Книга: Изгнание из Эдема
Назад: ГЛАВА ТРЕТЬЯ ДВА ТИПА ЕВРОПЕЙЦЕВ
Дальше: ГЛАВА ПЯТАЯ РАННИЕ АЗИАТСКИЕ ВЕТВИ
ГЛАВА ЧЕТВЕРТАЯ
ПЕРВЫЕ ШАГИ В АЗИЮ, ПЕРВЫЙ ПРЫЖОК В АВСТРАЛИЮ

Фото. Едва различимый, выполненный блекло-пурпурным цветом контур слева от колпачка объектива — древнейший датируемый наскальный рисунок фигуры человека. Его возраст — 17 тыс. лег. Окрестности Кимберли, Австралия.
В главе 1 мы уже знакомились с доказательствами того, что в древности исход из Африки совершила одна генетическая линия, L3, две дочерних линии которой, Манью и Насрин, совместными усилиями заселили все остальные просторы земного шара. Эта единичная генетическая линия имеет ключевое значение для обоснования гипотезы об одном-единственном исходе, совершенном по южному маршруту. Как мы видели в главе 3, тот удивительный факт, что европейцы являются потомками только одного из этих кланов — клана Насрин, позволяет определить локус возникновения этой ветви, появившейся в одной из колоний мигрантов в самом начале пути из Африки, по всей вероятности — где-то в районе Персидского залива. Полное отсутствие каких-либо следов дочерних генетических ветвей Манью и Насрин в Северной Африке и на Леванте и обилие их в Индии исключает возможность северного маршрута миграции в Европу и служит неопровержимым аргументом в пользу версии о южном маршруте исхода из Африки, пролегавшем через устье Красного моря. А теперь давайте обратимся к более детальному рассмотрению генетических и иных свидетельств самой ранней волны колонизации Южной Азии и побережья Индийского океана. С этого момента история Y-хромосомы становится более полной, и в ней появляются некоторые другие генетические маркеры, помимо генов Адама и Евы. К Южной Азии относятся все те страны, которые встретились мигрантам-первопроходцам, кормившимся собирательством на прибрежной полосе, на их пути вдоль северного побережья Индийского океана.
Выжившие в долгом пути: место и время
Если у всех людей, живущих ныне за пределами Африки, был один общий предок, то дата исхода предков аборигенов Австралии будет абсолютно тождественной времени исхода предков европейцев, индусов и китайцев. Более того, все эти многообразные линии должны сходиться в некой общей точке в пространстве и времени, и все поселения, основанные мигрантами на путях скитаний, должны содержать генетические и даже материально-культурные следы, позволяющие определить, кто их оставил. Так, проанализировав пробы мтДНК или Y-хромосомы у любого жителя нашей планеты за пределами Африки, мы можем отыскать их место на соответствующей ветви генетического древа выходцев из Африки. И то, какое место они занимают на ветви древа выходцев из Африки, часто может показать нам, как они попали туда, а порой даже сказать, когда это произошло.
Нам не следует поддаваться соблазну делать поспешные выводы, основанные на изолированных генетических данных. Истинная структура ветвей и их географическое распределение часто являются куда более красноречивыми, чем данные по ним, полученные при помощи молекулярных часов. Эти данные носят весьма приближенный характер, и поэтому необходимо брать их средневзвешенные значения с учетом других источников информации, а именно — эволюции климатических факторов, которые показывают, когда конкретно могли сложиться благоприятные для миграции условия. Еще одним источником сведений служат археологические свидетельства, разумеется, если таковые имеются.
Археологические свидетельства о миграции по прибрежной полосе неизбежно являются крайне приблизительными. В период между 60 и 80 тысячами лет тому назад колебания уровня Мирового океана составляли от 50 до 80 м ниже современного уровня, так что древние береговые полосы и даже обширные материковые земли, по которым бродили наши далекие предки, сегодня находятся глубоко под водой. Однако существует целый ряд интереснейших исключений из этих эффектов затопления.
На некоторых участках побережья Индийского океана нам до сих пор встречаются небольшие поселения так называемых туземных этнических групп, которые вполне могут оказаться прямыми потомками тех самых бродячих собирателей. Еще задолго до появления методов изучения мтДНК главным основанием для выделения этих туземных групп был тот факт, что их культура и внешний облик резко отличались от основной массы населения в прилегающих районах. Некоторые из этих туземных групп имели черты, общие с африканцами, например, курчавые волосы и очень темную кожу. В этой и следующей главах я намерен рассмотреть более объективные генетические и физические свидетельства в пользу этой гипотезы, но для начала приведу названия некоторых из этих групп.
Представителей таких туземных групп часто обозначают такими противоречивыми и предвзятыми терминами, как «австралоиды», «негроиды» и «негритосы», чтобы подчеркнуть их отличие от жителей прилегающих районов. К числу таких групп — начиная с побережья Южной Аравии — относится племя хадрамаут, представителей которого часто описывают как австралоидов, но которые, однако, отличаются куда более выраженной примесью африканской крови. На побережье Пакистана, в районе устья Инда, найдены генетические и исторические свидетельства в пользу гипотезы о заметной — и к тому же очень поздней — примеси африканской крови в жилах местных жителей, что можно считать результатом работорговли и завоза в эти места рабов из Африки. Иначе обстоит дело с другими, так называемыми негроидными, туземными группами, сохранившимися в Индии, в частности — народностями кадар и паньян. В той же Южной Азии существует целый ряд других туземных групп, включая так называемые прото-австралоидные этнические группы, такие, как корава, йанади, ирула, гадаба и ченьчу в Индии, а также ведды в Шри-Ланке. Недавнее исследование, посвященное двум из этих групп, ченьчу и койя, позволило выявить не только тот факт, что мтДНК и Y-хромосомные наборы их предков являются уникальными среди других народов Южной и Западной Азии, но и то, что они характерны для древнейшего генетического наследия всего этого региона. По всей видимости, их предки-собиратели, мигрировавшие из Африки, образно говоря, заложили генетические семена для последующей дифференциации генофондов Западной и Восточной Евразии, и с тех пор представители этих народов испытали крайне ограниченный приток генов извне.
На Андаманских островах, расположенных в Бенгальском заливе, между Индией и Юго-Восточной Азией, живет целый ряд таких туземных групп, сохранивших архаические дописьменные культуры и обладающих резко выраженными чертами внешности так называемого африканского типа. К числу групп, менее всего затронутых внешним влиянием, относятся народности джарава и онге, представители которых живут на большинстве южных островов Андаманского архипелага. Результаты недавних генетических исследований, находящиеся в процессе анализа и изучения, способны помочь выяснить происхождение жителей этих островов как по материнской, так и по отцовской линиям. С материнской стороны онге и джарава имеют две характерные митохондриевые группы, которые позволяют безоговорочно отнести их к суперклану Манью, что можно считать аргументом в пользу версии об одном-единственном исходе (миграции) из Африки. Эти генетические линии демонстрируют непосредственную взаимосвязь с основными ветвями двух древних и в своем роде уникальных кланов Манью в Индии — М2 и М4. М2 — это наиболее древняя и своеобычная группа клана Манью; кроме того, она имеет наиболее общие компоненты мтДНК среди всех вышеупомянутых туземных групп Индии. С отцовской стороны онге и джарава относятся только к клану Авель (наиболее редко встречающемуся из трех линий Y-хромосом выходцев из Африки и известному под общим названием группы D или азиатского YAP). Это вполне согласуется с точкой зрения о том, что онге и джарава остаются изолированными группами еще с тех самых пор, когда их предки, кормившиеся собирательством на прибрежной полосе, бродили вдоль побережья Индийского океана. Другие туземные группы на этих островах, именуемых Большими Андаманскими, также имеют некоторые отличия и в культуре, и во внешнем облике. С генетической точки зрения, поскольку они принадлежат к ветвям тех же уникальных генетических линий клана Манью, преобладающая часть их Y-хромосом восходит к Сифу, одному из трех сыновей Адама выходцев из Африки .
Еще далее на пути древней миграции нас ждет встреча с так называемыми негритосами — коренными жителями Малайского полуострова, которых обычно называют народностью семанг (см. Фото 14). Эти люди — по-видимому, наиболее реальные претенденты на титул потомков древних собирателей на прибрежной полосе. Другая туземная группа, вполне возможно, являющаяся потомками тех же собирателей в Индокитае и на Малайском полуострове — это так называемые туземные малайцы, которые с точки зрения физического облика занимают промежуточное положение между народностью семанг и монголоидами (см. главу 5). Самое поразительное здесь заключается в том, что они являются своего рода хранителями ключей к древнейшим ветвям линии мтДНК, характерной для монголоидного населения этого региона и известной как клан F. Эта материнская линия npe-F, родоначальница многих линий в Восточной Азии, присутствующая у туземных малайцев, является у них общей с другими архаическими группами, обитающими на других островах Больших Андаманских островов, о которых мы уже говорили выше, а также с жителями Никобарских островов, расположенных к югу от Андаманских.

Фото 14. Автор этой книги проводит генетические обследования у жителей Семанга, которые, весьма вероятно, являются реликтовыми потомками народов первого маршрута собирателей на прибрежной полосе, совершивших исход из Африки.
В этой и следующей главах я расскажу более подробно о различных народах и поселениях, оставленных мигрантами — участниками первого великого переселения вдоль береговой линии Индийского океана. Как всегда, главное здесь заключается в том, что генетическое древо говорит нам: это переселение отнюдь не было ранним исходом из Африки, как то полагают некоторые археологи и палеонтологи. Это было продвижение передовых групп на пути заселения всего Старого Света.
Самое забавное заключается в том, что некоторые (если не сказать — единственные) наиболее эффектные археологические свидетельства, которые можно использовать для датировки этой миграции собирателей вдоль побережья Индийского океана, обнаружены не в Индии, не в Южной Аравии или в Африке, а почти в самом конце этого долгого пути — на Малайском полуострове, на о. Новая Гвинея и в Австралии. Поэтому мы отправимся сначала в гости к тем, кто проделал наиболее долгий путь вокруг побережья Индийского океана, — к аборигенам Новой Гвинеи и Австралии (см. Фото 15).

Фото 15. Лица нашего времени (сверху, слева направо): бабушка-китаянка с внуком, мальчик с нагорий Новой Гвинеи, англичанин и австралийский абориген из земли Арнхема.
Первые австралийцы
Пожалуй, из всех проблем, связанных с археологией этого региона, самые жаркие споры в научных кругах вызывал вопрос о том, какие именно племена первыми проникли в Австралию и когда это произошло. Вопрос «какие», звучащий вызовом мультирегионалистам, которые настойчиво повторяют свои аргументы о региональных различиях в форме черепов древних людей (см. главу 1), выглядит все менее и менее актуальным перед лицом массы генетических свидетельств того, что все современные туземные жители Австралии и Новой Гвинеи принадлежат кланам Насрин или Манью. Другими словами, они вовсе не являются каким-то изолированным исключением и тем более людьми более архаического типа или же потомками от браков людей современного типа с местными представителями вида Homo erectus. Однако эти неоспоримые свидетельства не мешают ретроградам-мультирегионалистам типа Шпильберга предпринимать все новые и новые попытки отстоять свои устаревшие взгляды, используя для этого древнейшие пробы ДНК. Тем не менее сегодня наука располагает свидетельствами, полученными на основе проб генетического материала той же самой ветви L3 потомков выходцев из Африки (более подробно о дискуссии, посвященной новейшим исследованиям в этой проблемной области см. примечание). Зато вопрос «когда» привлекает все большее внимание. Историки просто обожают нашпиливать события на даты вместо того, чтобы довольствоваться простой хронологической последовательностью, так что мне придется сначала поговорить о датах, хотя этот вопрос уместнее было бы затронуть в последнюю очередь.
Наиболее ранние общепризнанные археологические свидетельства расселения людей современного типа за пределами Африки вплоть до недавнего времени были найдены только в Австралии. Однако ситуация в этой области меняется на редкость быстро. До начала 1990-х гг. ученые не располагали доказательствами присутствия человека в Австралии и Новой Гвинее в эпоху ранее 40 тысяч лет тому назад. И вот теперь оказалось, что эти датировки, как и датировки артефактов в Европе, были обусловлены ограниченными возможностями радиоуглеродного метода. Однако теперь появились новые методы. Один из них, так называемый метод датировки по люминесценции кремния, позволил исследователям заглянуть гораздо дальше рубежной для радиоуглеродного метода даты 40 тысяч лет назад. В 1990 г. австралийские геологи и археологи Ричард Роберт и Рис Джонс, исследуя возраст первого поселения в пещерах на побережье Земли Арнхема в Северной Австралии, получили цифры от 50 до 60 тысяч лет тому назад . Земля Арнхема расположена прямо напротив острова Тимор — ближайшего к Австралии острова Индонезийского архипелага и, таким образом, наиболее вероятной отправной точки для первых мигрантов, направлявшихся в Австралию.
После этого датировки австралийских артефактов приняли весьма драматический характер и, образно говоря, пробили брешь в стене. В 1996 г. археолог Роберт Фаллагар проводил исследования наскальных рисунков на стоянке Джинмиум. Он обнаружил, что стена пещеры была сплошь покрыта плюсками, и, как сообщает Фаллгар, возраст обнаруженных в отложениях выкрошившихся фрагментов песчаника, испещренного древними рисунками, согласно оценке по двум независимым методам датировки может составлять 50 и 73 тысячи лет. Более того, ученый обнаружил даже каменные артефакты в культурных слоях, датируемых по методу термолюминесценции эпохой между 116 и 176 тысячами лет, что позволяет считать их наиболее ранними следами деятельности человека на Австралийском континенте, в два или даже три раза более древними, чем уже упоминавшиеся находки в пещерах на Земле Арнхема. Однако эта проблема была практически снята после того, как было доказано, что контаминация (загрязнение)песчинками из камней, обвалившихся со сводов, дала датировки совершенно фантастической древности для слоя земли вокруг этих артефактов. Другими словами, эти датировки были недостоверными, и анализ отдельных песчинок помог выявить и устранить неточности, связанные с контаминацией. Таким образом, возраст наиболее ранних следов деятельности человека в Австралии остался прежним — около 60 тысяч лет.
Впрочем, недавно были опубликованы работы, где приводятся близкие датировки следов деятельности человека: 62 тысячи лет тому назад. Эти датировки основаны на применении различных методов для анализа почвы вокруг скелета стройного анатомически современного человека, найденного неподалеку от озера Мунго в районе Вилландровых озер на юго-востоке Австралии. В одной из этих работ говорится, что для того, чтобы иметь столь почтенный возраст, первые обитатели Австралии должны были попасть на континент в глубокой древности, когда уровень океана был гораздо более низким. Подобная возможность проникнуть в Австралию благодаря низкому уровню океана могла предоставиться около 70 тысяч лет тому назад, что свидетельствует о том, что анатомически современные люди, обитавшие на берегах озера Мунго, были в числе древнейших обитателей континента, поскольку предыдущий столь же низкий уровень океана имел место на 80 тысяч лет раньше, то есть около 150 тысяч лет тому назад, еще до появления человека современного вида.
Одна-единственная возможность в истории
Пока ученые выдвигают всевозможные аргументы и контраргументы, мы с полным основанием можем говорить, что датировки находок на многих стоянках близки или совпадают, указывая, что человек современного типа уже присутствовал в Австралии примерно 60 тысяч лет тому назад. Примерно в то же время закончился период очень низкого уровня Мирового океана, когда огромные массы воды были сосредоточены в ледниковых полярных шапках и уровень моря у берегов Австралии был примерно на 80 м ниже современного. И хотя считается, что точная дата наиболее низкого уровня океана — около 70 тысячи лет тому назад, уровень Мирового океана оставался очень низким вплоть до 65 тысячи лет назад, после чего начался его быстрый подъем.
Присутствие человека на северо-западном побережье Австралии 62 тысячи лет тому назад не вполне согласуется с реальными условиями для его прибытия морским путем, поскольку период низкого уровня Мирового океана завершился примерно за 3000 лет до этого. Таким образом, если дата прибытия человека в Австралию, равная 62 тысячам лет тому назад, реальна, это означает, что мигрантам пришлось столкнуться с рядом проблем. Крайне низкий уровень океана, имевший место несколько ранее, делал шансы мигрантов перебраться с Тимора в Австралию несравненно более высокими. На контурной карте глубины моря у северного побережья Австралии мы видим, что расстояние, разделявшее Австралию и Тимор, увеличилось со 170 км при уровне океана —80 м и 220 км при уровне —40 м до 470 км при уровне —20 м (см. рис. 4.1). Различие между 170 км и 220 км на первый взгляд представляется не слишком существенным, но дело в том, что за последние 20 тысяч лет расстояние 180 км было абсолютным пределом досягаемости для плаваний между островами во всем мире. Древнейшим свидетельством этой максимальной дальности плавания, которая подтверждается археологическими находками, можно считать заселение острова Манус, расположенного к северу от Новой Гвинеи. Это заселение имело место около 20 тысяч лет тому назад. Таким образом, если говорить о заселении Австралии в результате длительного плавания, то единственная реальная возможность для подобного плавания могла предоставиться людям где-то между 65 и 70 тысячами лет тому назад. Тогда они могли успеть создать те самые памятники древнего искусства, которые обнаружены археологами в Австралии и датируются временем между 55 и 62 тысячами лет тому назад. В любом случае заселение Австралии в результате случайного дрейфа представляется крайне маловероятным сценарием, если принять во внимание генетические свидетельства присутствия в Австралии многочисленных материнских линий-родоначальниц. Подобная дата появления человека на континенте вполне согласуется с генетической датировкой расселения мигрантов в Австралии, составляющей 68 тысяч лет тому назад.
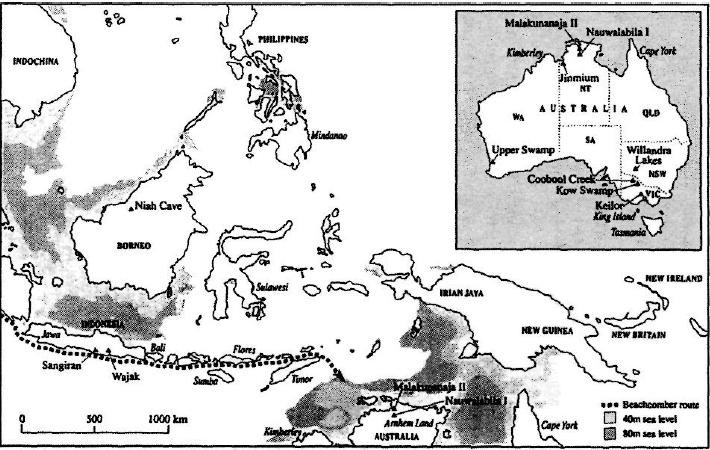
Рис. 4.1
Нам представляется вполне уместным перенести свидетельства деятельности человека, датируемые временем около 60 тысяч лет тому назад, в другой контекст. Дело в том, что на некоторых древнейших стоянках человека в Австралии найдены характерные груды раковин морских моллюсков. Это дает основание предполагать, что первые люди, прибывшие на северо-западное побережье Австралии, по-прежнему кормились собирательством на прибрежной полосе. А если это так, то у нас нет практически никакой возможности найти следы их первоначального присутствия на берегу океана, так как Тиморское море сегодня продвинулось на добрых 500 км в глубь прежнего материка, накрыв своими водами древнейшие поселения человека на берегу. Первые реально датированные свидетельства присутствия человека на северо-западе Австралии после периода минимально низкого уровня океана, естественно, были получены на горизонтах, расположенных выше высокого уровня океана. Подобный высокий уровень отмечался уже между 55 и 60 тысячами лет тому назад, когда уровень океана был ниже современного всего на 20 м, а береговая линия была уже близка к своим современным очертаниям. Береговая линия поблизости от пещер на Земле Арнхема в эту эпоху хорошо согласуется с расчетами предполагаемого уровня океана и датировкой следов деятельности человека в этих местах. Эта датировка, выполненная геологом Ричардом Робертом, дает цифры около 50—60 тысяч лет тому назад. При подобной, обусловленной изменением уровня океана, паузе между заселением и археологическими свидетельствами о деятельности человека следующий минимум уровня океана, отмечавшийся около 55 тысяч лет тому назад, был уже слишком запоздалым для трансокеанских плаваний и археологически документированных данных о присутствии человека в эпоху ранее 50 тысяч лет тому назад.
Хотя заселение ближайшего соседа Австралии, огромного острова Новая Гвинея, по всей видимости, представляло такие же проблемы для потенциальных колонистов, как и заселение Австралии, между ними все же были существенные различия. Есть вполне реальная вероятность того, что Новая Гвинея была заселена даже раньше, чем Австралия. Прежде всего возможны два пунктирных маршрута через острова Восточной Индонезии (см. рис. 4.1). Как мы уже знаем, для того чтобы плавание по прямому пути в Австралию с острова Тимор, через Тиморское море, имело шансы на успех, уровень океана в ту эпоху должен был быть предельно низким. Между тем маршрут в Новую Гвинею через о. Сулавеси и Молуккские острова позволял преодолевать гораздо меньшие расстояния между островами и практически не зависел от колебаний уровня океана.
Все острова на пути в Австралию и Новую Гвинею находились в пределах прямой видимости друг от друга. У этих островов издревле существует очень крутая береговая линия, и маршрут путешествия на Новую Гвинею включал в себя очень мало участков, пролегавших по континентальному шельфу. В результате перепады уровня океана почти не сказывались на расстоянии между островами, и этот маршрут можно считать одинаково легким (или трудным) в любую эпоху после того, как наши предки прибыли на о. Борнео сухопутным путем.
Можно задать вопрос: имеются ли доказательства того, что предки современных аборигенов Австралии не воспользовались этим альтернативным северным, немногим более протяженным маршрутом, ибо в ту эпоху Австралия могла быть соединена сухопутным перешейком с Новой Гвинеей? Генетические данные со всей определенностью показывают, что между аборигенами Австралии и Новой Гвинеи не существует общих генеалогических клановых линий и что жители Новой Гвинеи в генетическом отношении столь же далеки от аборигенов Австралии, как и от любого другого народа за пределами Африки Более того, оценки времени расселения туземных кланов мтДНК Новой Гвинеи дают цифру 77 тысяч лет тому назад, что значительно старше периода активного расселения австралийцев. Это говорит о том, что Новая Гвинея, по всей вероятности, могла быть заселена людьми гораздо раньше Австралии, а также о том, что австралийские генетические кланы не являются потомками кланов Новой Гвинеи. Другими словами, первые мигранты в Австралию могли отдать предпочтение маршруту от одного необитаемого острова к другому, хотя это и создавало ряд трудностей.
Однако при всей своей заманчивости эта генетическая история, хорошо согласующаяся с исключительной древностью и взаимосвязью между колонизацией Австралии и Новой Гвинеи, не согласуется с надежно датируемыми археологическими находками, сделанными на островах Восточной Индонезии или в Новой Гвинее. Вплоть до недавнего времени ученые не располагали свидетельствами о присутствии человека на всем протяжении обоих этих трансостровных маршрутов через Восточную Индонезию около 40 тысяч лет назад, не говоря уж о 70 тысячах лет назад. Однако новые поразительные находки на острове Флорес, одном из самых крупных островов на южном маршруте, способны в корне изменить сложившиеся представления. Глубокая (около 7 м) яма в пещере Лянг Буа, находящейся на острове Флорес, таила в себе удивительные находки, возраст которых — около 840 тысяч лет! Лянг Буа может сыграть значительную роль в качестве источника Ценнейших культурных и палеонтологических свидетельств, относящихся к большей части этого периода, включая и дату прибытия на остров человека современного типа. В ту эпоху в пещере обитали представители двух видов человека. Первые жители явно не были людьми современного типа. В настоящее время даты появления первых людей современного типа уточняются, однако они могут оказаться наиболее ценными для датировки наиболее раннего присутствия человека современного типа в Восточной Азии.
Таким образом, если, следуя гипотезе о климатическом «окне», допустить, что фактически колонизация Австралии имела место в период между 65 и 70 тысячами лет тому назад, а острова Флоры и даже Новая Гвинея были заселены около 75 тысяч лет тому назад, мы вправе задать вопрос: а за сколько тысяч лет до этого момента люди современного типа должны были покинуть Африку, чтобы проделать столь дальний путь вдоль побережья южных морей. Понятно, что ответить на такой вопрос крайне сложно. Прежде всего, мы не можем с уверенностью сказать, сколько времени могло потребоваться нашим собирателям, чтобы в буквальном смысле «проесть» себе путь к берегам Тимора или Сулавеси, если древние люди не оставили на нем никаких следов своего продвижения. Как показывают данные оценки уровня океана в ту эпоху, прибрежная полоса, по которой брели наши далекие предки 60—80 тысяч лет тому назад, давно ушла под воду, и нам трудно рассчитывать найти следы древних миграций, не опустившись на дно морское.
Древнейшие реликты маршрута собирателей
Существует и другая проблема. Если наши праотцы покинули Африку более 70 тысяч лет тому назад, они вполне могли использовать и создавать каменные орудия старого типа, аналогичные тем, которыми пользовались представители архаического вида Homo sapiens, которые, вполне возможно, покинули Африку по меньшей мере 100 тысяч лет тому назад (см. Пролог). Такие орудия эпохи Среднего палеолита во множестве встречаются в Индии и датируются временем около 160 тысяч лет тому назад, поэтому они не могут считаться надежными маркерами прихода людей анатомически современного типа Орудия эпохи Среднего палеолита в изобилии встречаются в песках береговой полосы южного побережья Аравийского полуострова, однако они не поддаются сколько-нибудь надежной датировке. Йемен, являющийся, по моему убеждению, пунктом прибытия мигрантов после исхода из Африки, пока что не привлек к себе достаточного внимания археологов, однако там тоже были найдены каменные орудия, аналогичные африканским орудиям позднего Среднего Каменного века.
Археологическая датировка ранних находок в Австралии и свидетельства еще более ранней колонизации Новой Гвинеи получили серьезную поддержку в результате открытия палеолитической культуры Кота Тампан в долине Ленггонг, что в Пераке на Малайском полуострове, на расстоянии примерно 2/3 пути из Африки в Австралию. Вначале эта культура была идентифицирована по находкам крупных, необычной формы и достаточно грубо сделанных каменных орудий, обработанных только с одной стороны, которые, как считали археологи в 1960-е гг., были делом рук представителей более ранних видов человека. Но когда была проведена переоценка возраста геологических слоев, в которых были найдены эти орудия, стало ясно, что они относятся к гораздо более позднему времени. Более заметный интерес к этим находкам возник в 1975 г., когда Том Харрисон, колоритный куратор Саравакского музея, попытался связать эти находки с грандиозным извержением вулкана Тоба на о.Суматра. Его предположение, основанное на исследовании вулканического пепла вокруг найденных на стоянке орудий, вызвало настоящую сенсацию, лихорадочный интерес к которой пошел на спад лишь совсем недавно.
Пожалуй, никто из ученых не отдал изучению культуры Кота Тампан и долины Ленггонг столько сил и времени, как археолог Зурайна Маджид из университета Сайнс Малайзия в Пенанге. Ее обширные исследования целого ряда стоянок этой культуры в долине Ленггонг показали, что местная культура каменных орудий, вполне возможно, просуществовала со дня грандиозного извержения вулкана Тоба до 7 или даже 4 тысяч лет тому назад. А если это так, то находки артефактов этой культуры могут дать ответ на один из самых волнующих вопросов, касающихся этих странных орудий овальной формы: кто были их создатели? Дело в том, что эти орудия не идут ни в какое сравнение с гораздо более сложными артефактами.
Орудия, обработанные гораздо тщательнее их, создавались в Африке и Европе людьми архаического типа гораздо раньше, так что с какой стати мы должны считать, что каменные сколы, найденные в вулканическом пепле, были сделаны людьми современного типа, жившими в эпоху извержения Тоба?
Двое наиболее авторитетных специалистов по эпохе палеолита в Юго-Восточной Азии, австралийские археологи Питер Веллвуд и Сандра Боудлер, согласны с Зурайной Маджид и Томом Харрисоном в том, что эти орудия скорее всего были созданы людьми анатомически современного типа. Прежде всего надо сказать, что датировки большинства каменных орудий, найденных в долине Ленггонг, относятся к слишком позднему времени, чтобы их могли создать люди более архаического типа. Во-вторых, на Малайском полуострове, не говоря уж о долине Ленггонг, не найдено ни единого образца останков человека типа, предшествовавшего современному. Визитной карточкой Зурайны Маджид стала вызвавшая массу откликов находка так называемого перакского человека, сделанная членами ее группы в 1990 г. в пещере Гунуг Рунту, которая расположена в долине Ленггонг. Этот полный скелет человека современного типа, окруженный каменными орудиями этого класса, был описан специалистами как останки, имеющие характерные австрало-меланезийские признаки. Его возраст — около 10 тысяч лет. Поэтому недвусмысленная связь этих каменных орудий с человеком современного типа решительно опровергает утверждение о том, будто каменные орудия культуры Кота Тампан слишком грубы для того, чтобы считаться делом рук человека современного типа. Таким образом, по крайней мере — на сегодня, перакский человек бесспорно является наиболее убедительным свидетельством того, что орудия архаического типа, найденные в вулканическом пепле, были созданы человеком современного типа.
Другой уважаемый специалист в области археологии Юго-Восточной Азии — Ричард Шатлер. Он выразил более обобщенное мнение, что подобные орудия были впервые принесены на острова Юго-Восточной Азии людьми вида Homo sapiens примерно 70 тысяч лет тому назад. (Понятие «островная Юго-Восточная Азия», или ОЮВА, в отличие от континентальной Юго-Восточной Азии, включает в себя все острова Индонезии и Филиппин, за исключением Западной Новой Гвинеи, но включая все острова Борнео.) Шатлер предостерег против поспешного вывода о том, будто такие орудия являются отражением технической отсталости, соглашаясь с другими исследователями в том, что качество наличного сырья для производства орудий во многом определяло их уровень и что для изготовления более сложных орудий, например ножей, вполне мог использоваться бамбук.
Так каков же возраст пепла на стоянках культуры Кота Тампан? Когда он оценивался впервые, несколько десятилетий тому назад, тогдашние методы показали такую цифру: около 31 тысячи лет тому назад. Однако подобная датировка извержения вулкана Тоба вызывала возражения со стороны геологов и даже археологов, в частности Питера Беллвуда. Главная проблема здесь заключалась в том, что в те времена у вулкана Тоба не было сколько-нибудь значительных извержений. Последнее крупное извержение вулкана Тоба, крупнейшее в мире за последние 2 млн. лет, произошло гораздо раньше, примерно 71—74 тысячи лет назад. Не так давно ряд видных геологов, включая и автора первоначальной датировки, пришли к согласию во мнении о том, что возраст пепла, в котором были найдены эти орудия, действительно составляет 74 тысячи лет. Эта датировка имеет огромную важность.
Если каменные орудия культуры Кота Тампан были созданы людьми современного типа, то в таком случае они являются наиболее древними надежно датируемыми свидетельствами деятельности человека современного типа за пределами Африки. Таким образом, складывается впечатление, что предки аборигенов Австралии покинули Африку и, продвигаясь вдоль береговой полосы, прибыли в Малайзию задолго до катастрофического извержения Тоба.
Но, пожалуй, фактором более важным, чем точность датировки, является то, что подобная взаимосвязь между каменными орудиями и вулканическим пеплом на стоянках в Малайзии позволяет утверждать, что древнейшие жители Индии и Пакистана тоже оказались непосредственно в зоне этой величайшей природной катастрофы в истории человечества. Извержение вулкана Тоба явилось самым грозным бедствием за последние 2 млн. лет. Подхваченное ветром громадное облако пепла, взметнувшееся над вулканом, двинулось на северо-запад и накрыло практически весь Индийский субконтинент. Даже в наши дни в этом регионе находят слои пепла толщиной в несколько метров, а в слое пепла на двух древних стоянках в Индии обнаружены орудия, датируемые эпохой Среднего и Верхнего палеолита. Важный вывод, который можно сделать на основе этой взаимосвязи между орудиями и пеплом, заключается в том, что в результате извержения Тоба Восток и Запад разделила глубокая и генетически стерильная борозда. Население Индии было восстановлено путем волн повторной колонизации как с Востока, так и с Запада. И подобная борозда, как мы вскоре увидим, действительно существует на генетической карте Азии.
Несмотря на близость вулкана Тоба к Пераку, облако вулканического пепла, взметнувшееся над ним, лишь вскользь задело Малайский полуостров. Люди культуры Кота Тампан оказались в числе несчастливцев, но большая часть полуострова осталась практически незатронутой извержением. Некоторые исследователи на основе сравнительного анализа морфологии черепов высказывают предположение, что туземные «негритосы» племени семанг, издревле кормившиеся охотой и собирательством, которые и сегодня живут в той же самой части густых влажных тропических лесов на севере Малайского полуострова, являются потомками той этнической ветви, к которой принадлежал и перакский человек. Непрерывный характер развития культуры Кота Тампан, версию о котором выдвинула Зурайна Маджид, позволяет провести прямую линию, ведущую к каменным орудиям 74-тысячелетней древности, обнаруженным в вулканическом пепле Тоба. В следующей главе мы рассмотрим материальные свидетельства и артефакты, связывающие нынешних людей племени семанг с древнейшими первопроходцами современного типа, мигрировавшими вдоль побережья Индийского океана.
В процессе съемок документального фильма «Истинная Ева», с которым перекликается наша книга, канал «Дискавери» осуществил финансирование исследования генетического наследия туземных групп Малайского полуострова. Это исследование я провел в тесном сотрудничестве с английским генетиком Мартином Ричардсом и рядом ученых из Малайзии. Это исследование явилось составной частью гораздо более обширной программы, осуществляемой генетиками ряда стран Азии. Результаты исследований мтДНК оказались впечатляющими: у 3/4 представителей этнической группы семанг (принадлежащий к различным типам «негритосов») были выявлены уникальные локальные варианты линий Манью и Насрин, почти не имеющие внешних примесей, что хорошо согласуется с мнением о том, что их предки могли прибыть в эти края вместе с первыми мигрантами — собирателями на прибрежной полосе. Найденные у них линии восходят непосредственно к самым корням Манью и Насрин (двух первых дочерей Евы за пределами Африки — см. главу 1). Выявленная у них линия Манью не встречается более ни у какой этнической группы в Юго-Восточной или Восточной Азии (и вообще ни у какого другого этноса) и, хотя она и подверглась активному генетическому дрейфу в результате сокращения численности ее носителей, она тем не менее сохранила достаточно широкое многообразие, что позволяет утверждать, что ее возраст составляет около 60 тысяч лет. Что касается их линии со стороны Насрин, R21, то она происходит непосредственно от Рохани, генетической дочери Насрин. Подобное отсутствие каких бы то ни было связей с другими этническими группами Евразии хорошо согласуется с гипотезой о том, что эти люди, прибывшие в эти края много тысячелетий назад, оставались в полной генетической изоляции в джунглях Малайского полуострова.
Есть и другие свидетельства из данного региона, позволяющие отнести появление анатомически современного человека на Дальнем Востоке ко времени около 70 тысяч лет тому назад. Это прежде всего знаменитый люцзяньский скелет, найденный на юге Китая. Люцзяньский скелет (см. Фото 16), от которого уцелели хорошо сохранившиеся череп и несколько костей, был найден в 1958 г. в пещере Тонтяньянь в Гуанси-Чжуаньском автономном районе. Его обнаружили крестьяне, собиравшие гумус и удобрения. Нет никаких сомнений том, что этот скелет принадлежал человеку современного типа, но в вопросе о его возрасте сразу же возникли разногласия во мнениях.
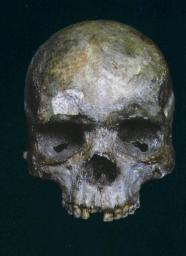
Фото 16. Люйцзяньский череп Китай) с толстокостными, но современными чертами. По данным уточненной датировки , его возраст составлят по меньшей мере 68 тыс. лет, что свидетельствует о весьма раннем распространении человека анатомически современного типа из Африки, имевшем место еще до извержения вулкана Тоба.
Есть данные о датировке по изотопам урана, согласно которой возраст скелета составляет около 67 тысяч лет, однако подобная датировка вызывала сомнения из-за ее несоответствия возрасту того надежно датируемого геологического слоя, в котором был обнаружен скелет. В декабре 2002 г. группа китайских исследователей во главе с геологом Шень Гуаньюнем сообщила о результатах проведенного ею повторного анализа стратиграфических параметров пещеры (а также нескольких соседних пещер) и датировки черепа. Ученые пришли к выводу, что череп следует отнести к эпохе между 70 и 130 тысячами лет тому назад и что он в любом случае не моложе 68 тысяч лет. Череп был найден в так называемой интрузивной брекчии — яме с грудой обломков и отходов вторичного происхождения, содержащей материальные следы различных эпох. Благодаря тому, что их статья об этих исследованиях была опубликована в журнале «Journal of Human Evolution», нижняя временная граница этого периода (68 тысяч лет) представляется вполне солидной датировкой, поскольку она основана на многочисленных датировках возраста каменных капов, закрывающих брекчию. (Каменный кап образуется, когда вода, стекающая со стен, приводит к осаждению кальцита на стене или полу.) Наиболее вероятный возраст этих капов — от 111 до 139 тысяч лет. Эти данные, полученные на основе изучения фрагментов капов и кальцитов в самой брекчии, представляются чрезмерно завышенными. Но главный вывод этих исследований заключается в том, что вполне реальная нижняя граница появления на Дальнем Востоке анатомически современного человека является дополнительным аргументом в пользу гипотезы о раннем исходе человека из Африки через устье Красного моря еще до извержения вулкана Тоба, имевшего место около 71—74 тысяч лет тому назад. Дело в том, что для того, чтобы попасть в Китай около 68 тысяч лет назад, предкам этих людей необходимо было проделать путь длиной в четверть окружности экватора.
Некоторые ученые по-прежнему убеждены в том, что аборигены Австралии представляют собой потомков миграции из Африки, более ранней, чем исход родоначальников будущих народов и рас Европы, Азии и Америки. Однако наши генетические следы говорят совсем иное. Ряд исследований присутствия генетических линий материнских кланов в Австралии показал, что все они принадлежат к двум крупнейшим неафриканским суперкланам — Манью и Насрин. А обширные исследования Y-хромосом позволили установить, что все мужские линии аборигенов Австралии относятся к тому же клану Адама выходцев из Африки, что и все прочие обитатели земного шара за пределами Африки, и принадлежат к линиям Сифа и Каина. Та же картина наблюдается и при анализе генетических маркеров, передаваемых через обоих родителей. Другими словами, вся совокупность генетических свидетельств с полной определенностью говорит о том, что австралийцы также являются потомками того самого единственного исхода из Африки. Логика такого подхода в сочетании с рассмотренными выше археологическими свидетельствами позволяет отнести дату прибытия людей современного типа на Малайский полуостров ко времени до 74 тысяч лет тому назад, а дату их появления в Австралии — до 70 тысяч лет тому назад. Это вполне согласуется с датой исхода из Африки, определяемой на основе следов собирательства на прибрежной полосе. Как мы уже говорили в главе 1, одним из мотивов исхода с африканского континента могла стать массовая гибель даров моря на всем побережье Красного моря, вызванная резким повышением солености его вод, начавшимся около 80 тысяч лет тому назад. По моим оценкам затрат времени на миграцию из Африки вдоль побережья Индийского океана, собирателям на прибрежной полосе могло потребоваться около 6000 лет, чтобы «проесть» себе путь вдоль побережья до Перака, и еще около 4000 лет, чтобы достичь берегов Австралии. Такие временные рамки вполне соответствуют паузе между исходом из Африки около 80 тысяч лет тому назад и появлением в Австралии около 70 тысяч лет тому назад. Эта первая дата хорошо согласуется с оценками времени исхода африканского комплекса L3 на основе датировки по молекулярным часам (см. также главу 1).
Взглянуть в лицо матери — родоначальницы миллиарда семейств
Главный вопрос, не дающий покоя журналистам и кинематографистам после выхода в 1988 г. знаменитой статьи в «Newsweek» об «истории» митохондриевой Евы, заключается в том, как могли выглядеть она и ее потомки. Вполне возможно, что все они имели весьма и весьма непохожую внешность и отличались друг от друга даже больше, чем случайно взятые представители разных рас и народов, живущие ныне за пределами Африки. Дело в том, что они, по крайней мере, происходят от одного и того же древнейшего прапредка, и все имеющиеся свидетельства говорят в пользу того, что вариации в пределах индивидуальных групп даже в наши дни уменьшаются по мере удаления из Африки. Однако что касается различий между разными группами, например, китайцами, коренными жителями Америки и аборигенами Полинезии, Меланезии и Австралии, то они, наоборот, нарастают по мере удаления от Африки; таким образом, можно говорить о том, что различия между так называемыми расами по мере удаления от Африки усиливаются. А теперь позвольте мне взглянуть на вопрос с несколько иной стороны. Моя дражайшая теща, которая была китаянкой, однажды сказала мне (причем, что называется, на полном серьезе), что европейцы кажутся ей совершенно одинаковыми, так сказать, все на одно лицо. Но европейцы с полным правом могут сказать то же самое о самих китайцах. Таким образом, налицо явный парадокс, который необходимо решить, прежде чем продолжить составление карты генетической диаспоры за пределами Африки.
Суть вопроса заключается в том, каким образом возникло это разделение. То, что на первый взгляд выглядит парадоксом, на самом деле является результатом многократного и неравномерного дробления групп мигрантов, сопровождавшегося активным генетическим дрейфом. Это можно объяснить следующим образом: по мере того как мигранты-переселенцы разделялись на все большее и большее количество ветвей, первоначальные генетические вариации между ними оказывались как бы сосредоточенными в пределах различных групп, предков будущих этносов, которые живут в наши дни на территории Евразии и Америки. За последние 50 тысяч лет члены этих разделившихся и изолированных групп диаспоры выходцев из Африки начали внешне более походить на своих сородичей, чем на представителей других групп. Так возникли различия между группами. И хотя впоследствии я попытаюсь доказать, что многие из подобных различий были не более чем эволюционно обусловленными проявлениями адаптации к новым условиям окружающей среды, некоторые из них выходят далеко за рамки случайного разлома этнического «пирога».
Чем же их можно объяснить? Ответ заключается в том же самом процессе, который «сработал» во время исхода из Африки первой небольшой группы мигрантов. Давайте представим — как мы поступили в одной из предыдущих глав, — что многообразие представителей рода человеческого подобно мешочку с камешками разных пород. Предположим, на момент исхода из Африки, имевшего место около 75 тысяч лет тому назад, в мешочке было 10 тысяч камешков. После того как мигранты переправились через Красное море, многообразие резко сократилось, и в колонии-родоначальнице осталось всего 250 камешков. Подобное сокращение известно под названием «эффекта родоначальников». И если члены этой первой колонии жили на юге Аравии изолированной группой на протяжении многих тысяч лет, одни из «камешков» не оставили себе подобных и их линии пресеклись, а другие линии, наоборот, заняли доминирующее положение. В результате число камешков в мешочке уменьшилось еще больше. Как мы уже говорили в главе 1, это и есть генетический дрейф.
В дальнейшем население колонии дробилось на неравные части. Когда численность группы стала слишком высокой и данный участок берега более не мог прокормить ее, от нее одна за другой отделялись небольшие группы первопроходцев, которые покидали обжитое побережье и направлялись в глубь материка, продвигаясь в основном вдоль русел рек. Эти небольшие группы несли с собой в мешочке еще меньшее число первоначальных генетических камешков мигрантов-выходцев из Африки. Затем они подвергались воздействию того же «эффекта родоначальников» и мигрировали дальше, продолжая генетический дрейф.
А теперь позвольте задать вопрос: что является кроной генеалогического древа и что — его стволом? Были ли собиратели на прибрежной полосе всегда и везде большинством, а первопроходцы, продвигавшиеся вдоль русла рек, — меньшинством, или же основная масса мигрантов в какой-то момент устремлялась в глубь материка? Такой вопрос может показаться уделом академической схоластики, но если наши журналисты действительно хотят заглянуть в лицо людям современного типа, жившим как можно ближе к моменту исхода, вопрос, на который мы должны ответить, будет звучать так: какие именно представители неафриканских народов в наши дни сохранили более всего вариантов тех самых «камешков» из мешочка древнейших обитателей Африки?
Нуклеарные генетические маркеры: чтобы найти больше древних маркеров, нужно иметь больше камешков в мешочке
Чтобы продолжить рассмотрение вопроса о многообразии, мы не можем полагаться на одни только генетические маркеры нашей митохондриевой ДНК или Y-хромосомы. Если уж говорить о них, надо признать, что в отцовских и материнских линиях ныне живущих на земном шаре народов уцелело всего по одному камешку от каждого из их африканских прародителей. Как я уже говорил, именно этот факт является доказательством одного-единственного исхода из Африки (см. главу 1). Понятно, что у одной линии нет и не может быть многообразия. Нет, если мы действительно хотим сравнить генетическое многообразие наших предков-африканцев с наследием остального населения земного шара, нам необходимо найти такие типы генетических маркеров, которые сумели пережить пресловутый эффект родоначальников и генетический дрейф исхода, сохранив достаточно широкое многообразие и количественный состав представителей каждой линии. Поэтому нам придется заняться анализом остальных компонентов нашей генетической структуры, которые присутствуют в ядрах клеток нашего тела и не связаны с тем или иным конкретным полом.
Приводимая мной аналогия с камешками, естественно, была нарочито упрощенной. Если обратиться к рассмотрению той на удивление малой части нашей ДНК, которая действительно несет некую полезную нагрузку, нетрудно заметить, что у каждого гена существует от одного до целой дюжины вариантов, несколько отличающихся друг от друга
Такие варианты обычно могут быть объединены в составе семейного генного древа. Основание каждого генного древа и его нижние ветви обычно являются по преимуществу африканскими, тогда как верхние ветви и побеги, а иногда и некоторые из нижних ветвей встречаются в основном за пределами Африки. Кроме того, мы можем рассмотреть и нефункционирующие части нашей ДНК, где накапливается громадная масса совершенно бесполезной информации, которая, однако, может использоваться генетиками в качестве своего рода почтового ящика. Эти излишние массивы данных могут накапливаться до тех пор, пока они не начнут составлять около 10% общего объема нашей ДНК. В то же время отсутствие таких специфических массивов у того или иного человека означает, что он сохраняет большинство признаков своего родового или африканского типа.
Если мы посмотрим, у представителей каких именно этнических групп более всего сохранилось генетическое наследие наших далеких африканских предков, нам придется вернуться в Австралию и Новую Гвинею. Эти два обширных массива суши образуют остатки грандиозного и сегодня частично затопленного водами океана континентального шельфа материка Сахул, где сохранилось значительно больше первичных генетических типов африканских предков, чем в любом другом регионе мира за пределами Африки. Однако в наши дни аборигены Новой Гвинеи и Австралии мало походят друг на друга, и сегодня единственной крупной этнической группой, по внешнему облику напоминающей современных африканцев, являются аборигены Меланезии, имеющие кожу очень темного цвета и густые вьющиеся волосы. Но и в этом случае аборигены горных районов Новой Гвинеи, которые считаются «наименее изменившимися» представителями меланезийской расы по сравнению с людьми плейстоценового периода, имеют крепкокостное строение (толстые кости и выраженные надбровные дуги — см. главу 5), чем сами современные африканцы. Однако в таком случае возникает вопрос: а как выглядели африканцы в эпоху исхода людей современного типа, около 80 тысяч лет тому назад? Можно практически наверняка утверждать, что их внешний облик был мало похож на современных тонкокостных банту, жителей долины Нила, пигмеев или людей народа кунг. Ископаемые останки черепов кафзехского и скульского человека, датируемые временем между 90 и 120 тысячами лет тому назад, являются куда более крепкокостными, чем черепа современных африканцев. Это свидетельствует о том, что жители Африки могли измениться куда больше, чем горцы Новой Гвинеи. Прожив несколько лет в Новой Гвинее и Австралии, я могу сказать, что человек с внешностью, воссозданной при реконструкции одного из черепов кафзехского человека в ходе съемкок нашего документального фильма, был бы сегодня практически неотличим от аборигенов Новой Гвинеи, но ему вряд ли нашлось бы место среди жителей Африки (см. Фото 14 и 15).
Генетические следы исхода среди реликтовых этнических групп, живущих недалеко от Африки
Какие же еще народы в генетическом отношении являются более близкими к африканцам в результате того, что их исконные земли занимают особое положение на пути собирателей-мигрантов? Что может нам дать сопоставление встречаемости и сохранения архаических наследственных генотипов у африканцев и неафриканских народов? Здесь следует выделить два региона. Ближайшими (помимо аборигенов Австралии и Новой Гвинеи) в генетическом смысле к африканцам народами следует считать архаические туземные группы, живущие в Пакистане и на юге Аравийского полуострова. За ними следуют европейцы и жители Индии. И в самом конце этой «очереди» находятся народы Восточной Азии и Америки, проделавшие долгий путь миграции и сохранившие меньше всего следов генетического многообразия своих африканских предков.
Ключевые позиции в сохранении древнего генетического многообразия Африки занимают туземные группы Пакистана и Южной Аравии, то есть народы, издревле живущие на пути предполагаемого южного маршрута исхода из Африки. В пользу этой версии говорит целый ряд свидетельств. На южном побережье Аравийского полуострова живет изолированная народность хадрамаут, которую некоторые исследователи относят к австралоидам. Их материнская генетическая структура включает в себя 40% африканских генных линий, но хотя некоторые из этих маркеров связаны с родоначальницей-Евой выходцев из Африки, большинство этих линий проникли сюда из Африки в более поздние времена. Далее, на всем протяжении океанского побережья Индийского субконтинента встречаются этнические группы, занимающие в генетическом отношении более близкое положение к африканским корням исхода, чем жители районов Азии, лежащих дальше к Востоку. Этнические группы Индии, как кастовые, так и родоплеменные, стали объектом детального исследования наличия у них нуклеарных аутосомных (не связанных с половой сегрегацией) маркеров. Они, как было установлено, сохранили более высокую встречаемость африканских наследственных типов, чем европейцы и любые другие расы и этнические группы Азии.
Существуют и другие признаки того, что в Пакистане в значительной мере сохранилось генетическое многообразие Древней Африки. В то время как жители Пакистана в Целом имеют общие древние составляющие ДНК с Индией, Европой и Средним Востоком, они обладают также некоторыми уникальными маркерами, которые не встречаются более нигде за пределами Африки. Существуют этнические группы, имеющие очень древние наследственные корни. Одна из архаических групп так называемых негритосов, макрани, живет в дельте Инда и на океанском побережья Белуджистана в Пакистане. Есть предположение, что предки макрани пришли в эти места после исхода из Африки. У макрани выявлен специфически африканский Y-хромосомный маркер, который ранее встречался только в Африке и считался характерным для региона, прилегающего к Сахаре. Тот же генетический маркер, правда, несколько реже, встречается и у других этнических групп южного Пакистана, Саудовской Аравии и Объединенных Арабских Эмиратов. Кроме того, он имеет весьма высокую встречаемость в Иране. Есть и другой уникальный Y-хромосомный маркер, который встречается за пределами Африки только в этом регионе Пакистана. Наконец, третий древний Y-хромосомный маркер указывает на Пакистан как на очень ранний пункт, где началось разделение ветвей. Именно здесь присутствует ранняя ветвь потомков Адама выходцев из Африки, отличающаяся высокой встречаемостью в Пакистане и несколько меньшей — в Индии (особенно у изолированных племен). Кроме того, она отмечается и далее к северу, на Среднем Востоке, в Кашмире, Центральной Азии и Сибири. Тот факт, что этот маркер не выявлен далее к востоку в Азии, свидетельствует о том, что единственным путем, которым он мог проникнуть в Центральную Азию, была миграция с юга, вверх по течению Инда в Кашмир и далее на север.
Истоки происхождения европейцев: где появилась на свет Насрин?
Ключевые позиции Южной Азии в качестве средоточия генетического многообразия африканских линий заставляют предположить, что именно там, в Южной Азии, произошло разделение ветвей мигрантов. Теперь нам следует сосредоточить внимание на том, какой биологической силой обладали генетические древа отцовских и материнских линий, оставившие следы древнейших переселений народов. И хотя находка корней и ранних ветвей линий Манью и Насрин на побережье Южной Аравии и Белуджистана является убедительным аргументом в пользу гипотезы о том, что один-единственный исход из Африки по южному маршруту прежде всего привел мигрантов в Южную Азию, она не обязательно является свидетельством того, что именно там появились на свет сама Насрин и ее шесть западных «дочерей», в том числе Рохани, или западно-евразийские «дочери» самой Рохани (U, HV и JT) (рис. 4.2). Хотя Рохани, вне всякого сомнения, появилась на свет где-то в Южной Азии (см. ниже), место рождения ее западно-евразийских дочерей менее ясно и представляет огромный интерес. Однако, если мы рассмотрим генетическую структуру митохондриевой ДНК во всем Ближневосточном регионе (включая Левант, Анатолию, Армению, Азербайджан, Северный Курдистан, а также более южные районы. Йемен, Саудовскую Аравию, Ирак и Иран), приводимую в анализе линий-родоначальниц современной Европы, мы без труда заметим огромное генетическое многообразие линий западных дочерей Рохани в Ираке — государстве на побережье Персидского залива (см. главу 3). Для Ирака характерно большое количество неклассифицированных генетических типов и отсутствие других западных дочерних групп Насрин, таких, как W, I или X. Таким образом, вполне возможно, что западные «дочери» Насрин появились на свет далеко на юге, в Месопотамии или возле побережья Персидского залива, и произошло это либо накануне первой, направленной на север, миграции в район Плодородного Полумесяца, либо во время нее.
Этот вопрос можно решить только посредством формального анализа линий-родоначальниц, такого, которым воспользовались Мартин Ричардс и его коллеги при отслеживании линий предков европейцев (см. главу 3), но в данном случае необходимо сопоставить Южную Азию с Левантом. На мой взгляд, для того чтобы индийская внучка Рохани, U2i, могла иметь такой же возраст (около 50 тысяч лет), как и ее европейская сестра, линия U5, проще всего предположить, что Рохани и ее «дочери» (Европа, HV и JT) появились на свет южнее, в районе Персидского залива, на полпути между Индией и Европой(см. рис. 4.2). Понятно, что, как мы уже говорили в главе 3, имеется веский аргумент в пользу того, что несколько ранних неафриканских Y-хромосомных групп возникли, сформировались именно в Южной Азии, а не на Леванте.
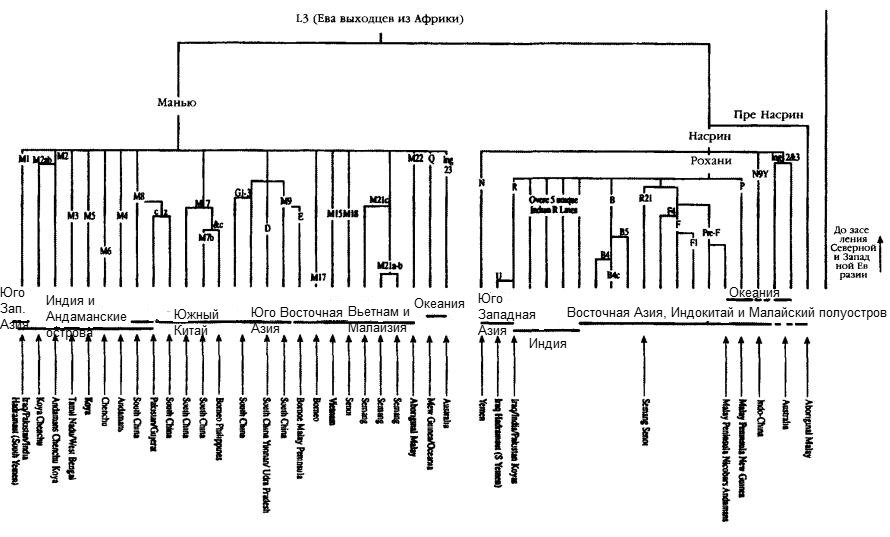
Рис. 4.2
Южная Азия: исток всех азиатских линий?
Хотя проблеме генетических маркеров на юге Аравийского полуострова посвящено не слишком много публикаций, по этой же тематике применительно к Индийскому субконтиненту существует обширный массив самой разной информации. Большую работу по исследованию митохондриевой ДНК провели эстонские генетики, которым, а также их коллегам из стран Азии мы выражаем глубокую признательность за труд. Ряд групп европейских и американских генетиков ведут активные исследования в области изучения Y-хромосом. И хотя их разработки в известной мере ставят под вопрос наш интерес к Пакистану и Индии, мы все же считаем, что корень проблемы следует искать именно там. Пакистан — это древнейшая прародина индусов, основной путь, ведущий из Южной Азии в Центральную, в регион, лежащий к западу от Гималаев. По территории Индии, Бангладеш и Бирмы также протекают великие реки Азии — Ганг, Брахмапутра и Салуин, поящие влагой обширные районы к югу и востоку от Гималаев.
Индийский субконтинент, где отмечается широкое многообразие типов внешнего облика и культур населяющих его народов, представляет собой богатейшую этническую и географическую «транзитную зону» между Западной и Восточной Евразией. К северу и западу от нее, по мере продвижения из Индии в Пакистан, из Пакистана в Афганистан и Иран и далее в Европу, мы наблюдаем плавные и постепенные изменения внешнего облика коренных жителей, включая и пигментацию (цвет) кожи (см. главу 5). Внешнее сходство между европейцами и многими народами Южной Азии просто поразительно.
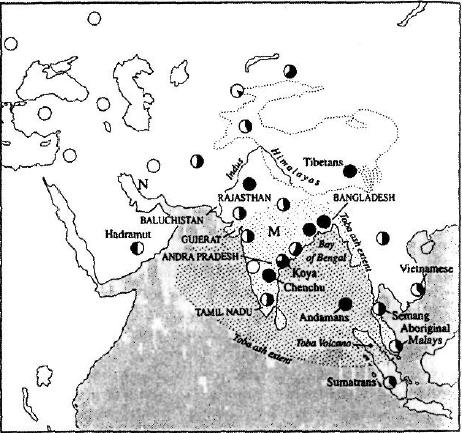
Рис. 4.3
Этот процесс постепенного изменения физического облика у жителей Северной Индии и Пакистана являет собой как бы параллель относительной частоте встречаемости двух генетических дочерних суперкланов, Насрин и Манью, на которые делится линия Евы выходцев из Африки (L3). (См. рис. 4.3.) Если обратиться к рассмотрению генофонда народа хадрамаут, живущего в Йемене, мы увидим, что соотношение встречаемости линий Насрин и Манью у них составляет 5 к 1, что хорошо согласуется с точкой зрения о том, что ветвь Насрин возникла далее к востоку, чем Манью, примерно в районе Персидского залива. В районе между Красным морем и устьем Инда в Пакистане западноевразийские генетические потомки Насрин продолжают превосходить в численности представителей второго азиатского суперклана — Манью.
Миновав устье Инда, оказавшись в Белуджистане и приближаясь к самой Индии, мы увидим, что картина начинает быстро меняться. Соотношение линий Насрин и Манью здесь, на западе Индии, в штатах Раджастан и Гуджарат, снижается до 2 к 1. А когда мы достигнем Бенгали и проникнем в Бангладеш, это соотношение изменится на противоположное и клан Манью будет доминировать там в пропорции 5 к 1. Таким образом, мы установили, что доминирующим кланом в Индии является Манью, причем его численность по мере продвижения с запада на восток постоянно возрастала.
Что касается районов на севере и востоке Индии, то там такие изменения выражены еще более резко. В Непале, Бирме и восточной Индии уже можно встретить первые лица монголоидного типа. Жители этих мест в основном говорят на языках Восточной Азии, что составляет резкий контраст с их ближайшими соседями, говорящими на индо-арийских или дравидийских языках. К тому времени, как мы достигнем территории восточной Бирмы и окажемся в Тибете, лежащем у северных склонов Гималаев, переход внешнего облика и этнолингвистического типа жителей уже можно считать завершенным, поскольку здесь имеет место быстрое и полное изменение соотношения представителей митохондриевых субкланов Манью и Насрин. В частности, на Тибете соотношение представителей Манью и Насрин составляет 3 к 1, и у нас нет достоверных данных о присутствии этих субкланов в Индии. На Тибете выявлено примерно 70% субкланов Манью и Насрин, типичных для Европы и Юго-Восточной Азии, а остальное приходится на долю неклассифицированных типов Манью местного происхождения. Таким образом северо-восточная часть Индийского субконтинента демонстрирует наиболее четкую и резко выраженную границу между востоком и западом. Эта граница, вполне возможно, отражает глубокую генетическую борозду, проходящую через всю Индию и связанную с облаком вулканического пепла, накрывшим Индийский субконтинент в результате извержения вулкана Тоба около 74 тысяч лет тому назад.
На юге Индийского субконтинента основной тип внешнего облика меняется, и преобладающим становится так называемый дравидский тип, для которого характерны курчавые волосы, очень смуглая кожа и круглые глаза (см главу 5). Сравнение формы черепа указывает на связь между тамилами, крупной этнической группой юга Индии, и представителями народности сеной, туземной группой живущей на Малайском полуострове и занимающей в этническом отношении промежуточное положение между народностью семанг и аборигенами Малаи (см. выше).
Ветвь Манью возникла в Индии, а Насрин — несколько западнее, в районе Персидского залива
Ветвь Манью, которая практически полностью отсутствует в Западной Евразии, дает немало оснований утверждать, что ее древней прародиной является Индия. Дело в том, что именно там, в Индии, Манью имеет наибольшее многообразие и древность. Более нигде в мире она не обладает таким множеством вариаций и обилием основных ветвей. Возраст древнейшей из ее «дочерей» в Индии, ветви М2, составляет около 73 тысяч лет. И хотя эта датировка времени распространения М2 не слишком точна, она вполне может отражать местное восстановление населения после гибели в результате катастрофического извержения вулкана Тоба около 74 тысяч лет тому назад. Ветвь М2 широко представлена у ченчу, народа охотников и собирателей, принадлежащего к австралоидной группе и живущего ныне в штате Андхра-Прадеш. Ченчу обладают целым рядом уникальных местных вариантов ветви М2, а также имеют общего предка с типами М2, выявленными в других районах Индии. В целом есть все основания, чтобы считать местом возникновения Манью именно Индию, а не какой-либо другой район, находящийся к западу от нее или в Африке.
Спектр типов Насрин, выявленных в Индии, также резко отличается от типов, встречающихся к западу от нее. Хотя типы, характерные для ее «дочери» Европы, выявлены у 13% индийцев, у которых были взяты пробы ДНК, практически все они приходятся на долю двух подветвей Европы, U7 и U2i (локальных индийских версий клана U2). Наряду с рассеянием других кланов Европы, выявленным к западу от Индии, там почти нет коренных типов Насрин. Это свидетельствует о том, что хотя U7 и U2i являются очень древними кланами в Южной Азии, они проникли туда с запада, во времена первой волны заселения Европы из Южной Азии, около 50 тысяч лет тому назад, то есть явно после первоначального исхода из Африки. У нас также нет данных, которые позволили бы поместить локус возникновения Насрин в Индии, хотя там и наблюдается рассеяние первого поколения ее «дочерей», которые имеют двойников в Западной Евразии.
Самой удивительной и любопытной среди всех родичей клана Насрин в Индии является ее «дочь», Рохани. Если читатель помнит, Рохани — самая плодовитая «дочь» Насрин, ставшая праматерью большинства людей Запада, включая Европу, не говоря уже о двух ее «дочерях» на Дальнем Востоке, каждая из которых тоже обладает обширным и равзветвленным потомством. При анализе присутствия Рохани в Индии создается впечатление, что ее плодовитость напоминает планирование семьи. Эстонские генетики сумели идентифицировать многочисленные ветви потомков Рохани, берущие начало в Индии, причем ни одна из них не встречается ни в каком другом регионе. Эти новые ветви настолько богаты, что удалось даже датировать время их распространения — около 73 тысяч лет тому назад.
Нигде более, ни на западе, ни на востоке, мы не находим столь широко разветвленного многообразия генетической линии Рохани. Одного этого вполне достаточно, чтобы считать Южную Азию исходной прародиной Рохани. Казалось бы, более логично предположить, что Рохани и ее европейские дочери появились на свет именно в районе Персидского залива, а не в Индии, но у нас нет никаких генетических оснований для подобного взгляда. Принимая во внимание сравнительно небольшую удаленность и огромную дистанцию во времени, можно признать, что возможен любой сценарий развития событий. И все же представляется крайне маловероятным, что Рохани могла появиться на свет на Леванте или в Европе. Дело в том, что в этих регионах не обнаружено никаких следов Рохани, а это аргумент в пользу версии о том, что врата для любой миграции на север из районов, прилегающих к Сахаре, в ту эпоху были уже закрыты.
Но, пожалуй, самое интересное здесь заключается в том, что невиданно пышный расцвет линий кланов Манью и Насрин в Индии говорит о том, что они увековечили генетическую память о людях, которым посчастливилось выжить во время катастрофического извержения вулкана Тоба около 74 тысяч лет тому назад. Обезлюдевшая Индия, вполне возможно, начала заселяться с запада представителями разных типов клана Рохани, а с востока — представителями ветвей Манью. Дополнительными аргументами в поддержку этой гипотезы являются результаты недавнего исследования Т. Кивисилда и его коллег, изучавших две племенные группы на юго-востоке штата Андхра-Прадеш. Одна из этих групп, охотники-собиратели племени ченчу, относящиеся к австралоидному типу, почти целиком принадлежит к клану Манью и сохраняет некоторые ветви Манью, как характерные для Индии, так и и уникальные для нее. Другая этническая группа, народность койа, не принадлежащая к австралоидному типу, сохранила столь же богатый спектр индийских ветвей Манью (60% от общего числа линий) и, кроме того, имеет 31% типов Рохани. Племенные группы ченчу и койа представляют собой как бы древнюю библиотеку генетических линий Рохани и Манью в Индии, которые являются предками большинства материнских генетических линий из всего их многообразия, отмечаемого на остальной территории Индийского субконтинента. Ни у одного из этих типов не выявлено параллельное присутствие западно-евразийских типов Насрин. Наличие типов Рохани у народности койа, а не у австралоидов ченчу, возможно, объясняется некоторыми аспектами повторного заселения Индийского субконтинента с запада. Любопытно, что у этих двух племенных групп, несмотря на всю их древность и независимую друг от друга эволюцию, а также несомненно индийские генетические корни и локальный характер, нет общих материнских генетических типов (т. е. нет общих точек соприкосновения).
Первые люди Азии: три источника
Можем ли мы проследить маршрут и восстановить историю миграции линий-родоначальников Адама выходцев из Африки вдоль побережья Аравийского полуострова и далее через Пакистан в Индию? Задача была бы куда легче, если бы на пути миграции остались представители Адама выходцев из Африки, но корневая линия Адама, как и Евы выходцев из Африки, полностью вымерла, и у него остались лишь три непосредственных «сына» — Каин, Авель и Сиф, три мужские линии, которые охватывают все остальное человечество за пределами Африки.
Одна из этих трех корневых ветвей, С (или RPS4Y), которой я дал название Каин — по имени первородного сына Адама, представляет ветвь, встречающуюся только у неафриканского населения и в небольшом количестве присутствующую у коренных жителей Южной и Восточной Азии, Океании, а также Северной и Южной Америки. С еще более низкой (около 5%) встречаемостью линия Каин присутствует в разных районах Индии, включая и наиболее австралоидное из ее этнических меньшинств — племя ченчу. Линия Сифа, F (или М89), также является практически полностью неафриканской, за исключение разве что нескольких корневых типов, которые, как и ветви материнской линии Манью (M1), простираются вплоть до Северной Африки и, в частности, присутствуют в Марокко. Сиф — бесспорно, самая распространенная из этих ветвей; она встречается во всем мире за пределами Африки, причем ее встречаемость особенно высока в Австралии и Новой Гвинее, что со всей определенностью демонстрирует, что она представляет собой соединение двух генетических сыновей Адама на пути их эпических странствий по прибрежной полосе. На долю Сифа приходится около 95—97% всех мужских линий Индии, и он образует там множество ветвей.
Три наиболее распространенные в Индии ветви не встречаются практически более нигде в мире (единственное исключение — некоторые районы Центральной Азии к северу от Индии) и, таким образом, словно эхо отражают это пышное цветение материнских линий в Индии. Две из них представляют собой линии потомков группы К, Кришны, наиболее плодовитого сына Сифа, тогда как третья — это ветвь первого поколения, идущая от Сифа и встречающаяся не только в упомянутых районах Центральной Азии, но и, что особенно важно, далеко на юговостоке, в Камбодже. Все эти типы активно представлены у племен и этнических групп Южной Индии, особенно — у австралоидных ченчу. Люди племени койа, кроме того, сохранили и редкий корневой тип Сифа, встречающийся у четверти койа. Этот тип ограничен Индийским субконтинентом и соседними районами Центральной Азии, что является аргументом в поддержку версии о том, что именно Индия явилась для Сифа важнейшей «опорной базой» на его южном пути из Африки.
Согласно другой концепции исхода из Африки, полученной на основе данных по Y-хромосоме, в первоначальной группе мигрантов-собирателей на прибрежной полосе продвижения присутствовали только линии Каин и Авель, Сиф же появился много позднее, пройдя через Северную Африку и Левант и направившись в Центральную Азию, а уже оттуда — в Индию и Юго-Восточную Азию. Как сказано в этой и следующей главах нашей книги, я не разделяю подобную точку зрения, доказывая посредством целого ряду аргументов (включая близкое географическое и генетическое совпадение линий-родоначальниц), что объяснить все имеющиеся находки лучше всего позволяет версию об одном-единственном, южном, маршруте исхода. Что же касается присутствия линии Сифа в Центральной Азии, то наиболее распространенный индийский тип Y-хромосом является ключом к решению вопроса о том, берут ли многочисленные Y-хромосомные линии, присутствующие в Индии и Центральной Азии, свое начало в Индии или наоборот. Например, тип M17, который я уже упоминал в главе 3 как одного из главных фигурантов в Европе, пришел именно из Центральной Азии. Кроме того, M17 имеет в Индии среднюю встречаемость порядка 27%, но достигает в одном из ее штатов, штате Пенджаб, уровня 47%. Наибольшего многообразия — куда более широкого, чем в Центральной Азии или Европе, — М17 достигает в Иране, Индии и Пакистане. Это относится и к Южной Азии, которая является его древней прародиной. Ключевой вопрос здесь — поиск М17 при высокой встречаемости и широком многообразии у представителей племенных групп Южной Индии, в частности у австралоидных племен ченчу и ядхава. Если бы M17 у представителей этих племенных групп была привнесенной, а не коренной линией, ее многообразие было бы значительно более низким.
Вопрос о противоречивости линии YAP: африканская она или азиатская?
Третьей основной линии, часто называемой YAP, я дал имя Авель — по имени второго сына Адама. Одна из ветвей этой линии — небольшая, глубокая и очень древняя азиатская ветвь, распространение которой носит очаговый, дискретный характер, в основном — на Дальнем Востоке (более подробно о ней см. ниже). Другая, западная, ветвь YAP образует несколько ответвлений, представители которых заселили многие районы Северной Африки и Среднего Востока, хотя в небольшом количестве она присутствует у жителей Пакистана, Индии и Центральной Азии. Некоторые подветви YAP относятся к числу наиболее распространенных в Африке, тогда как другие характерны только для Среднего Востока.
Здесь можно вспомнить аналогию с Библией: в первой ее книге, книге Бытия, сказано, что Сим и Хам были двумя братьями, потомки которых заселили соответственно Средний Восток и Северную Африку. К большому сожалению для наших изысканий, первоначальная мужская линия Авеля, возникшая еще до разделения YAP+ на западную и восточную ветви, по всей вероятности, не сохранилась и полностью вымерла, так что от нее остались лишь два «сына». Все, что оставила нам эта ветвь, — это тайну своего происхождения. Где же она возникла: в Эфиопии или Аравии? Вопрос, вынесенный в заглавие параграфа «африканская или азиатская?» присутствует и в названии «хамито-семитская», используемом для характеристики языковой семьи, на языках которой говорит подавляющее большинство потомков YAP. Но, независимо от того, какое конкретно название мы используем применительно к ней — старое, восходящее к Библии, или же более новое, «афроазиатская», — главный вопрос сводится к тому, являются ли корни происхождения этой языковой семьи африканскими или азиатскими. Учитывая сравнительно молодой возраст большинства крупных языковых семей, можно предположить, что любая миграция через Красное море, будь то с востока на запад или с запада на восток, по-видимому, имела место около 10 тысяч лет тому назад.
Известный американский генетик Майк Хаммер настаивал, что линия YAP возникла и сложилась за пределами Африки и что один из ее «сыновей» впоследствии возвратился на древнюю прародину. Другая группа генетиков во главе с Питером Андерхиллом отстаивает версию об африканском происхождении YAP. В этом вопросе я склонен согласиться с Майком Хаммером и утверждать, что линия YAP присутствовала в первой колонии выходцев из Африки, однако неясность происхождения афроазиатской языковой семьи заключает в себе нечто большее, чем простую аналогию (см. рис. 4.4). Неафриканские генетические линии, которые восходят к Эфиопии и Северной Африке, вполне могли сложиться и после последнего ледникового максимума, когда люди начали активно возвращаться на зеленые пространства, еще недавно бывшие пустынями.
Большой интерес представляет вопрос о том, когда именно после последнего оледенения имело место это расселение. Некоторые археологи и лингвисты считают, что заметное увеличение крупнейших языковых семей было связано с сельскохозяйственной революцией эпохи неолита, имевшей место в последние 5000—7000 лет. Однако генетические данные показывают, что повторное расселение имело место в эпоху мезолита (т. е. примерно 12 500 лет тому назад), то есть, другими словами, гораздо раньше эпохи неолита.
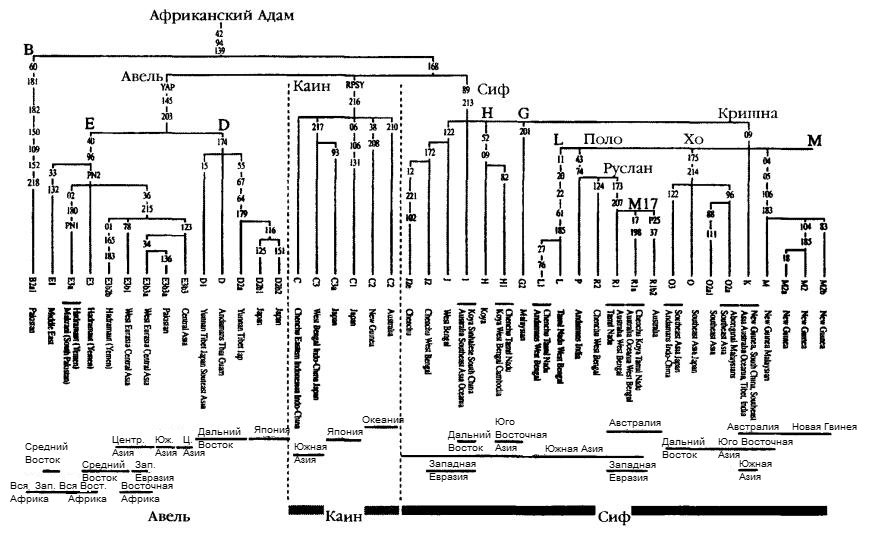
Рис. 4.4
Загадочным аспектом миграции линии Авеля является зияющая пауза в ее расселении между Западной Евразией и Дальним Востоком и, что особенно важно, ее практически полное отсутствие в Индии. Тот факт, что она тоже присутствовала в группах мигрантов-собирателей, продвигавшихся по прибрежной полосе, заметен благодаря присутствию азиатской YAP у аборигенов Андаманских островов, Камбоджи и Японии (см. главу 5). Данные недавних исследований свидетельствуют о присутствии азиатской ветви YAP на уровне встречаемости 3—6% и такой же частоты на западе, далеко к северу от прибрежной полосы, в частности в Центральной Азии (Узбекистан). Это отражает картину расселения линии Каин, которая хотя и не полностью отсутствует в Индии, тем не менее имеет там гораздо более низкую встречаемость, чем на севере, в Центральной Азии. Учитывая, что азиатская ветвь YAP хотя и редко, но все же встречается по древнему пути миграции собирателей на прибрежной полосе и рядом с ним, вполне возможно, что ее отсутствие в Индии — а также низкую встречаемость линии Каин в Южной Азии — можно объяснить опустошительным влиянием катастрофического извержения вулкана Тоба на весь Индийский субконтинент, — извержения, о котором свидетельствуют данные геологии и генетической истории материнских линий (см. выше). Извержение Тоба могло создать генетическую перемычку, за которой последовало активное местное восстановление линии Сифа за счет двух братьев-собирателей на прибрежной полосе — Каина и Авеля.
Три азиатские мужские линии-родоначальницы: все они были собирателями на прибрежной полосе
Давайте возвратимся к трем основным мужским линиям выходцев из Африки. Можем ли мы задать такой же вопрос, который обращали к нуклеарным и митохондриевым маркерам относительно следов древнейшего исхода из Африки? Я думаю — да. Датировка Y-ветвей пока что находится в зачаточном состоянии, однако, если мы обратимся к географическим локусам стоянок предков и потомков, мы сможем оценить географическую последовательность распределения ветвей. Это — так называемый филологогеографический метод. Для начала возьмем линию Каин. Поскольку на ее долю приходится 60% всех типов Y-хромосом в Австралии, она является там доминирующей линией. На таких островах Восточной Индонезии, как Молуккские острова и о. Нуса-Тенггара, которые первыми встретились мигрантам на их пути в Австралию, мы видим наименее модифицированные или даже корневые типы линии Каин, от которой происходит уникальный аборигенный австралийский тип. Единственным местом в мире, где также выявлен этот корневой тип, является Индия, где он имеет довольно низкую встречаемость, в том числе и у представителей туземных племен австралоидного типа. На соседнем острове Новая Гвинея вы встречаем линию С2, близко родственную («братскую») австралийской линии Каин. Взглянув на локусы других древнейших колоний, мы без труда заметим, что Каин практически отсутствует в Западной Евразии, но зато широко распространен к востоку и северу от Индии. Резкое падение встречаемости линии Каин показывает, что она по большей части присутствует на восточно-азиатском побережье Тихого океана и гораздо реже — в глубинных континентальных районах Центральной, Северной и Южной Азии. Эти азиатские типы по большей части принадлежат к одному клану, который впоследствии мигрировал в Америку, явившись родоначальником тамошних аборигенов. Широкое многообразие линий этого клана отмечено в Японии.
Распространение линии Каина в прибрежных районах Восточной Азии и выраженные региональные различия у ее потомков, наблюдаемые в Юго-Восточной Азии и Океании (рис. 4.5), свидетельствуют о том, что эта линия была в числе родоначальниц мужских линий на пути первоначальной миграции собирателей по всему южному побережью Азии, хотя ее и нельзя считать единственной, поскольку обе линии-«брата» также оставили следы своего продвижения на побережье Индийского океана в ЮгоВосточную Азию и Японию. Однако длинный путь в Австралию, а также к побережью Восточной Азии преодолела только одна из двух оставшихся ветвей — линия Сиф. Ее следы выявлены благодаря присутствию (правда, достаточно редкого) линии Сиф в ее первоначальном или корневом виде во всех указанных регионах. В Австралии выявлены два основных локальных типа Y-хромосом: один из них происходит от линии Каин (см. выше) и является наиболее распространенным, поскольку его численность составляет 60% от общего числа линий, а другой — от азиатского генетического «сына» Сифа, Кришны, на долю которого приходится до 30% оставшихся линий. Такой уровень встречаемости Кришны (около 30%) сохраняется более или менее стабильно на всем побережье Юго-Восточной Азии и западного побережья Тихого океана вплоть до Кореи. В то же время линия потомков Сифа присутствует на уровне нескольких процентов в Юго-Восточной Азии и Австралии, а также на тихоокеанском побережье, в Корее. Эти данные свидетельствуют о том, что первый маршрут собирателей на прибрежной полосе пролегал не только в Новую Гвинею и Австралию, но и продолжался далее, по всему тихоокеанскому побережью Китая, Японии и Кореи. Третья корневая линия Y-хромосомы, азиатская YAP, доходит до Камбоджи, а затем круто сворачивает в сторону Австралии, вместо того чтобы продолжать путь на север, вокруг побережья Тихого и Индийского океанов. Она имеет эпизодическую встречаемость в Юго-Восточной Азии и очень высокую — на Тибете, в южном Китае и Японии.
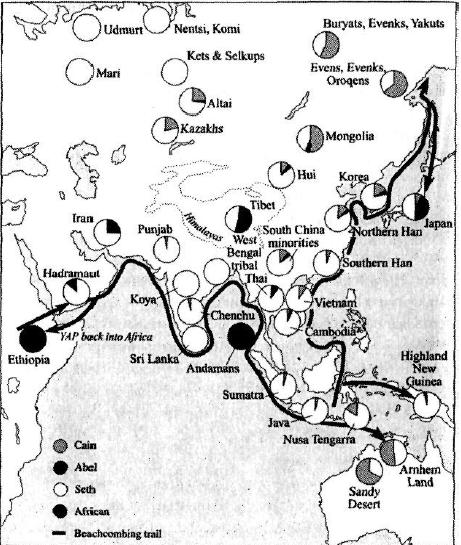
Puc. 4.5
Подобная картина распространения мужских генетических линий служит зеркальным отражением миграций материнской мтДНК и очень много, хотя и в косвенной форме, говорит нам об идентификации и возрасте первых мужских линий-родоначальниц. Согласно прогнозам, южный маршрут имел место где-то в самом начале пути, и прежде чем приступать к его отслеживанию, мы должны попытаться найти точку, в которой сходятся все три древнейшие неафриканские линии. В настоящее время ведутся исследования на юге Аравийского полуострова и в районе Персидского залива, но если мы обратимся к рассмотрению этих трех мужских линий-родоначальниц в Пакистане и Индии, мы без труда установим, что там представлены все три эти линии. Мы можем найти и корень, и ветви линий Каин, Сиф и генетических «сыновей» и «внуков» Сифа. Более того, мы найдем и ряд уникальных типов YAP от ветви YAP, которая обычно встречается только в Африке и Западной Евразии.
Нигде более за пределами Африки мы не встречаем такого широкого многообразия корней и ветвей Y, если не считать регионов Центральной и Северной Азии, да и там они, надо признать, представлены далеко не столь пышно и обильно. Эта картина, характерная для Центральной Азии как промежуточной зоны между Востоком и Западом, обусловлена активным смешением европейских и азиатских материнских линий мтДНК, представленных в этом регионе, что свидетельствует о том, что одним из самых первых этапов разделения после прибытия в Индию был маршрут на север, вверх по течению Инда, ведущий в Центральную Азию. Этот ранний маршрут, устремленный в глубь континента, более подробно рассмотрен в следующей главе.
Таким образом, весь регион Южной Азии, древнейшей прародины участников одного-единственного и успешного исхода по южному маршруту, свидетельствует о присутствии в нем генетических корней, связанных с миграциями не только так называемых туземных этнических групп вдоль побережья Индийского океана, но и многих предков современных народов. Среди этих корней мы можем выявить базовые генетические «лагеря» и опорные пункты большинства последующих миграций групп и племен в глубь континента, по необъятным просторам Евразии. Эти маршруты миграций после небольшой паузы были направлены в Европу, Кавказский регион и Центральную Азию. Создается впечатление, что передовые отряды этих мигрантов по прибрежной полосе сохранили удивительное первозданное генетическое многообразие, характерное группе выходцев из Африки, и двинулись в путь по побережью Индийского океана. Таким путем они довольно скоро достигли Индонезии и островов западной Океании, проникнув в Австралию задолго до того, как их кузины и кузены, отправившиеся на запад, оказались в Европе.
Точная хронологическая взаимосвязь между исходом из Африки и проникновением в Юго-Восточную Азию, с одной стороны, и катастрофическим извержением вулкана Тоба около 74 тысяч лет тому назад, с другой, имеет критически важное значение. Прежде всего, извержение Тоба — это одно из наиболее точно и достоверно датируемых событий эпохи палеолита, и выпадение его вулканического пепла можно считать точным критерием датировки для всей Южной Азии. Во-вторых, пагубный эффект вулканического пепла и последовавшая за ним долгая «ядерная зима» неизбежно должны были оказаться губительными для всего живого на пути выпадения пепла, а фактически захватили куда более обширный регион. Наличие в вулканическом пепле Тоба орудий, созданных, как считается, людьми современного типа на стоянках на Малайском полуострове, свидетельствует, что передовые отряды мигрантов-собирателей на прибрежной полосе проникли на Дальний Восток задолго до извержения Тоба. Триангуляция этой рубежной даты и других дошедших до нас фактов говорит в пользу такого сценария. В числе других «ключей» — новые данные по изучению люйцзяньского черепа, данные датировки по люминесцентному методу объектов в Австралии и оценки минимального уровня Мирового океана, позволявшего совершить миграцию в Австралию около 70 тысяч лет тому назад практически сухопутным путем, генетические данные о расселении группы L3 около 83 тысяч лет тому назад и, наконец, период резкого повышения солености Красного моря, начавшийся около 80 тысяч лет тому назад. Наиболее убедительными свидетельствами очень раннего присутствия человека современного типа в Азии могут стать находки ископаемых останков и исследование всего контекста их культурного слоя. Такие работы активно ведутся на стоянке Лианг Буа на о. Флорес.
Таким образом, если взрыв вулкана Тоба действительно произошел уже после того, как Индия была заселена людьми, мы вправе ожидать массовой гибели населения на всем Индийском полуострове в результате извержения, затронувшего восточную часть субконтинента значительно сильнее, чем западную. Это всего лишь одно из возможных объяснений парадокса генетической картины Индии, где можно выявить генетический след древнейших мигрантов-собирателей на прибрежной полосе. В то же время множество индийских подгрупп линий Манью и Рохани являются уникальными и ограничиваются только самим субконтинентом; в особенности это относится к племенам на юго-востоке полуострова. А это — именно та картина, которую мы вправе ожидать после грандиозной катастрофы. Возраст наиболее древних из этих локальных линий достигает 73 тысяч лет.
В следующей главе мы рассмотрим вопрос о том, как повели себя эти первопроходцы на севере и востоке азиатского континента и каким образом они проникли в эти места.

