Книга: Двигатели жизни. Как бактерии сделали наш мир обитаемым
Назад: Глава 6. Защита важнейших генов
Дальше: Глава 8. Крупномеры страны чудес
Глава 7. Сокамерники
Одна из стратегий, используемых природой, чтобы удостовериться в том, что ее интеллектуальная собственность достаточно устойчива перед лицом потенциальных глобальных катастроф, состоит в распространении риска среди широкого круга микроорганизмов. Инструкции для наномеханизмов распространяются при помощи горизонтального переноса генов. И хотя горизонтальный перенос генов является для микроорганизмов основным эволюционным режимом, этот процесс не полностью случаен и лишен системы. Один из основных его двигателей – экологический: это симбиотическое объединение микроорганизмов для оптимизации потребления скудных питательных веществ. Этот двигатель хорошо послужил эволюции жизни на планете.
Микроорганизмы не живут в изоляции, большинство из них – симбионты, то есть они живут вместе и зависят друг от друга в добывании ресурсов. Говоря более конкретно, микроорганизмы используют в пищу продукты жизнедеятельности друг друга. Такое использование продуктов жизнедеятельности, также называемое рециркуляцией (ресайклингом) элементов, является одной из основных концепций в экологии, и оно сильно повлияло на эволюцию микробиологических наномеханизмов. У микробиологов ушло немало времени на то, чтобы оценить взаимодействия своих предметов наблюдения в глобальном масштабе, но в конечном счете такая оценка привела к гораздо лучшему пониманию эволюции жизни на Земле.
В течение десятилетий основной подход, использовавшийся микробиологами для изучения микроорганизмов, состоял в том, чтобы изолировать единичные клетки от их среды и пытаться вырастить их как чистую культуру. Такие клоны – колонии клеток, произведенные от одной материнской клетки, считались золотым стандартом, и их применение было утверждено Кохом в числе четырех принципов, доказывающих, что конкретный организм ответствен за конкретное заболевание. Такой подход не лишен ценности. Зачастую очень незначительные вариации внутри популяции одного вида микроорганизмов ведут к большим изменениям в способности клона вызывать заболевание. Классический пример – пищевое отравление, вызываемое распространенной бактерией Escherichia coli, имеющейся в кишечнике у любого из нас.
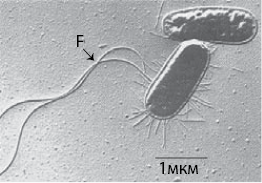
Рис. 26. Электронная микрофотография бактерии Escherichia coli – возможно, наиболее изученного микроорганизма в биологии. Она находится в кишечнике человека, однако патогенные штаммы (с виду идентичные непатогенным) зачастую вызывают у людей пищевые отравления. Эти организмы имеют жгутики, позволяющие им плавать в жидкой среде
E. coli – вероятно, наиболее изученный в биологии организм. Ее очень легко выращивать, она широко распространена, благодаря чему стала образцовой моделью для микробиологов-генетиков. Небольшие изменения в генах этого организма, передающиеся с пищей, могут вести к обширным и порой очень тяжелым вспышкам кишечных заболеваний и даже к смерти людей. В таком контексте изучение пищевых запросов клона, скорости его роста, чувствительности или сопротивляемости к антибиотикам и так далее становится вопросом первостепенной важности. Однако при широком применении генного секвенирования быстро стало очевидным, что неболезнетворные штаммы E. coli могут очень быстро стать патогенными и вызывать обширные внутренние кровоизлияния при попадании внутрь человеческого организма. Они приобретают патогенные гены от других штаммов посредством горизонтального переноса генов, путем конъюгации – аналога полового акта у микроорганизмов, который в данном случае позволяет болезнетворному штамму распространять гены, вызывающие заболевания у людей. Для того чтобы сделать E. coli патогенной, требуется лишь небольшое число генов. Патогенные штаммы отделились от неболезнетворных около четырех миллионов лет назад, однако до наступления эпохи генного секвенирования было очень сложно точно установить различия между этими двумя штаммами самого изученного из микроорганизмов. Если мы не можем отличить друг от друга два штамма E. coli, не изолируя их в чистой культуре и не секвенируя их геномы, как мы собираемся понять жизнь микроорганизмов в окружающем нас мире?
Более 99 % микроорганизмов в океанах, почвах, на поверхности горных пород и даже в наших собственных кишках, идентифицированные посредством генного секвенирования, не были изолированы и не выращивались в лабораторных культурах. Предпринималось множество попыток изолировать триллионы микроорганизмов из океанов, почв, гидротермальных источников на океаническом дне, из наших кишечников и ротовых полостей, а также множества других сред. Иногда эти попытки были удачными, и микроорганизм удавалось уговорить расти в чистой культуре, но чаще всего они проваливались. Долгое время считалось, а зачастую считается и сейчас, что причина нашей неспособности изолировать микроорганизмы как чистые клонированные культуры состоит в том, что ученые попросту не знают, какие питательные вещества требуются этим предположительно хрупким конкретным организмам для их роста. Сколько сахаров и какого типа, какие аминокислоты и сколько соли необходимо для каждого конкретного вида? Число комбинаций практически бесконечно. В этом отношении у людей нет почти никакого представления о том, как функционируют микроорганизмы. И поэтому в лабораториях, где цель обычно состоит в том, чтобы заставить множество микроорганизмов расти как можно быстрее, им предоставляют в избытке сахара, аминокислоты и все остальное, что нужно, чтобы убедить их расти. Концентрации питательных веществ в лабораторном бульоне чаще всего в тысячи раз превышают то, что можно наблюдать в реальном мире. Сахара, аминокислоты и другие питательные вещества, за очень небольшими исключениями, имеются в природе в очень скудных количествах, и у микроорганизмов уходит много энергии на то, чтобы их добыть. Понимание того, как микроорганизмы добывают себе пропитание в реальном мире, потребовало нового подхода. Микробиологи-экологи стали по сути социологами, изучающими взаимодействия между микроскопическими организмами.
Для того чтобы минимизировать затраты энергии, уходящей на добывание питательных веществ, микроорганизмы в природе стремятся образовывать сообщества, в которых, к примеру, сахар, выделяемый одним организмом, потребляется другим, в то время как сам потребитель сахара предоставляет другим членам сообщества аминокислоты. Получается, что в общем и целом микроорганизмы, как и мы сами, являются социальными существами. Если им не хватает сложных форм поведения, то они искупают это инновационным метаболизмом, в большой степени основанным на гибкости их наномеханизмов и их способности приспосабливаться к изменениям в окружающей среде.
Микробиотические сообщества, или консорции, представляют собой микроскопические джунгли, в которых десятки и даже сотни видов микроорганизмов обитают в пределах одного ареала. Следует отметить, что часто бывает трудно строго определить, что является «видом» для микроорганизма. Традиционное определение этого слова, исходящее из жизнеспособности потомка половой рекомбинации особей и легко проверяемое у животных и растений, для микроорганизмов чаще всего неприменимо. Дело не только в том, что у большинства микроорганизмов сложно определить пол – горизонтальный перенос генов делает само понятие «вида» несколько расплывчатым. Тем не менее для того чтобы лучше вникнуть в функционирование микробиотической консорции, мы будем говорить о «видах» микроорганизмов в контексте какой-либо наблюдаемой биологической функции, в первую очередь метаболизма. Представим себе, что один микроорганический вид выделяет в окружающую среду какой-либо секрет или газ, который другой вид может использовать как источник энергии. Затем второй вид выделяет собственные секреты и газы, которые могут быть возвращены первому виду или переданы дальше другим видам, или и то и другое одновременно. Результатом является возникновение микроскопического микробиотического сообщества, которое по сути представляет собой миниатюрный биологический рынок электронов.
Представление о рынке электронов в микробиотической консорции – не метафора. Микроорганизмы внутри консорции в буквальном смысле обмениваются газами и другими материалами, имеющими избыток или недостаток электронов. Например, и у метана, и у сероводорода избыток электронов. Эти восстановленные молекулы могут вырабатываться несколькими различными участниками консорции и выделяться в окружающую среду. Другие микроорганизмы используют такие богатые электронами молекулы как источник энергии. В свою очередь, продукты их секреции, например углекислый газ и сульфат, могут подвергнуться рециркуляции или же быть потеряны сообществом, уйдя во внешнюю среду. Микробиотические консорции могут сохранять стабильность на протяжении дней, декад или даже дольше; мы попросту не знаем этого, но ответ может скрываться в том, что написано выше. Тем не менее мы знаем кое-что об основных законах такой консорции.
Одно из правил микробиотической консорции состоит в том, что ни один ее член не может превзойти остальных вплоть до их исключения. Если эта установка будет нарушена, консорция развалится и «победивший» микроорганизм окажется в энергетическом проигрыше, ему придется закупать дефицитные питательные вещества на отдаленных рынках, вместо того чтобы жить и обедать, наслаждаясь роскошью прямой доставки продуктов местного производства прямо к своим микроорганическим дверям.
Означает ли это, что все микробы «играют честно»?
Микроорганизмы могут быть социальны, но также способны проявлять агрессию и стремиться победить. Часто они производят молекулы, убивающие другие микроорганизмы. Собственно, большая часть самых действенных антибиотиков, борющихся с инфекционными болезнями, производится микроорганизмами. Однако в контексте микробиотической консорции такие молекулы зачастую служат только для защиты от захватчиков и не применяются против членов самой консорции. Другими словами, чтобы нам было полностью понятно, существует джентльменское соглашение: определенные микробы с определенными функциями имеют допуск в клуб обедающих, в то время как все остальные из него исключены.
Эта гипотеза легко подвергается проверке. Люди рождаются без каких-либо микроорганизмов в своем кишечнике. Однако очень быстро мы приобретаем их из окружающей среды. Мы получаем их, когда, едва родившись, трогаем свою маму и сосем ее грудь; мы едим сырую пищу; мы едим грязь; мы даже можем попробовать на вкус свои какашки. Фактически одним из первых микроорганизмов, колонизирующих наш кишечник, является E. coli – будем надеяться, неболезнетворный штамм.
Со временем каждый из нас начинает выращивать в своем кишечнике микробиологический зоопарк, уникальный для каждого – возможно, даже более уникальный, нежели последовательность ДНК. Общее число микроорганизмов в кишечнике любого человека приблизительно в десять раз превышает общее количество клеток в его теле. Дело не только в том, что микрофлора нашего кишечника приспособлена к нашей персональной диете и окружению – состав консорции также чрезвычайно важен для нашего личного здоровья. Консорция помогает нам получать питательные вещества из пищи, способствуя расщеплению сложных углеводов и жиров, помогает производить для нас витамины, а также не дает «плохим» микроорганизмам вызывать у нас болезни, препятствуя их росту. Мы все так или иначе знаем об этом. Любой, кому случалось во время путешествия по незнакомой стране заболеть, попив воды из-под крана, удивлялся, почему все местные жители не умирают еще в детском возрасте. Фактически возможно, что для многих все тем и закончилось, но у выживших в кишечнике имелись микроорганизмы, защитившие их от болезнетворных микробов в воде, которую они пили. Вы же у себя дома не имели возможности получить из пищи или воды таких же защитников. Если бы вы прожили в чужой стране подольше или были там рождены, у вас бы они тоже были – в противном случае вас ждало бы истощение, смерть или по меньшей мере неудачи при попытках размножения.
В наше время часто случается так, что мы в какой-то момент нашей жизни заболеваем, и врач выписывает нам антибиотик или даже два. Мы принимаем курс антибиотиков, одним из побочных эффектов которого часто является гастроэнтерит – сопутствующий ущерб от приема антибиотиков состоит в гибели многих микроорганизмов в нашем кишечнике. Для нас это означает не просто плохое самочувствие; это также изменяет характер взаимодействия микроорганизмов в нашей кишечной консорции. Порой необходимо несколько месяцев для того, чтобы консорция вернулась к тому состоянию, в котором находилась до приема антибиотиков. У некоторых людей она может не восстановиться даже через год. В других случаях нам бывает сложно приспособиться, у кого-то возникает неадекватная реакция на пищу, которую мы привыкли употреблять, и это продолжается какое-то время после приема курса. Наши персональные взаимоотношения с нашими кишечными микроорганизмами, на которые в целом приходится около двух килограммов массы нашего тела, могут рассматриваться как микрокосм, миниатюрная модель того, что микроорганизмы делают в глобальном масштабе.
Консорции представляют собой микроскопический образ глобального электронного рынка; однако каждой конкретной группе микроорганизмов внутри консорции неизбежно не хватает одного или нескольких ключевых метаболических путей, чтобы соблюдать в группе энергетический баланс. Например, какая-либо группа может обладать способностью связывать азот, но эта функция может быть не востребована, если в консорции имеется избыток азота. Другая группа может связывать углерод, но этот элемент может не ограничивать рост консорции. Одной (а чаще нескольких) ключевых реакций всегда не хватает или же они не сбалансированы. Из этого следует, что кругооборот питательных веществ и газов внутри консорции всегда далек от совершенства, и консорция, чтобы сохранять жизнеспособность, принуждена постоянно копаться на электронном рынке.
Между микробиотической консорцией и окружающей ее средой всегда существует измеримый конечный обмен газами. Например, консорция может либо поглощать, либо производить кислород, углекислый газ, метан, сернистый газ, сероводород, азот или какой-либо другой газ. Фактически, отслеживая такой обмен газами с окружающей средой, часто можно определить, какого рода микроорганизмы составляют консорцию. Хотя консорции и обладают относительной независимостью, они неизменно выделяют газы во внешний мир. Эти газообразные отходы их жизнедеятельности затем распространяются через атмосферу или океаны, которые эффективно действуют как проводники, связывающие метаболизм микроорганизмов на всей поверхности планеты.
Давайте рассмотрим эту концепцию на локальном, персональном уровне – вновь заглянув в наш собственный кишечник. Не углубляясь в интимные подробности, скажем, что наши личные микробиотические консорции также явно не пребывают в сбалансированном состоянии. Большая часть нашего газообмена с внешним миром происходит через нос и рот. Тем не менее у нас имеется и другой путь обмена газами, и этот способ может очень много рассказать нам о наших микробиотических консорциях. Практически все газы, выходящие из анаэробных кишечников всех млекопитающих, являются окисленными – и здесь однозначно преобладают азот и углекислый газ. Однако некоторые из газов оказываются восстановленными, и из них сульфиды наиболее очевидны для нашей обонятельной системы. Два других восстановленных газа, не имеющие заметного запаха, – это метан и водород. Приблизительно у половины из нас в толстой кишке имеются метанообразующие бактерии, и почти все мы испускаем газообразный водород. Оба эти газа могут гореть. Все газы, произведенные микроорганизмами нашего кишечника, являются побочными продуктами метаболизма, не сбалансированного с окружающей средой. Если бы это было не так, газы были бы идентичны тем, что содержатся в планетарной атмосфере, а в данном случае ничего подобного не наблюдается. Если смесь газов в нашем кишечнике не находится в равновесии с атмосферой планеты, из этого следует, что совокупность всех консорций микроорганизмов в кишках всех животных не находится в равновесии с метаболическими путями на планете. Чтобы обмен электронами между триллионами микробиотических консорций был действенным в глобальном масштабе, необходимы какие-то глобальные же механизмы проверки и балансировки – ученые часто называют их метаболическими обратными связями.
За немногими исключениями изменения концентрации и состава газов в планетарной атмосфере по причине исключительно естественных процессов обычно не могут быть измерены по временной шкале столетий. Микроорганизмы создают глобальный рынок электронов, который стабилизируется благодаря интеграции метаболизма сотен миллиардов консорций, распространенных по всей планете – от поверхностных пленок на озерах до осадочной толщи и горных пород океанического дна на глубине сотен метров. Метаболизм Земли – это продукт консорции консорций, для которого индивидуальные консорции не имеют большого значения, но механизмы всех реакций передачи электронов распределяются неслучайным образом, в зависимости от наличия и доступности ресурсов. Природная система страхования состоит в распространении рисков, прежде всего, путем инвестирования в глобальный микробиологический электронный хеджевый фонд. Это инвестирование заключается в потенциальной возможности наномеханизмов работать, основываясь на доступности в окружающей среде любой молекулы, которая может послужить либо источником, либо потребителем электронов.
В микроскопическом масштабе организмы, составляющие консорцию, живут в очень тесной близости. В такой обстановке возможности для горизонтального переноса генов многократно увеличиваются. А значит, внутри консорции перенос генов зачастую допускает распространение метаболических наномеханизмов среди многих групп микроорганизмов, тем самым делая возможным строгий контроль потоков элементов между организмами. В глобальном масштабе функционирование этих наномеханизмов привело к формированию макроскопического аппарата жизнеобеспечения, контролирующего потоки основных газов.
Контролирующие устройства встроены в химические сигналы, которыми микроорганизмы обмениваются внутри сообщества и которые несут информацию о том, кто чем занят и где сколько микроорганизмов находится. Система межклеточной сигнализации, называемая кворумным восприятием, возникла в результате эволюции определенных молекул, производимых и использующихся микроорганизмами для оценки плотности их собственной популяции, а также для передачи своим собратьям сигналов о том, кто они такие и где находятся. Такой способ межклеточной коммуникации пока остается для нас довольно неясным, хотя мы знаем, что некоторые клетки высылают специальные молекулы, которые пребывают в свободном плавании до тех пор, пока не прикрепятся к специальным рецепторным участкам на мембране другого микроорганизма. Примерно того же пытаются добиться парфюмерные компании: чтобы все мужчины ощущали присутствие женщины, и наоборот; так же и эти молекулы, производимые микроорганизмами, сигнализируют другим организмам, кто они и где находятся.
После прикрепления такая молекула принимается за работу по изменению экспрессии содержащихся в клетке генов. Кворумное восприятие позволяет консорции устанавливать пространственную модель микробиологического метаболизма, которая в дальнейшем повышает эффективность рециркуляции питательных веществ. Однако так же она может изменять и поведение консорции.
Здесь вы можете задать резонный вопрос: разве у микробов есть «поведение»? Ответ утвердительный. У них нет мозга, но имеются сенсорные системы, которые зачастую могут быть весьма сложными. Они могут воспринимать сигналы из окружающей среды и друг от друга, передавать сигнал к рецептору и порождать ответную реакцию. Давайте рассмотрим один пример, который и привел к обнаружению кворумного восприятия.
Кворумное восприятие является примером эмерджентного (внезапно возникающего) свойства в социальных взаимоотношениях микроорганизмов. Оно было случайно открыто в 1979 году двумя друзьями и коллегами – Кеном Нильсоном, в то время работавшим в Океанографическом институте Скриппса, и Дж. Вудландом (Вуди) Хастингсом из Гарвардского университета. Этих ученых интересовало, как функционируют люминесцентные бактерии, живущие в светящихся органах некоторых морских рыб. В таких органах бактерии расположены с необычайно большой плотностью – свыше 100 млрд клеток на кубический миллиметр. Когда микроорганизмы из этих органов были изолированы и выращены в чистой культуре с низкой плотностью популяции, люминесценции не было; однако по мере того как клетки росли и плотность популяции увеличивалась, колонии начали светиться. Нильсон и Хастингс знали, что у бактерий имеется особый набор генов, необходимый для вырабатывания света. Эти гены каким-то образом выключались, когда концентрация выращиваемых клеток была низкой, и начинали работать, когда концентрация повышалась. Исследователи обнаружили, что сигнал, дающий генам команду функционировать, представляет собой особое химическое соединение, выделяемое клетками, и когда его концентрация становится достаточно высокой, клетки в буквальном смысле зажигаются.
Впоследствии над кворумным восприятием работали многие микробиологи, и, хотя многое об этом феномене нам пока еще не известно, некоторые из основных принципов уже понятны. Не вызывает сомнения тот факт, что микроорганизмы используют химические сигналы, чтобы «включать» и «выключать» различные функции внутри собственной популяции, а также одновременно в нескольких популяциях других микроорганизмов. Эти химические сигналы являются предвестниками возрастающей сложности, но они далеко не обязательно требуют возникновения новых наномеханизмов. Микробиологическая коммуникация посредством химических сигналов – ключевой механизм для регулирования метаболизма среди различных групп организмов внутри консорции. Однако может случиться и кое-что еще.
Ситуация, когда много различных организмов живут в тесной близости друг к другу, может привести к самым неожиданным последствиям. По-видимому, что-то подобное произошло более двух миллиардов лет тому назад, когда один микроорганизм поглотил другой, однако не только сохранил в себе набор генов поглощенного организма – он оставил весь этот организм. Такой процесс тотального горизонтального переноса генов получил название эндосимбиоза – это симбиотическая связь внутри клетки, или, более точно, симбиотическая связь между двумя клетками, одна из которых размещается внутри другой.
Изначальная концепция может быть прослежена вплоть до статьи, опубликованной в 1883 году Андреасом Шимпером – тем самым немецким ученым, который впервые описал хлоропласты. Он заметил, что хлоропласты в клетках растения делятся таким же образом, каким это делают цианобактерии, и логически вывел, что хлоропласты – на самом деле цианобактерии, живущие внутри клетки. Гипотезу Шимпера подхватил русский ботаник Константин Мережковский, который изучал лишайники, представляющие собой симбиотическую ассоциацию фотосинтезирующих микроорганизмов (зачастую это цианобактерии) и грибов. В 1905 году Мережковский опубликовал статью на русском и немецком языках под названием «О природе и происхождении хроматофор в растительном царстве», в которой высказал предположение о том, что хлоропласты являются симбионтами внутри растительных клеток. Его работа была практически забыта во время Первой мировой войны и последовавшей за ней русской революции – не вследствие самих этих событий, но из-за связанной с ученым скандальной истории. Мережковского обвинили в педофилии, и в 1918 году он бежал сначала во Францию, а затем в Швейцарию. Он продолжал писать о симбиозе, но в 1921 году покончил с собой, и его идеи были преданы забвению.
Идею о том, что внутриклеточное тело может представлять собой ранее самостоятельную бактерию, впоследствии поглощенную клеткой-хозяином, также разрабатывал в 1927 году американский биолог Айвен Уоллин, работавший на соответствующем факультете медицинского колледжа в университете Колорадо. Он заявил, что митохондрии могут быть выращены за пределами своих клеток-хозяев. Позднее было доказано, что образцы митохондрий, предоставленные Уоллином, на самом деле были заражены бактериями, что сильно дискредитировало его работу.
Развитие гипотезы эндосимбиоза получило новый толчок в начале 1960-х годов, когда было обнаружено, что и хлоропласты, и митохондрии содержат в себе свою собственную ДНК, имеющую явственные отличия от ДНК ядра клетки, а также содержат собственный набор рибосом. Таким образом, модель клетки, представляющая ее как куклу-матрешку, получила сильный довод в свою пользу; однако вместе с тем было очевидно, что ни хлоропласты, ни митохондрии не могут воспроизводиться вне своих клеток-хозяев. Более того, анализ последовательностей рибосомальной РНК, проведенный Вёзе и Фоксом как в хлоропластах, так и в митохондриях, показал, что обе эти органеллы произошли от бактерий. Этот анализ однозначно доказал, что гипотезы Шимпера и Уоллина в основе своей были верны: хлоропласты действительно имеют отношение к цианобактериям, а митохондрии имеют родственные связи с другим типом бактерий, представители которого, что любопытно, являются анаэробными фотосинтезирующими организмами.
Представление об эндосимбиозе в конце концов заслужило уважение и широкое признание, когда в 1967 году американский биолог Линн Маргулис написала статью, возрождая гипотезу Мережковского, – не на основе новых данных, но с теоретических позиций. Впоследствии она продолжила выступать с доводами в пользу этой концепции в различных статьях и нескольких книгах. Маргулис была исключительно здравомыслящим ученым и моим хорошим другом. На протяжении большей части своей выдающейся научной карьеры она превозносила концепцию эндосимбиоза как движущей силы эволюции жизни на Земле. Частично она была права.
Феномен эндосимбиоза встречается относительно часто, но очень редко приводит к образованию новой органеллы. Фактически единственные две органеллы, в происхождении которых в результате такого процесса мы можем быть абсолютно уверены, – это митохондрия и хлоропласт; однако события, которые привели к включению этих двух организмов в клетку хозяина, изменили весь ход эволюции. Если бы не эндосимбиоз, нас бы не существовало. В обоих случаях процесс начался в океанах задолго до того, как появились сколь-нибудь значительные свидетельства наземной жизни, и в обоих случаях химическая сигнализация сыграла важнейшую роль.
Загадка истории эволюции эукариотов пока что не до конца разгадана. Представляется, однако, что микроорганизм, послуживший в качестве клетки-хозяина, принадлежал к археям – организмам, похожим на те, что обитают в нашем кишечнике, продуцируя метан. Согласно одному из возможных сценариев, проглоченный организм был близким родственником ныне живущих пурпурных несерных фотосинтезирующих бактерий. Эти бактерии более древние, нежели цианобактерии, и могут использовать световую энергию для фотосинтеза только в том случае, когда в их окружении нет кислорода. В таких условиях они при помощи световой энергии перемещают электроны по замкнутому контуру и выстраивают протонный градиент по разные стороны мембраны. Затем поток этих протонов может проходить через фактор сопряжения, синтезируя АТФ. Это в точности тот же самый наномеханизм, что был описан нами ранее.
При наличии кислорода, однако, электрический контур подавляется, и клетки теряют свою способность синтезировать пигменты, поглощающие свет. Чтобы выжить, они «меняют проводку» в своих внутренних электронных контурах так, чтобы кислород мог стать акцептором водорода, получаемого из органических соединений. Та же самая бактерия, которая днем, в анаэробных условиях, играет роль фотосинтезирующего доктора Джекилла, в аэробных условиях может стать респирирующим мистером Хайдом. На протяжении дня она может использовать солнечную энергию, становясь чистым спонсором органического вещества в микробиотическом мире – но только если поблизости нет кислорода. При наличии кислорода бактерия преображается в потребителя органического вещества и использует энергию его молекул для своего роста. Другими словами, в присутствии кислорода несерные бактерии респирируют, точно так же, как делаем мы и все остальные животные. Животные сохранили в себе внутриклеточного мистера Хайда – митохондрии.
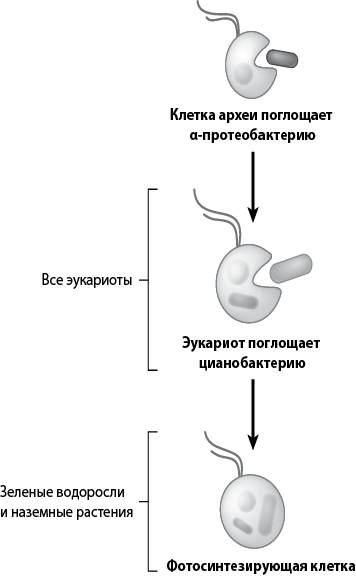
Рис. 27. Схема, отображающая два основных эндосимбиотических события, которые привели к созданию эукариотических клеток. Во время первого события клетка-хозяин (одна из архей) поглотила пурпурную несерную бактерию, которая, возможно, была фотосинтезирующей (значительно позже эта бактерия эволюционирует в митохондрию). Второе событие состояло в том, что клетка, содержащая протомитохондрию, поглотила также цианобактерию (из этих цианобактерий впоследствии разовьются хлоропласты). Два этих первичных симбиотических события являются основой эволюции микроскопических организмов, таких как зеленые водоросли (см. рис. 9), доминировавших в океанах задолго до начала эволюции животных и растений
Каким же образом поглощенная анаэробная фотосинтезирующая бактерия в конечном счете превратилась в поглощающую кислород митохондрию? Наномеханизмы пурпурных фотосинтезирующих бактерий – это в точности те же самые наномеханизмы, при помощи которых мы генерируем энергию в каждой клетке нашего тела, причем это не совпадение, а закономерность. Наши источники энергии – митохондрии – были унаследованы от пурпурных несерных бактерий задолго до того, как появились первые животные. Тем не менее те первоначальные анаэробные пурпурные несерные микроорганизмы, что были поглощены и удержаны клетками архей, почти наверняка не являлись крупными источниками энергии наподобие современных митохондрий. Скорее всего, они представляли собой всего лишь ловушку для питательных веществ, выделяемых клеткой-хозяином. Таким образом, эта эндосимбиотическая анаэробная фотосинтезирующая органелла должна была обладать способностью утилизировать питательные вещества, такие как аммоний или фосфаты, которые в противном случае были бы извергнуты клеткой-хозяином в океан. Я предполагаю, что такая симбиотическая связь была избрана ради сохранения питательных веществ внутри новой, одноклеточной консорции.
Это исключительное событие – поглощение и удержание пурпурной несерной фотосинтезирующей бактерии внутри клетки археи – в конечном счете привело к возникновению первых эукариотических клеток. Лишь гораздо, гораздо позднее одиночные, самостоятельные эукариотические клетки сами образуют организованные консорции, которые впоследствии станут животными и растениями. Однако прежде, чем это могло произойти, механизмы этих протомитохондрий должны были быть запущены в обратную сторону. Весь электронный контур пурпурных несерных бактерий был настроен на производство органических соединений. Современные митохондрии больше этим не занимаются – наоборот, они поглощают органические соединения. Для обращения вспять электронного контура был нужен кислород, однако ни пурпурные несерные фотосинтезирующие бактерии, ни клетка-хозяин не умели его вырабатывать. Такое разделение труда требовало другого набора умений. Однако для того, чтобы это соглашение работало и на хозяина, и на новопоглощенную клетку, двум партнерам было необходимо как-то общаться друг с другом.
По приобретении анаэробной пурпурной фотосинтезирующей бактерии клетка-хозяин должна была как можно быстрее получить контроль над внутриклеточным организмом. Представьте себе, что было бы, если бы внутриклеточный организм рос хотя бы чуточку быстрее, нежели клетка-хозяин. Спустя несколько поколений он перерос бы хозяина, и внешняя клетка бы погибла. Теперь представьте себе обратное: новоприобретенный внутриклеточный организм рос медленнее, чем хозяин. В таком случае хозяин был бы принужден замедлить свой рост и, возможно, не смог бы участвовать в соревновании за получение питательных веществ наравне со своими ничем не обремененными сородичами, не получавшими никакого внутриклеточного организма. Контроль за новоприобретенным внутриклеточным организмом включал в себя перемещение его ключевых генов в клетку-хозяина и потерю внутриклеточным организмом многих других генов. Новая клетка, теперь ставшая эукариотом, превратилась, таким образом, в консолидированную микробиотическую консорцию, в которой клетка-хозяин успешно поработила своего внутриклеточного партнера-эндосимбионта. Со временем внутриклеточный организм потерял столько генов, что больше уже не мог воспроизводиться за пределами клетки-хозяина; тем не менее он сохранил некоторые гены, относящиеся к ключевым наномеханизмам, вырабатывающим энергию, а также отвечающим за способность синтезировать белки. Теперь в пределах одной клетки оказались две фабрики по производству белка.
Контроль за тем, чтобы одна белковая фабрика не вырастала быстрее другой, поначалу, прежде чем были осуществлены перенос и потеря генов, потребовал некоторых усилий. Для этого была необходима система химической сигнализации между двумя клетками – процесс, который до сих пор до конца не выяснен. Химические сигналы посылаются от митохондрии к ядру клетки-хозяина, и одновременно другая сигнальная система действует в обратную сторону. В конечном счете митохондрии превратились в очень сложные механизмы. Они могут «включать» и «выключать» гены в ядре хозяина, усиливать определенные метаболические пути и изменять поведение хозяина. Этой сигнальной системе было дано неудачное название ретроградной сигнализации, но по сути своей она очень похожа на кворумное восприятие – в данном случае у двух клеток, делящих одно пространство, так сказать, сокамерников. Таков был первый шаг к эволюции сотрудничества множества подобных же клеток, функционирующих как единый комплекс. Однако, прежде чем это могло произойти, случилось второе эндосимбиотическое событие.
Второе эндосимбиотическое событие заключалось в том, что анаэробная клетка, уже содержавшая фотосинтезирующую пурпурную бактерию (протомитохондрию), взяла себе еще одного жильца. На этот раз им была кислородпродуцирующая цианобактерия. Вероятно, такая трехэлементная схема была испробована неоднократно, поскольку почти наверняка большинство попыток заканчивалось гибелью анаэробной пурпурной фотосинтезирующей бактерии. За всю историю своей эволюции пурпурные фотосинтезирующие бактерии, скорее всего, ни разу не подвергались воздействию значительного количества кислорода, не говоря уже о целом нескончаемом потоке этого газа, изливавшемся, когда светило Солнце. Давайте проследим логику этого миниатюрного микробиологического зоопарка и посмотрим, как же все это получилось.
Продукт жизнедеятельности цианобактерий кислород должен был использоваться одним из двух других партнеров, который и без того уже был неплохо устроен. В интересы пурпурной фотосинтезирующей бактерии вообще не входило брать нового жильца, но теперь она была поставлена перед угрозой потенциального отравления кислородом, вырабатываемым цианобактерией в той же самой клетке-хозяине. Хозяин, несомненно, был неразборчив, но зачем ему пытаться убить своего первого эндосимбионта, который вполне успешно справлялся со своей задачей по рециркуляции питательных веществ? Чтобы избежать гибели и потенциального исчезновения с лица Земли, пурпурной фотосинтезирующей бактерии необходимо было эволюционировать так, чтобы научиться каким-то образом использовать кислород. Она обнаружила, что кислород, хороший акцептор электронов, может с готовностью принимать электроны от органических соединений, но, для того чтобы этот процесс заработал, необходимо было появление еще одного наномеханизма – такого, который смог бы переносить электроны и протоны к кислороду. Этот наномеханизм, цитохром с-оксидаза, отличался чрезвычайной сложностью; его отдельные части возникли задолго до того, как цианобактерии начали вырабатывать кислород. Из собранных вместе древних компонентов был составлен новый сложный наномеханизм; это происходило путем восстановления и перераспределения компонентов других, более простых наномеханизмов, которые были обнаружены как у бактерий, так и у архей. Цитохром с-оксидаза почти наверняка не был изначально предназначен для транспортировки электронов к кислороду; скорее всего, его эволюционной задачей было удалять кислород из клетки. Современная инкарнация цитохрома с-оксидазы содержит до 13 белковых субкомплексов и использует медь для осуществления своих химических реакций. После появления на эволюционном поле этого наномеханизма мир уже никогда не мог стать прежним.
Благодаря кислороду клетки смогли получить поистине суперзаряд энергии. Использование электрического поля, возникающего по обе стороны от митохондриальной мембраны, позволило синтезировать 36 молекул АТФ из одной молекулы глюкозы. Теперь клетки получили возможность привести в действие маленькие моторчики, вращавшие волосоподобные структуры – жгутики, благодаря чему клетки приобрели невиданную доселе подвижность. Они открыли для себя новые метаболические пути, пользуясь преимуществом обладания кислородом и энергией для создания сложных липидов, таких как холестерин, а также многих других, более сложных молекул. Таким образом, и приобретатели, и приобретенные организмы пришли к тому, чтобы стать «сокамерниками» навечно.
В этой новой клетке, построенной усилиями самих заключенных, имелись потенциальные бонусы для всех участников, однако для того, чтобы механизм мог функционировать, сокамерники должны были научиться работать сообща. При таком новом устройстве в одной клетке заключались целых три набора генетической информации: свой набор генов был у хозяина, свой у протомитохондрии и свой у новоприобретенной цианобактерии – зародыша хлоропласта. Все эти генетические комплекты должны были действовать заодно, так, чтобы ни один из эндосимбионтов не перерос своего хозяина, а хозяин не перерос эндосимбионтов; а для этого требовались некоторые изменения и система сигнализации.
Одной из первых внесенных корректировок была массовая потеря генов новоприобретенной цианобактерией – в точности то же, что мы уже видели в случае с пурпурной фотосинтезирующей бактерией. Цианобактерии было позволено сохранить некоторые из своих генов, чтобы синтезировать важнейшие белки, в особенности те, из которых формируются наномеханизмы фотосинтетических реакционных центров, но многие из генов, позволявших ей расти вне клетки-хозяина, были попросту выкинуты или перемещены в хозяйскую клетку.
Два эндосимбиотических события, заложивших основу для развития эукариотических клеток, являются примерами чисто горизонтального переноса генов, наделившего новую фотосинтезирующую клетку качествами, которые бы она иначе никогда не приобрела. Возникновение зародышевых хлоропластов в клетке, содержащей протомитохондрию, впоследствии дало толчок эволюции многих новых видов – от отдельных лишайников до крупных деревьев. Однако независимо от формы тела все эукариотические фотосинтезирующие организмы используют абсолютно одинаковые древние наномеханизмы для того, чтобы вырабатывать энергию, синтезировать белки и строить новые клетки.
В дальнейшем этим новым организмам предстояло становиться все более сложными и процветающими. И действительно, после Кислородной катастрофы останки эукариотических клеток в осадочной толще встречаются все в больших количествах. С эволюцией эукариотических клеток фаза исследования и развития ключевых наномеханизмов жизни по существу закончилась.
После этого история эволюции уже была связана со строением организмов, то есть в организмах с каким строением наномеханизмы будут размещаться. Эукариотические клетки сами могли формировать консорции и приобретать новые формы. Они могли плавать быстрее и дольше, чем их прокариотические родственники, которых они теперь пожирали, добывая из них питательные вещества. Однако в новых эукариотических клетках также развились новые, более сложные системы коммуникации. Эти системы восприятия представляют собой биллионы химических соединений, обеспечивающих внутри– и межклеточный обмен сигналами – усложненный вариант кворумного восприятия. На протяжении последующих 1,5 млрд лет этим коммуникационным системам предстояло развиться в сложные интегрированные многоклеточные консорции – животных и, позднее, растений.
Давайте теперь посмотрим, как и почему наномеханизмы, развившиеся за 2,5 млрд лет из микроорганизмов, сохранились в макроскопических консорциях эукариотических клеток – животных и растениях, которые были столь же знакомы Дарвину, сколь и любому из нас.
Назад: Глава 6. Защита важнейших генов
Дальше: Глава 8. Крупномеры страны чудес

