Утеха для червей
Будет ли тот или иной ген активирован, зависит от химического состава окружающей среды, а он регулируется работой каскадов других генов, называемых генами-переключателями либо регуляторными генами. Клетки щитовидной железы, например, сильно отличаются от мышечных, как и от всех других, понятное дело, хотя гены у всех одинаковые. Хорошо, скажете вы: клетки мышц и щитовидной железы различаются у более или менее развившихся эмбрионов. Но эмбрион начинается с однойединственной клетки. Клетки щитовидной и поджелудочной желез, клетки печени и костей, клетки кожи и мышц — все это потомки оплодотворенной яйцеклетки, их судьбы складываются в ветвящееся генеалогическое древо. Это древо, берущее начало в зачатии, не имеет ничего общего с эволюционным древом, уходящим в прошлое на миллионы лет, которым мы любовались в других главах. Позвольте показать вам, например, полную родословную 558 клеток только что вышедшей из яйца личинки червя-нематоды Caenorhabditis elegans. (Не знаю, почему этим червям дали видовое название elegans — изящный, но теперь можно сказать, что они его заслуживают. И, хотя я знаю, что не все читатели одобряют отступления от темы, исследования C. elegans — это такое торжество научной мысли, что сейчас меня ничто не остановит.)
В 1960-х блистательный южноафриканский ученый Сидней Бреннер выбрал C. elegans в качестве идеального подопытного животного. Он как раз закончил свою работу с Фрэнсисом Криком и другими коллегами из Кембриджа, посвященную расшифровке генома, и обдумывал, за что бы взяться. С его легкой руки — и благодаря его мощному творческому заделу! — в исследованиях генетики и анатомии нервной системы C. elegans появилось целое сообщество специалистов по C. elegans, которое теперь насчитывает тысячи людей. Можно сказать, что мы знаем о C. elegans почти все. Нам полностью известен его геном. Мы точно знаем, где находится каждая из его 558 клеток (у личинок; у взрослой особи-гермафродита их 959, не считая репродуктивных), и для каждой составлена полная родословная. Нам известно большое количество мутантных генов, приводящих к появлению атипичных червей, и мы точно знаем, где в теле проявляется мутация. Мы знаем и полную историю образования этих отклонений. Это маленькое создание известно нам снаружи и изнутри, вдоль и поперек («О, храброславленный герой!»). В 2002 году Бреннер был, пусть запоздало, удостоен Нобелевской премии, и один из видов, близких к C. elegans, был назван в его честь — Caenorhabditis brenneri. Его постоянная колонка «Дядя Сид» в журнале «Каррент байолоджи» являет образец эрудированности и бестрепетной научной мудрости, остроумной и прекрасной, как вся область исследований C. elegans, им инспирированная. Однако мне хотелось бы, чтобы микробиологи общались с зоологами (как это делал сам Бреннер) и перестали бы относиться к Caenorhabditis как к единственной нематоде или даже как к единственному червю, будто других не существует.
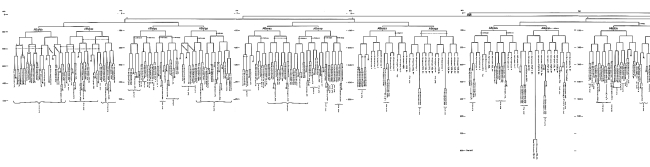

Родословная клеток Caenorhabditis elegans
Разумеется, в диаграмме вы не сможете прочесть, где какие типы клеток находятся (чтобы написать их разборчиво, понадобилось бы семь страниц), но там указано: «глотка», «мышцы кишечника», «мышцы тела», «сфинктер», «кольцевой ганглий», «поясничный ганглий». Все эти клетки вне зависимости от типа приходятся друг другу буквально двоюродными сестрами с учетом их общего происхождения. Я рассматриваю мышечную клетку MSpappppa. Она является сестрой другой мышечной клетки, двоюродной сестрой двух мышечных клеток, двоюродной племянницей еще двух мышечных клеток, троюродной — шести клеткам глотки, четвероюродной сестрой еще семнадцати клеткам глотки, и так далее. Не правда ли поразительно, что мы можем уверенно использовать слова вроде «племянница» по отношению к клеткам, каждая из которых поименована и однозначно определена? Количество «поколений» клеток, отделяющих ткани от первоначальной яйцеклетки, невелико. В конце концов, в теле C. elegans всего 558 клеток, а теоретически можно получить 1024 (два в десятой степени) клеток всего за десять делений. Количество поколений клеток у человека гораздо больше. Тем не менее, теоретически можно составить аналогичную родословную каждой из триллионов клеток (сравните с 558 клетками женской личинки C. elegans), проследив линию предков до одной оплодотворенной яйцеклетки. Впрочем, для млекопитающих невозможно определить и поименовать конкретные клетки, и так, чтобы это соответствовало их расположению. У нас речь идет о некой усредненной популяции клеток, немного различающихся у разных людей.
Надеюсь, что моя ода во славу Caenorhabditis не увела нас слишком далеко от разговора о том, как различные клетки отдаляются друг от друга, меняя форму и другие признаки по мере ветвления древа клеток в ходе эмбрионального развития. В том месте, где двум линиям клеток суждено разойтись в разные стороны, должно произойти нечто, что будет отличать клетку, чья судьба — стать клеткой гортани, от ее сестры-близнеца, которая превратится в кольцевой ганглий. А иначе как они определят, какие гены какой клетке задействовать? Дело в том, что когда ближайший общий предок двух клонов делился пополам, две половины его клетки уже не были одинаковыми. Поэтому две дочерние клетки, хотя и получили одинаковый набор генов, различались по химическому составу. Вследствие этого у них были активированы разные гены, что и определило дальнейшую судьбу их потомков. Этот принцип действует на протяжении всего развития эмбриона, с самого начала. Ключ к клеточной дифференциации у всех животных — асимметричное деление клеток. Сэр Джон Салстон с коллегами проследил путь каждой из клеток тела червя вплоть до одной из шести клеток-основательниц. Мы можем назвать их клетками-родоначальницами: AB, MS, E, D, C и P4. Когда ученые давали клеткам названия, они стремились сделать так, чтобы история каждой была сразу ясна. Название клетки начинается с названия одной из шести клеток-родоначальниц. Дальнейшее название клетки состоит из букв, указывающих направление деления: перед (anterior), зад (posterior), спина (dorsal), брюхо (ventral), лево (left) или право (right). Например, Ca и Cp — две «дочери» клетки-родоначальницы C (передняя и задняя соответственно). Обратите внимание: у каждой клетки может быть не более двух «дочерей» (одна может погибнуть). Вот мышечная клетка Cappppv. У нее говорящее имя. У клетки C была передняя дочерняя клетка, от которой отделилась задняя «дочь», у которой также была задняя «дочь», и у двух следующих также. Последняя породила брюшную дочернюю клетку, на которую я смотрю. Каждая клетка организма описана строчкой, заглавные буквы которой являются именем одной из клеток-основательниц. Еще один пример — клетка ABprpapppap, нервная клетка, расположенная в брюшном нерве, тянущемся по всей длине червя. Наверное, нет необходимости перечислять все ее деления. Замечательно то, что у каждой клетки есть имя, описывающее историю ее развития. Каждое из десяти делений, которые привели к появлению клетки ABprpapppap (понятно, не только ее, но и остальных), было асимметричным, что предопределило активацию разных генов для двух дочерних клеток. Таким же образом происходит дифференциация тканей во всех живых существах. Разумеется, у большинства животных больше 558 клеток, и их эмбриональное развитие, как правило, задано менее жестко. Как по-дружески напомнил мне сэр Джон Салстон (и о чем я уже коротко упоминал), у млекопитающих (особенно у них) клеточные родословные различны для каждой особи, а у Caenorhabditis они практически идентичны, исключая мутантных особей. Тем не менее, принцип остается тем же. У любого животного клетки различных частей тела, хотя и несут одинаковый генетический код, отличаются друг от друга именно из-за асимметричного деления в процессе короткой эмбриональной истории.
Подведем итоги. Не существует никакого плана развития, никакой схемы, никакого чертежа, да и самого проектировщика также нет. Развитие эмбриона, а в итоге и всего организма, определяется набором локальных механизмов, применяемых к отдельным клеткам, взаимодействующих в местном масштабе здесь и сейчас. Внутриклеточные процессы также регулируются локальными правилами, применяемыми к отдельным молекулам, особенно белкам. Повторяю: все эти механизмы и правила локальны — только «здесь и сейчас»! Никто не может, глядя на последовательность ДНК оплодотворенной яйцеклетки, предсказать внешний вид животного. Единственный способ узнать это — позволить яйцеклетке развиваться естественным путем и посмотреть, что выйдет. Ни один компьютер не способен на подобное предсказание, только если он не запрограммирован на имитацию биологических процессов, но в этом случае нам просто не нужен компьютер. Подобный метод создания сложных структур в корне отличен от метода с применением чертежей. Если бы ДНК являла собой своего рода чертеж, вытянутый в строчку, было бы относительно несложно запрограммировать компьютер на чтение схемы и воспроизведение животного. С другой стороны, было бы почти невозможно представить себе, как такое животное могло бы эволюционировать.
Теперь, чтобы эта глава не выглядела отступлением от главной темы книги — эволюции, вернемся к Холдейну и скептически настроенной даме. Если гены определяют развитие эмбриона, а не форму взрослого организма, если ни естественный отбор, ни Бог, в отличие от процессов эмбриогенеза, не изготавливают крылышки, то как действует естественный отбор у животных, когда формирует их тела и поведение? Как естественный отбор действует на эмбрионы, сортируя и вытачивая детали, предоставляя взрослым животным именно то, что им необходимо для выживания: крылья или плавники, щитки или крепкую броню, жало или щупальца?
Естественный отбор — это выборочное выживание успешных генов по отношению к другим, менее успешным. Но естественный отбор не выбирает гены напрямую. Вместо этого он отбирает их носителей — отдельных особей. Этот выбор происходит сам собой. Ненамеренно отбираются те, кто сумел выжить и оставить потомство, передав ему те самые гены. Выживание гена зависит от выживания организма, на образование которого повлиял этот ген: ген, будто всадник, едет вместе с телом, они вместе живут и вместе умирают. Любой ген может существовать одновременно в виде множества копий, путешествующих в большом числе тел в поколении, и передаваться из поколения в поколение в популяции. Соответственно, ген, который в среднем увеличивает шансы на выживание, будет стремиться постепенно к увеличению количества своих копий в популяции. Таким образом, гены, которые мы встречаем в генофонде, скорее всего будут теми генами, которые успешно формируют тела. В этой главе мы увидели, как именно гены формируют тело. Дама, задавшая вопрос Холдейну, полагала, что естественному отбору вряд ли хватило бы миллиарда лет, чтобы создать генетический рецепт изготовления человека. Я же утверждаю, что это вероятно. Хотя, разумеется, ни я, ни кто бы то ни было еще не может подробно рассказать, как это происходило. Это оттого, что все правила имеют локальный характер. Любой акт естественного отбора отбирает мутацию, которая сказалась одновременно на большом количестве клеток многих особей в виде небольшого изменения формы белка. Он, в свою очередь, ускоряет какую-нибудь химическую реакцию во всех клетках, в которых активирован данный ген. Это, например, увеличивает скорость роста зачатка челюсти у эмбриона. Впоследствии это отразится на форме головы (укорачивается морда, черты становятся скорее человеческими, чем обезьяньими). Естественный отбор может оказывать давление каким угодно образом. Например, через эстетическое предпочтение потенциальных половых партнеров. Изменение формы челюсти может оказать небольшое влияние на способность грызть орехи или кусаться. Комбинация давлений естественного отбора, конфликтующих, находящих компромиссы невообразимой сложности, определяет статистический успех данного гена, пытающегося распространиться в генофонде. Сам же ген не подозревает ни о чем. Единственное, что он делает, передаваясь из поколение в поколение, — это изменяет аккуратную впадинку на белковой молекуле. Все остальное — неизбежное следствие небольших каскадных изменений локального масштаба, которые в итоге образуют организм.
Но еще сложнее, чем сложение векторов отбора, действующих в рамках экологических, половых, социальных законов, устроена невообразимо сложная сеть взаимодействий, реализующихся внутри и между развивающимися клетками: влияние генов на белки, генов друг на друга, влияние белков на экспрессию генов, одних белков на другие, мембран, химических градиентов, физических и химических потоков внутри эмбриона, гормонов и других посредников для управления на расстоянии, клеток, нацеленных на клетки с комплементарными маркерами. Никто не может охватить картину целиком, но для того, чтобы принять действенность естественного отбора, это и не требуется. Естественный отбор обеспечивает распространение в генофонде мутаций, которые отвечают за важные изменения в эмбриональном развитии. Картина складывается из сотен тысяч мелких локальных взаимодействий, в принципе понятных (хотя иногда на практике очень сложно перепутанных) любому, кто возьмет на себя труд разобраться. Целое ошеломляет, кажется таинственным, но на самом деле тайны нет. Нам известны и принципы эмбриологии, и механизмы эволюционной истории, ведущей к доминированию в генофонде определенных генов. В процессе эволюции усложнения постепенно накапливались. Каждый шаг уводил ненамного дальше. И каждый шаг совершался за счет небольшого изменения локальных правил. Когда имеется достаточное число элементов (клеток, белковых молекул, мембран), подчиняющихся локальным правилам и влияющих друг на друга, результат может быть впечатляющим. В зависимости от своего влияния на эти элементы, гены могут исчезнуть или, напротив, уцелеть, и тогда в дело вступает естественный отбор. Собеседница Холдейна ошибалась. В этом нет ничего принципиально невозможного. И, как сказал ей Холдейн, на это нужно всего девять месяцев.
Назад: Ферменты
Дальше: Глава 9 Ковчег континентов

