Клетки и скворцы
При клеточном моделировании Остер и его коллеги следовали той же стратегии, которую мы упоминали, говоря о скворцах. Вместо того чтобы задать программу поведения бластуле, они задали программу одной ее клетке. Затем ученые в компьютере «вырастили» много клонов клеток и проверили, что получится, когда эти клетки собраны вместе. Говорить «задали поведение одной клетке» не совсем точно, лучше так: «создали математическую модель одной клетки, построенную на основе известных фактов о клетке», но все же удобнее пользоваться первоначальной простой формулировкой. Особенно важно, что внутреннее пространство клетки связано пересекающимися эластичными нитями-микрофиламентами, способными сокращаться наподобие мускульных волокон. Модель Остера упрощает клетку до двухмерного изображения, удобного для представления на экране компьютера, оставляя всего полдюжины филаментов, соединяющих узловые точки геометрической формы модельной клетки. В модели всем микрофиламентам придаются определенные количественные характеристики, смысл которых хорошо понятен физикам: например, коэффициент вязкого затухания или константа упругости. В действительности неважно, что эти характеристики обозначают; все их физики обожают использовать, когда имеют дело с пружинами или другими упругими телами. В реальной клетке сокращаться способны все Субкортикальный филаменты, но для упрощения модели Остер в своей модели разрешил сокращаться только одной нити из шести. Если реалистичный результат получится даже при отбрасывании ряда известных свойств клетки, то уж при сохранении большей части свойств результат поведения усложненной модельной системы должен только улучшиться. Вместо того чтобы дать микрофиламенту возможность сокращаться как ему угодно, ученые встроили (запрограммировали) свойство, которое присуще любому мышечному волокну: в ответ на растяжение выше определенного предельного значения нить сокращается до меньшей длины, чем в равновесном (не растянутом и не сокращенном) состоянии.
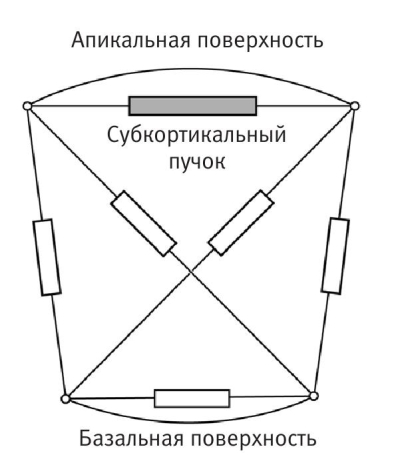
Микрофиламенты внутри модельной клетки Остера
Таким образом, у нас есть сильно упрощенная модель клетки: двухмерный абрис, внутри которого протянуты шесть эластичных нитей, причем одна из нитей способна активно сокращаться в ответ на внешний сигнал растяжения. Это первая стадия процесса моделирования. На второй стадии Остер и его коллеги смоделировали клон из нескольких десятков таких клеток и организовали их в круг, как будто это двухмерная бластула (или тонкий срез бластулы). Затем они взяли и «потянули» одну клетку (запрограммировали мгновенное увеличение натяжения), чтобы спровоцировать сокращение активного микрофиламента. То, что произошло дальше, восхитительно: модельная бластула приступила к гаструляции! Посмотрите на шесть последовательных кадров, запечатлевших это событие. В обе стороны от активированной клетки распространилась волна сокращений, и клеточный шарик начал спонтанно сгибаться и впячиваться.
Дальше — больше. Остер понизил пороговое значение сокращения активного микрофиламента. В результате начавшееся впячивание продолжилось, и в конце концов выщепилась «нервная трубка». Важно понять, что данная модель — это не точная имитация нейруляции. Можно, конечно, отбросить то обстоятельство, что модель работает с двухмерным клеточным кругом со сверхупрощенными внутренними свойствами, но это все равно даже не модельная двухслойная гаструла. Это та же самая бластулоподобная отправная фигура, с которой при других параметрах начинается и гаструляция. Но ведь же моделям и не полагается точно повторять реальные объекты. А наша модель должна показать, насколько просто имитировать важнейшие аспекты клеточного поведения в раннем эмбрионе. То, что даже простой двухмерный кружок из одинаковых клеток спонтанно отвечает неким реалистичным образом на внешний стимул, служит важным доказательством того, что эволюция различных эмбриональных процессов на ранних стадиях вовсе не требует каких-то сложных решений. И нужно помнить, что мы имеем дело с простой моделью, а не с простым явлением. Подобного соотношения следует ожидать от любой хорошей модели.
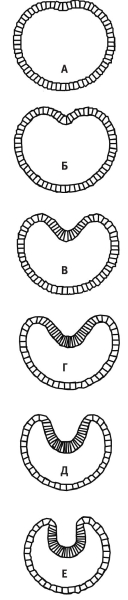
Гаструляция модельной бластулы Остера
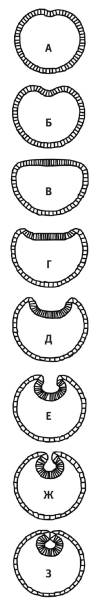
Формирование нервной трубки в модели Остера
Я рассказал о моделях Остера, чтобы продемонстрировать принцип, лежащий в основе взаимодействия клеток друг с другом, — взаимодействия, в результате которого они организуются в оформленное тело. Этот базовый принцип справляется с работой без всяких чертежей, представляющих идею готовой конструкции. Простейшие операции построения раннего эмбриона уже были названы: оригамиподобное складывание, инвагинация и выщепление как в модельной бластуле Остера. Более тонкие операции идут на дальнейших стадиях эмбрионального развития. Так, например, в ходе остроумнейших экспериментов было доказано, что нервные отростки, выходящие из спинного и головного мозга, доходят до места назначения, до своего органа, не следуя заранее установленному плану, а за счет химического привлечения; они подобны псу, находящему по запаху суку. Один из классических ранних экспериментов, превосходно иллюстрирующих этот механизм, провел Роджер Сперри, нобелевский лауреат-эмбриолог. Сперри с коллегами взяли головастика и вырезали у него со спинки и с брюшка одинаковые по размеру квадратики кожи. А потом пришили их обратно, но поменяв местами: кусочек со спинки оказался на брюшке, а с брюшка — на спинке. Когда головастик стал лягушкой, то результат оказался весьма забавным, как это зачастую бывает в эмбриологических экспериментах. У лягушки на темной крапчатой спинке красовался квадратик кожи с брюшка, светлый, как и должно быть на брюшке, а на брюшке выделялся темный квадратик кожи со спинки. Обычно, если пощекотать лягушку кисточкой по спине, она потрет раздраженное место лапой, будто сгоняет муху. Но когда Сперри пощекотал светлый квадратик кожи на спинке своей лягушки, она потерла брюшко! А когда Сперри щекотал темный квадратик на брюшке, то лягушка сгоняла воображаемую муху со спинки.
Согласно Сперри, при нормальном развитии эмбриона происходит следующее: аксон — длинный тонкий трубчатый отросток нервной клетки — выходит из спинного мозга, вынюхивая, словно собачий нос, кожу на животе. Другой аксон вырастает, нацеливаясь на кожу на спине. При ненарушенном развитии все происходит правильно: щекочешь спинку — лягушка чувствует спинку, щекочешь брюшко — брюшко. Но у лягушки Сперри некоторые нервные клетки, идущие на «запах» кожи с живота, оказались на спине, а другие, которые направлялись к коже спины, — на брюшке. Тех, кто верит в теорию «чистой доски», говорящей о том, что индивид рождается без врожденного умственного содержания, начинает жизнь с чистого листа, заполняя его по мере накопления опыта, результат эксперимента с лягушкой должен удивить. По этой теории лягушка должна была бы научиться сопоставлять ощущения с той или иной частью тела. Вместо этого, оказывается, каждая нервная клетка в спинном мозге имеет особую метку, например, нервная клетка для брюшка или нервная клетка для спинки, даже если клетка пока не пустила свой аксон к месту назначения. И клетка находит свой участок кожи, где бы он ни находился. Если бы по спине экспериментальной лягушки ползла муха, то лягушка почувствовала бы, как муха внезапно перескочила на брюшко, поползала там, а потом снова перескочила на спинку.
Эксперименты позволили Сперри сформулировать свою «хемоаффинную» гипотезу, согласно которой нервные волокна протягиваются в нужных направлениях не благодаря наличию общего плана, а в силу того, что каждый аксон ищет свой пункт конечного назначения, следуя индивидуальной химической аффинности, или химическому сродству. Здесь снова срабатывают местные условия и правила. Клетки в изобилии снабжены метками, химическими значками, которые помогают им найти партнеров. Если вернуться к аналогии оригами, то в ней и для принципа мечения найдется место. В бумажном оригами обычно обходятся без клея, но теоретически его можно применить. В эмбриологическом оригами, где телу животного необходимо скреплять клетки, эквивалент клея все же используется. Много клеев, если уж на то пошло, потому что в организме их не один, не два, и метки приходятся как нельзя более кстати. В каждой клетке, на поверхности, где клетки контактируют друг с другом, есть много скрепляющих молекул. Это клеточное склеивание играет важную роль в эмбриональном развитии всех частей тела. Однако клеточные клеи отличаются от привычных нам. Для нас клей — это клей. Им склеивают, и все. Один клей схватывается крепче, другой — быстрее, некоторые типы клея больше подходят для древесины, другие — для металлов или пластика. Но вот, пожалуй, и все, что можно сказать о наших клеях.
Молекулы клеточного клея гораздо более изощренные. Более причудливые, если можно так сказать. Если наш клей склеивает почти любые поверхности, то молекулы клеточного клея приклеятся только к строго определенным склеивающим молекулам. Один из классов таких связывающих молекул у позвоночных называется кадгеринами. Кадгерины включают больше восьмидесяти известных сегодня веществ. За небольшим исключением, из примерно восьмидесяти кадгеринов каждый свяжется только с такой же молекулой. Впрочем, оставим клей, есть аналогия лучше: детская игра, в которой каждому игроку дается метка с названием какого-либо животного, и меток этих по паре на каждого животного. Игра заключается в том, что дети носятся по комнате и издают звуки, характерные для своего животного, а задача игрока — услышать в этой какофонии напарника и извлечь его из толпы. Кадгерины действуют сходным образом. Но и здесь не предполагается наличие плана: действуют микроусловия и локальные правила.
Назад: Кое-что о развитии
Дальше: Ферменты

