популяции и скорость воспроизводства мышей. Частота появления мутации зависит от размера популяции и уровня рождаемости. Мышей много; в каждом поколении появляется на свет множество детенышей. В той популяции мышей, о которой я рассказываю, насчитывается от 10 тыс. до 100 тыс. особей. Чтобы оценить частоту появления черных мутантов, нужно учесть еще и плодовитость мышей. Самки этого вида мышей приносят от двух до пяти детенышей два или три раза в год. Допустим, каждая самка приносит пять детенышей в год. Если умножить число самок на число детенышей каждой самки, мы получим количество новорожденных мышат в год в данной популяции. Предположим, в популяции 10 тыс. мышей; половину из них составляют самки; каждая приносит по пять мышат. Таким образом, в этой популяции ежегодно появляется 25 тыс. мышат. Теперь умножим это число на 1/25 000 000 (вероятность мутации у одной особи) и получим, что одна черная мышь появляется один раз в 1 тыс. лет. Таким образом, за миллион лет мутации, приводящие к появлению черной окраски, могут независимо возникнуть 1 тыс. раз. Итак, в нашей популяции из 10 тыс. особей черная мышь появляется один раз в 1 тыс. лет. Если численность популяции составляет 100 тыс. мышей, черные особи появляются чаще — один раз в 100 лет. Для сравнения, если вы покупаете 10 тыс. билетов лотереи «Пауэрболл» в год, то будете выигрывать каждые 7500 лет.
Теперь поговорим о скорости распространения черной окраски в популяции. После того как мутация появилась, ее распространение зависит от скорости отбора и от времени. Ключевую роль в этом процессе играет селективное преимущество черного меха и эффективный размер популяции (это не общее число особей, а количество участвующих в размножении особей с учетом дополнительных факторов; этот параметр обозначают как Ne). Чем больше преимущество, тем быстрее распространяется признак, но чем больше размер популяции, тем больше времени требуется для того, чтобы все мыши стали черными. Для расчета времени (f, измеряется в поколениях), за которое все мыши станут черными, можно воспользоваться формулой:
t = (2/s) In (2Ne).
Ниже представлено несколько значений t при разных значениях преимущества s в популяции из 10 тыс. особей. Обратите внимание, что при повышении коэффициента отбора время, необходимое для распространения благоприятной мутации, сокращается.
s = 0,001, t = 19 807 поколенийs = 0,01, t = 1981 поколениеs = 0,05, t = 396 поколенийs = 0,1, t = 198 поколенийs = 0,2, t = 99 поколений
Оценочное значение коэффициента отбора темных мышей, живущих на поверхности лавы, превышает 0,01. Как следует из таблички, в этом случае мутация, ответственная за темную окраску меха, распространится по всей популяции менее чем за 2 тыс. поколений, или менее чем за 2 тыс. лет. Таким образом, даже мутация, приносящая небольшое преимущество, распространяется в популяции за весьма короткий отрезок времени по геологической шкале.
Чтобы точнее оценить вероятность событий, происходящих в природе, необходимо учесть еще несколько факторов. Одним из этих факторов является вероятность потери полезной мутации до того, как она начнет распространяться. Мыши или их потомки могут не дожить до половой зрелости или не передать мутацию своим потомкам. Вероятность потери мутации также зависит от комбинации случайности и отбора. Я не стану выводить формулу, только скажу, что вероятность успешного распространения мутации в популяции приблизительно вдвое превышает коэффициент отбора. В нашем случае, когда мутация обеспечивает небольшое преимущество с s = 0,01, вероятность распространения мутации составляет 2 %. Если бы коэффициент s был равен 0,05, вероятность распространения мутации составляла бы 10 %. Таким образом, если за 1 млн лет мутация может появиться 1 тыс. раз, за это время она имеет возможность распространиться от 20 до 100 раз.
Кроме того, в приведенных выше расчетах я не учитывал миграцию животных. На самом деле животные не сидят на месте — в частности, мыши могут перемещаться из областей со светлым песком в области с темной почвой и наоборот. При этом на светлой почве имеют преимущество светлые мыши, а на темной — темные. Это усложняет описание реальной картины, но не влияет на вывод о том, что эволюция может идти очень быстро.
Важно учитывать, что расчет времени появления нового варианта окраски был основан на том, что вся популяция мышей имела окраску, адаптированную к обитанию на песчаной почве. В реальности в любой популяции мышей, голубей, людей или других видов организмов обычно существует множество вариантов признака. В качестве примера на рис. С представлено 18 вариантов окраски одного вида подвязочной змеи.

Я привожу пример с окраской мышей по той причине, что он иллюстрирует временные рамки, в пределах которых мутации и отбор в типичной популяции животных приводят к эволюционным изменениям. Но еще этот пример хорош тем, что он отражает реальную ситуацию. В пустыне Пинакате в Аризоне есть участки земли, покрытые черной лавой в результате вулканических извержений, произошедших миллионы лет назад. Здесь живут мыши двух цветов — черные и песочные. Майкл Нахман из Университета Аризоны и его коллеги показали, что мыши песочного цвета чаще встречаются на песчаной почве, а черные мыши — на темных лавовых полях (рис. 2.6).

Рис. 2.6. Окраска диких мышей. Темные мыши (верхний рисунок) чаще встречаются на черной каменистой земле, тогда как светлые мыши (нижний рисунок) обитают на песчаной почве. Из статьи S. B. Benson, 1933, University of California Publications in Zoology 40:1.
Ученые также установили генетическую основу различия между темными и светлыми мышами (подробнее мы поговорим об этом в шестой главе). Этот пример хорош тем, что в нем реальные экологические и генетические данные демонстрируют действие естественного отбора в реальных временных рамках. Вывод из этого и многих других подобных примеров заключается в том, что мутации могут быть «созидательными» и что эволюцию ограничивает не потенциальная возможность мутаций, а экологическая необходимость.
Назад: Естественный отбор в дикой природе
Дальше: Время

